










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa zoológica mexicana
versión On-line ISSN 2448-8445versión impresa ISSN 0065-1737
Acta Zool. Mex vol.25 no.2 Xalapa ago. 2009
Artículos originales
Biological and behavioral aspects of two laboratory strains of Ceratitis capitata (Diptera: Tephritidae): The influence of periodic introduction of wild flies in the colony
Aspectos biológicos y comportamentales de dos colonias de Ceratitis capitata (Diptera: Tephritidae): La influencia de la introducción rutinaria de moscas silvestres en la colonia
Iara Sordi JOACHIM–BRAVO 1,*, Alberto MOREIRA DA SILVA NETO1 & Vanessa SIMÕES DIAS1
1 Instituto de Biologia, Universidade Federal da Bahia. R. Barão do Geremoabo s/n Campus Universitário de Ondina, Salvador, Bahia, BRAZIL. 40170–290. *Author for correspondence: e–mail: ibravo@ufba.br; iara_bravo@yahoo.com.br
Recibido: 31/07/2008.
Aceptado: 26/01/2009.
ABSTRACT
The healthy maintenance of insects reared under laboratory conditions requires strategies to retain the natural characteristics of their life–histories traits. Rearing strategies include artificial selection to laboratory conditions, hybridization with compatible strains, and supplying the colony with wild individuals. We compared behavioral as well as life–history aspects of two laboratory strains of Ceratitis capitata Wiedemann (Diptera, Tephritidae) that had either been reared for 15–20 years under laboratory conditions with or without introducing wild specimens (Lab–Pop and Hybrid–pop, respectively). The parameters evaluated were: performance and food ingestion of immatures, adult size and longevity, female oviposition preference and fecundity, egg viability, and mating choice. Analyses of these parameters were conducted under laboratory rearing conditions. The largest differences observed between the two strains were related to behavioral components: food ingestion rate at the larval stage and oviposition behavior. In general, Lab–Pop individuals were less selective and more adapted to the artificial diet than Hybrid–Pop individuals (diet consumption), but there were no significant differences between the two strains in terms of percentage of emergence, egg viability, adult size, fecundity, and mating choice. It is suggested that the use of hybrid strains is more appropriate than the use of strains without the incorporation of wild individuals, especially in studies that involve behavioral parameters, as hybrid strains behave very similarly to wild populations, according our previous studies.
Key words: fruit flies, laboratory rearing, feeding behavior, life history.
RESUMEN
El mantenimiento saludable de insectos criados en condiciones de laboratorio requiere estrategias que conserven características naturales de sus historias de vida. Las estrategias en cría de insectos incluyen selección, hibridación con linajes compatibles y suministro a la colonia con individuos silvestres. Para investigar este último punto, comparamos los aspectos comportamentales así como los aspectos de las historias de vida de dos colonias de laboratorio de Ceratitis capitata Wiedemann (Diptera, Tephritidae) que han sido criadas entre 15 y 20 años en condiciones de laboratorio sin introducir ejemplares silvestres (Lab–Pop), o con introducciones ocasionales (Hybrid–Pop). Los parámetros evaluados fueron: el desempeño y la tasa de ingesta larval, el tamaño y la longevidad de los adultos, la preferencia de oviposición y fecundidad de la hembra, la viabilidad del huevo, y la elección para el apareamiento. Se realizaron análisis de estos parámetros bajo condiciones de cría con una dieta artificial. Las mayores diferencias entre individuos de las dos colonias se registraron en variables comportamentales: tasa de ingesta larval y comportamiento de oviposición. En general, la colonia Lab–Pop fue menos selectiva y más adaptada a la dieta artificial que la colonia Hybrid–Pop (consumo de la dieta), pero no hubo ninguna diferencia significativa entre las dos poblaciones en el porcentaje de emergencia, la viabilidad del huevo, el tamaño de los adultos, la fecundidad, y éxito en el apareamiento. Se sugiere que el uso de colonias híbridas es más apropiado al uso de colonias a las que no se les incorporan ejemplares silvestres, sobre todo en estudios que involucran parámetros comportamentales, dado que las colonias híbridas se comportan como las poblaciones silvestres, de acuerdo con estudios anteriores.
Palabras clave: moscas de las frutas, colonias de laboratorio, comportamiento de alimentación, historias de vida.
INTRODUCTION
Several fruit flies species of the Tephritidae family are reared for basic research as well as control and/or eradication programs, as several fruit flies species are important fruit pests (Christenson & Foote 1960, Leppla 1989).
Several studies have demonstrated modifications in some behavioral, physiological, and demographic characteristics of populations reared under laboratory conditions (Prokopy et al. 1975, Wong & Nakhara 1978, Leppla et al. 1983, Economopoulos 1992). Continued laboratory breeding may result in relaxed selection for some life–history characteristics (Sgro & Partridge 2000, Mack et al. 2001) and may facilitate the accumulation of harmful mutations that can appear in a later phase of adult life (Promislow & Tatar 1998). Appropriate protocols must be followed under laboratory rearing conditions (regardless of the colony size) to avoid these changes when they affect ecological, behavioral or genetic characteristics that are under study or when rearing alters the effectiveness of the insects as agents in pest control strategies (Boller & Chambers 1977).
In order to maintain healthy laboratory rearing regimes, a number of factors should be taken into consideration, including deterioration, excessive homozygosis, or other divergences from the original population (Joslyn 1984). Several basic protocols for the healthy maintenance of these populations have been developed, including re–establishing the lineage through relaxed–rearing, artificial selection of more resistant lineages, hybridization of compatible lineages, and supplying the colony with wild individuals (Leppla 1989, Zucoloto 2000). This latter procedure is commonly used in small–scale rearing programs (for basic or applied research) with the objective of maintaining genetic variability in the populations and reduce the harmful effects of artificial rearing, but it can also be used to restore mating competitiveness in Sterile Insect Programs (Rull & Barreda–Landa 2007). In a previous work (Joachim–Bravo & Zucoloto 1998), flies maintained for long periods of time under laboratory conditions were compared with wild flies in relation to the behavioral parameters of diet selection and oviposition behavior – as well as to some biological parameters, such as duration of larval stage, percentage of emergence and adult size. The results of this research suggested that wild flies were more selective to oviposition choice and were poorly adapted to artificial diet (lower consumption) compared with laboratory ones.
The present study evaluated the effects of introducing wild flies into laboratory colonies of Ceratitis capitata (Diptera: Tephritidae) on several biological and behavioral parameters. Our hypothesis is that the hybrid flies could show some behavioral patterns, typical of wild flies, such as high selectivity for oviposition substrates and for mating choice, which are apparently lost in the process of colonization in the laboratory, and at the same time maintain a good performance in biological parameters.
MATERIAL AND METHODS
Two laboratory strains of C. capitata were compared in these experiments. The first strain (hereafter called Lab–Pop) has been maintained under laboratory conditions for approximately 20 years without any introduction of wild flies; the second strain (hereafter called Hybrid–Pop) has been maintained for approximately 15 years in the laboratory, but with the re–introduction of wild flies (females and males) once every year since the beginning of colonization. For the present study, the Hybrid–Pop was restocked with wild individuals (in numbers equal to approximately 20% of the colony size) once a month for one year before beginning the experiments. Wild flies were always collected from tropical almond fruits, Terminalia cattapa L. (Combretaceae). To ensure lab–wild crossings, wild newly emerged males were exposed exclusively to lab–females in a cage and vice versa. Viable eggs from these crosses were placed in the laboratory colony (Hybrid–pop).
Both stocks were maintained at the Laboratory of Nutritional Ecology of Insects (Department of General Biology/ Institute of Biology/ Federal University of Bahia, BA, Brazil) following rearing procedures described by Cocareli et al. (1986, 1988). The larval diet used to rear both strains and in the experiments described here is described in Zucoloto et al. (1979).
The following aspects of both strains were compared: larval performance, adult size, adult longevity, life–time female fecundity, egg viability, food consumption by immatures, oviposition preference, and mating choice.
All experiments were conducted at 25 ± 1 °C and approximately 70 % r.h., in the dark (experiments involving larvae), or with 10 h of light per day at 400 lux (experiments involving adults).
Data was analyzed using the SPSS statistical program for Windows (version 8.0); Welch's corrected t–test was used to analyze immatures performance, egg viability, estimated diet ingestion by immatures, adult longevity, and female life–long fecundity; a paired t–test was employed for oviposition preference analysis; and sexual preference was analyzed using a Chi–square test. The survival analysis was performed with the Kaplan–Meyer method and comparative survival analysis with log–rank test, using Graph Pad Prism 5 software package. The level of significance was 0.05 for all statistic analyses.
Immature performance. Immature performance was tested following Joachim–Bravo & Zucoloto (1998), as follows: 20 newly hatched larvae of each strain were placed in Petri dishes (90 x 16 mm) lined with slightly moistened filter paper and containing 5.0 mg of artificial diet. The larvae were removed to a new piece of this artificial diet on a daily basis until pupation, and the following parameters were subsequently analyzed: percent of adult emergence, duration of immature stage (from egg hatch to adult emergence), egg production during the pre–oviposition period, which is the time until females start laying eggs (15 females from each strain), and adult size (estimated by measuring the wing nervure, R4+5 to cu–m, of 15 females from each strain, according to Zucoloto (1987). Six replicates were carried out for each test.
Egg viability. Recently emerged adults from both sexes derived from larvae fed on artificial diet were placed into rearing cages supplied with water and a sucrose solution and an oviposition substrate (red agar–agar balls). Oviposited eggs were subsequently collected daily for five days and placed (in groups of 10) in a Petri dish lined with a moistened filter paper piece. A total of 250 eggs from each strain were observed.
Diet consumption. Fifteen 3–day old larvae were placed on 1 g of artificial diet in Petri dishes lined with slightly moistened filter paper. The larvae were carefully removed 3 h later and the remaining artificial diet was weighed. A control dish with no larvae was used to correct for evaporation. For each strain, three dishes with larvae (experimental dishes) and one control dish were used. Procedures details are described in Joachim–Bravo & Zucoloto (1998). The experiment was replicated three times.
Adult longevity. To assess adult longevity, 30 recently–emerged flies of each sex from both strains were placed (in groups of 5 individuals of same sex) in acrylic boxes (11x11x3 cm) and fed with a saturated sucrose solution and water ad libitum (both offered in glass tubes covered with cotton). Mortality was recorded on a daily basis until death of all individuals.
Female fecundity. For female fecundity assessments, ten couples of recently–emerged flies derived from Lab–Pop or Hybrid–Pop larvae fed on artificial diet were placed in acrylic boxes (11x11x3 cm) and maintained as described above. The eggs laid by the females in red agar–agar balls used as oviposition substrates were counted daily from the first day after experiment assembly until all the flies had died. Female fecundity was estimated as the number of eggs laid / the number of alive females in the cages. The experiment was repeated 10 times.
Oviposition preference. A choice–test, in which females were allowed to oviposit on pieces of fruit (mango) or an aliquot of artificial larval diet was used to assess oviposition preference. Ten pairs of recently–emerged flies from both strains were placed in cages (20x20x25 cm) and fed on a yeast–sugar (1:2) based diet with water supplied ad libitum. At the beginning of the peak oviposition period (approximately 10 days after emergence) two pieces of mango (5.0 g) and two pieces of artificial diet (5.0 g) were introduced into the cages to serve as potential oviposition substrates. The mango pulp was partially covered with aluminum foil so that just the skin was exposed. The diet used as substrate was an agar–based diet (agar–agar – 2.0 g, brewer yeast – 6.5 g, sugar – 11.0 g, citric acid – 1.0 g, nipagin – 1.0g and water – 100.0 g), wrapped with parafilm, and presented in a spherical form. The pieces of mango as well as artificial diet were exchanged for fresh ones after 24 h; the experiments were terminated after 48 hours, and were repeated 10 times. Eggs oviposited on each aliquot were counted using a stereo microscope.
Mating behavior. One hundred females from each strain were allowed to choose between Lab–Pop and Hybrid–Pop males. Mating choice tests were carried as follows: one sexually mature female of a given group was put in a plastic cage (16x10x10 cm) simultaneously with one male from each test group (previously marked with different colors). Their mating behavior was observed from 07:00 to 15:00 for two consecutive days. The males were removed from the cage after the first day of observation and replaced at same cage at 07:00 am of the second day. This procedure was performed to assure 100% of females mating. When a mating pair was recorded, all flies were removed from the cage and the experiment was finished.
RESULTS
Immature performance. Both strains showed very similar development in terms of the parameters percentage of emergence, immature stages duration, adult size and egg production in the pre–oviposition period (time until females start laying eggs) (Table 1).
Egg viability. There were no statistical differences between the two strains in terms of mean hatching percentages (Lab–Pop: mean±SD = 81.29 ± 13.90 % and Hybrid–Pop: mean±SD = 83.60 ± 15.20 %; Welch's corrected t test: t = 0,5809, d.f = 47; p = 0.5641).
Diet consumption. The results for estimated food ingestion (mg/larva/day) demonstrated that Lab–Pop larvae ingested larger amounts of artificial diet (Mean±SD = 13.00 ± 2.5 mg) than Hybrid–Pop larvae (Mean±SD = 6.70 ± 2.5 mg) (Welch's corrected t test: t = 5.657, d.f = 16; p < 0.0001).
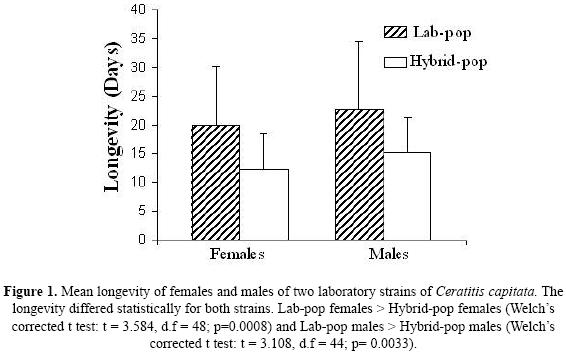
Adult longevity. Results showed that Lab–Pop adults lived longer than Hybrid–Pop adults (Fig.1).
The survival curve for females (Fig. 2A) analyzed by Kaplan–Meyer method also showed differences in mortality patterns between the strains (Log–rank (Mantel–cox) test: X2 = 14.01, d.f = 1; p = 0.0002; hazard ratio = 0.3109). The Hybrid–Pop females suffered reductions of 50% in their populations near the 10th day, while Lab–Pop females demonstrated the same reductions only near the 18th day. After the 25th day, survival was also much more reduced among Hybrid–Pop females (0 % survival by the 30th day) than among Lab–Pop females (0% of survival on the 55th).
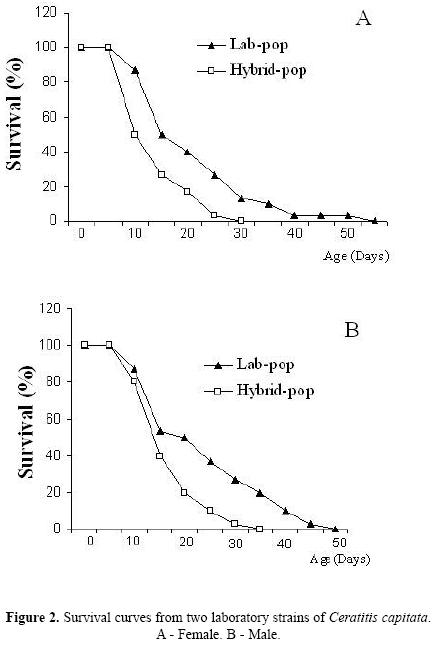
The male survival curve (Fig. 2B), analyzed by Kaplan–Meyer method, showed population reductions of 50 % among Hybrid–Pop males near the 12th day while these same levels were obtained in Lab–Pop males only near the 18th day. Subsequent mortality was also more accentuated in Hybrid–Pop (0 % survival by the 35th day) than in Lab–Pop (0 % survival by the 50th day) (Log–rank (Mantel–cox) test: X2 = 10.69, d.f = 1; p = 0.0011; hazard ratio = 0.3647).
Female fecundity. Lab–Pop females had the shortest pre–oviposition period (Mean±SD = 3.60 ± 0.84 days) and laid the greatest number of eggs during the oviposition period (Mean±SD = 67.30 ± 28.37 eggs/female). In contrast, Hybrid–Pop flies demonstrated the longest pre–oviposition period (Mean ±SD pre–oviposition period = 6.40 ± 2.01 days, and Mean±SD total laid eggs = 27.20 ± 10.18 eggs/female) (Welch's corrected t test: t = 4.060, d.f = 12, p = 0.0016 pre–oviposition period; t = 4.206, d.f = 11, p = 0.0015 total eggs deposited). The life–time fecundity curves (Fig. 3) demonstrated that the total oviposition period was greater for Hybrid–Pop than for Lab–Pop. The maximum time of oviposition for Hybrid–Pop was 22 days and 17 days for Lab–Pop. The two populations also differed in other aspects of their reproductive patterns (Fig.4 and Fig. 5).

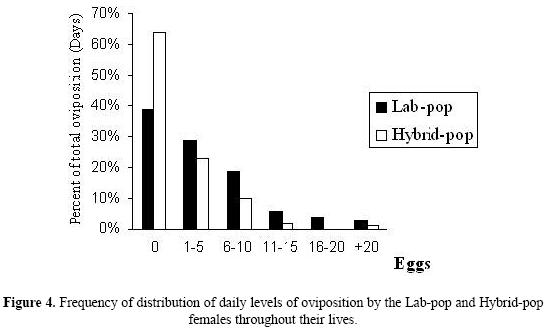
The analyses of the frequency distribution of the number of eggs per day (Fig. 4) showed that Hybrid–Pop females did not oviposit during 64% of their laying period, while Lab–Pop females passed only 39% of their laying period without ovipositing. The period during which the females produced more than 20 eggs per day was also higher for the Lab–Pop than the Hybrid–Pop (3% and 1% respectively).
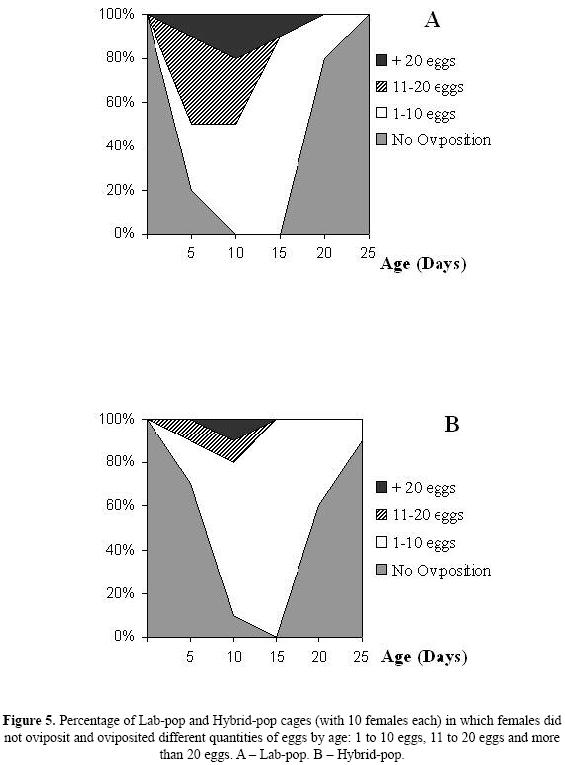
Examination of the percentage of females that laid eggs at different ages (Fig. 5) showed that the fraction of females that did not lay eggs ("no oviposition") in Lab–Pop fell from 100% at day (age) 1, to 0% at age 10 days. This change was more gradual for the Hybrid–Pop, reaching 0% only on the 15th day. After day 15 the percentage of females that did not lay eggs began to rise rapidly in both strains, and at the end of oviposition period (approximately 20 days for Lab–Pop and 25 days for Hybrid–Pop) about 100% of females had ceased oviposition.
Fifty percent of the females from the Lab–Pop strain laid more than 11 eggs per day between the 5th and the 10th day. Most of the Hybrid–Pop females laid from 1 to 10 eggs during almost the entire oviposition period, and only a small percentage of females (approximately 10 to 20%) oviposited 11 or more eggs only in the period from 10 to 15 days (Fig. 5).
Some similarities were found in the reproductive patterns of the two strains. Both strains had oviposition peaks between 6 and 10 days after emergence (Fig. 3) and they produced 1–5 eggs in 20–30% of their posturing periods (Fig. 4).
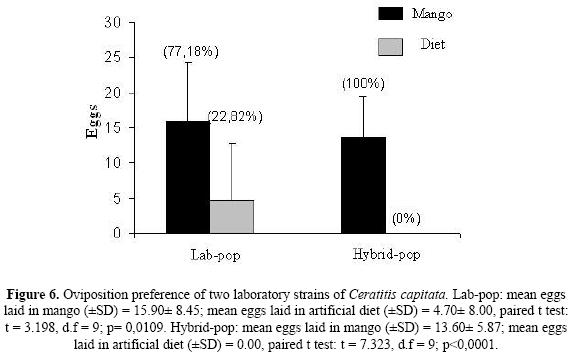
Oviposition preference. Lab–Pop and Hybrid–Pop preferred pieces of mango fruit to the artificial substrate for oviposition (Fig. 6), with Lab–Pop females laying only approximately 20 % of their eggs on the artificial substrate, while Hybrid–Pop females oviposited exclusively on the mango fruit. Lab–pop: mean±SD eggs laid on mango = 15.90 ± 8.45; mean±SD eggs laid on artificial diet = 4.70 ± 8.00, paired t test: t = 3.198, d.f = 9; p = 0.0109. Hybrid–pop: mean±SD eggs laid on mango = 13.60± 5.87; mean±SD eggs laid on artificial diet = 0.00, paired t test: t = 7.323, d.f = 9; p<0.0001.
Mating competitiveness. The evaluation of competitiveness in small cages among males from the two strains showed similar mating competitiveness. Fifty eight percent of the Lab–Pop females (n = 100) chose Lab–Pop males, while 42% chose Hybrid–Pop males (Chi–square test: X2 = 2.56, d.f = 1; p> 0.05). The Hybrid–Pop females (n = 100) showed similar preferences: 53% chose Lab–Pop males while 47% chose Hybrid–Pop males (Chi–square test: X2 = 0.36, d.f = 1; p> 0.05).
DISCUSSION
The results of the present study indicate that the largest differences between the two strains are related to behavioral components: food consumption rate in the larval stage and oviposition behavior. In general, Lab–Pop females were less selective and larvae more adapted to the artificial diet than Hybrid–Pop flies, according the food consumption rate data. Other differences between the two strains were related to longevity (higher for the Lab–Pop) and fertility (pre–oviposition and oviposition periods shorter in the Lab–Pop, although with greater egg production). There were no significant differences observed in terms of emergence percentage, egg viability, adult size, or mating choice between the two strains.
These results indicate that it is not a simple task to evaluate the effects of long–term artificial rearing in the laboratory either in terms of the biological or the behavioral parameters of this species, as a significant effect was observed for some parameters but not for others. This may also reflect that some parameters are more conserved or simply not influenced by long term rearing. Adult size, for instance, seems to be a variable more associated to seeding density instead of genetic deterioration due to inbreeding. In D. melanogaster it has been demonstrated that immature developmental time and fecundity are more affected by laboratory rearing than body size (Sgro & Partridge 2000). It has been shown that populations of some Tephritidae species reared in the laboratory have higher reproductive rates, earlier reproductive periods, and shorter life cycles than wild populations (Carey 1984, Vargas & Carey 1989, Sivinski 1993, Joachim–Bravo & Zucoloto 1998), probably due to selection favoring females with high fecundity.
In the present study, Lab–Pop larvae had a diet consumption rate twice as fast as the Hybrid–Pop larvae. These results could indicate that a Hybrid–Pop strain has a lesser degree of adaptation to that nutrient source, but biological parameters of the immature phase were not visibly affected by this lower rate of nutrient consumption. The artificial diet has high concentrations of proteins compared with fruits, and therefore the amount of diet consumed by larvae could be enough to supply their requirements until emergence.
Both strains had high percentages of emergence, showed similar levels of egg production in the pre–oviposition period, and had high egg viability. Adults of both strains were of similar size upon emergence. All of these results suggest that immatures from both strains responded well to the artificial larval diet.
The two strains showed differences in adult parameters such as longevity and fecundity throughout life. Comparisons of the survival curves of the different populations reared on artificial diet showed that the maximum longevity of both females and males was greater for Lab–Pop than for Hybrid–Pop. These results are in agreement with other studies involving species in which a reduced survival in wild populations reared on an artificial diet has been found (Leppla 1983, Economopoulos 1992, Ekesi et al. 2007).
Lab–Pop females also showed longer oviposition periods, and greater percentages of females produced 11 or more eggs per day over a longer period of time with the artificial diet. The artificial diet contains larger amounts of proteins – one of the most important nutrients for egg production (Dadd 1985). It has been demonstrated that C. capitata reared under laboratory conditions must ingest protein in the immature phase to assure later egg production, and they demonstrated increased egg production when able to ingest these nutrients in the adult phase (Chan et al. 1990, Cangussu & Zucoloto 1995, Kaspi et al. 2002). Our observations support the results from previous studies (Carey 1984, Vargas & Carey 1989, Sivinski 1993, Meats et al. 2004) showing the tendency of laboratory strains to evolve a higher productivity and shorter maturation periods. The results of the present study can be attributed to a better use of the artificial diet by Lab–Pop, or to a reduced ability of Hybrid–Pop flies to adapt to artificial diet or rearing conditions.
Lab–Pop was less selective than Hybrid–Pop in terms of oviposition behavior. Lab–Pop females oviposited approximately 20 % of their eggs in the artificial substrate while Hybrid–Pop females laid their eggs exclusively in mangos. C. capitata females of both strains have adapted to ovipositing directly into holes made in the rearing cages, and have never been directly exposed to any natural or artificial substrate. Nonetheless, it is evident that both strains have conserved their ability to recognize and choose between a fruit and an artificial substrate for egg–laying, with Hybrid–Pop being more selective and rejecting the artificial substrate.
The fact of the laboratory females have been less selective in relation to the oviposition substrate has important implications in the use of such strains in oviposition behavior studies. The oviposition behavior is a central theme in host–plant selection theory, because female choice determines the number of plant species that may be considered hosts for an herbivore insect. Thus, it is suggested that females from laboratory lineages maintained without introduction of wild individuals should be avoided for oviposition experiments because selective ability to recognize sites of breeding may be affected, once its selective capacity to recognize oviposition sites can be affected.
In tests of sexual competitiveness, males from both strains had similar performance in small cages. Studies of mating behavior in C. capitata have shown that various parameters determine male mating success, including body size, body symmetry, protein intake and courtship activity (Blay & Yuval 1997, Orozco & Lopez 1993, Norry et al. 1999, Rodriguero et al. 2002). Considering the good performance of Hybrid–Pop flies during their immature phase, and the large size of emerging adults, it makes sense that Hybrid–Pop males could successfully compete with Lab–Pop males for laboratory–reared females. The data also indicate that females of both strains were similar in terms of selectivity for male choice and did not distinguish males of different strains, probably because both had similar performance. Male competitiveness could have been underestimated because the experiments were conducted in laboratory cages, but previous studies comparing mating competitiveness have shown a similarity of results between experiments performed in laboratory cage and in a field cage (Silva–Neto, 2007).
The use of hybrid strains or the replenishment of laboratory strains with wild males will produce satisfactory results for Sterile Insect Technique (SIT) if the hybrid males can compete with wild males better than non–hybrid strains. In a study of male mating competitiveness with a hybrid strain of A. ludens (produced by wild males x laboratory females), the results indicated that the production of this hybrid strain restored male mating competitiveness in the F1 generation, but without losing desirable laboratory production traits (Rull & Barreda–Landa 2007). The results of our work do not yet allow us to conclude that males from Hybrid–pop would have better competitiveness for mating than the Lab–pop ones, in the field, because they must be tested with wild females and in field cages.
By comparing the over–all results for both strains in the present study with previously published works (Joachim–Bravo & Zucoloto 1998) comparing wild populations with exclusively laboratory reared strains, Hybrid–Pop individuals responded as wild individuals especially in terms of some behavioral parameters such as high selectivity in terms of oviposition behavior and diet consumption rate.
It has been suggested that the experimental use of hybrid strains is more appropriate than exclusively laboratory–reared strains in studies involving behavioral aspects. In the present study, Hybrid–Pop appeared to behave as wild populations, and therefore may represent a realistic model. This result is important to both experimental and applied research because wild populations are difficult to obtain regularly and in large quantities. Additionally, wild populations do not easily adapt to laboratory conditions –showing high mortality rates that can render continued studies unfeasible.
ACKNOWLEDGEMENTS
The authors wish to thank CNPq/PIBIC for grants supporting this research (Process: 520818/89–9).
REFERENCES
Blay, S. & B. Yuval. 1997. Nutritional correlates to reproductive success of male Mediterranean fruit flies. Animal Behavior 54:59–66. [ Links ]
Boller, E.F. & D.L. Chambers. 1977. Quality aspects of mass–reared insects. Pp. 219–235. In: R.L. Ridgway and S.B.Vinson (eds). Biological Control by Augmentation of Nature Enemies. Plenum Press, New York. [ Links ]
Cangussu, J.A. & F.S. Zucoloto. 1995. Self–selection and perception threshold in adult females of Ceratitis capitata (Diptera, Tephritidae). Journal of Insect Physiology 41:223–227. [ Links ]
Carey, J.R. 1984. Host–specific demographic studies of the Mediterranean fruit fly Ceratitis capitata. Ecological Entomology 9:261–270. [ Links ]
Chan, H.T., J.D. Hansen & S.Y.T. Tam. 1990. Larval diets from different protein sources for Mediterranean fruit flies (Diptera: Tephritidae). Journal of Economic Entomology 83:1954–1958. [ Links ]
Christenson, L.D. & R.P. Foote. 1960. Biology of fruit flies. Annual Review of Entomology 5: 171–192. [ Links ]
Cocareli, N.M., J.A. Cangussu & F.S. Zucoloto. 1986. Use of arrowroot as an agar agar substitute in diets for Ceratitis capitata reared in the laboratory. Brazilian Journal of Medical and Biological Research 19:743–745. [ Links ]
Cocareli, N.M., M.I.T. Ferro & F.S. Zucoloto. 1988. Nutritive value of beer yeast for Ceratitis capitata. Revista Brasileira de Zoologia 5:253–259. [ Links ]
Dadd, R.H. 1985. Nutrition: Organisms. Pp. 313–389. In: G.A. Kerkut and L.I. Gilbert (eds) Comprehensive Insect Physiology, Biochemistry and Phamacology. Vol. 4, Pergamon Press, Oxford. [ Links ]
Economopoulos, A.P. 1992. Adaptation of the Mediterranean fruit fly (Diptera: Tephritidae) to artificial rearing. Journal of Economic Entomology 85:753–758. [ Links ]
Ekesi, S., P. W. Nderitu & C.L. Chang. 2007. Adaptation to and small–scale rearing of invasive fruit fly Bactrocera invadens (Diptera: Tephritidae) on artificial diet. Annals of Entomological Society of America 100:562–567. [ Links ]
Joachim–Bravo, I.S. & F.S. Zucoloto. 1998. Performance and feeding behavior of Ceratitis capitata: comparison of a wild population and a laboratory population. Entomologia Experimentalis et Applicata 87:67–72. [ Links ]
Joslyn, D.J. 1984. Maintenance of genetic variability in reared insects. Pp. 20–29. In: E.G. King and N.C. Leppla (eds). Advances and challenges in insect rearing. Agric. Res. Serv., USDA, U.S. Government Printing Office, Washington, D. C. [ Links ]
Kaspi, R., S. Mossinson, T. Drezner, B. Kamensky, & B. Yuval. 2002. Effect of larval diet on developmental rates and reproductive maturation of male and female Mediterranean fruit flies. Physiological Entomology 27:29–38. [ Links ]
Leppla, N.C. 1989. Laboratory colonization of fruit flies. Pp. 91–103. In: A.S. Robinson and G. Hooper (eds). Fruit flies: their biology, natural enemies and control. Vol. 3B, Elsevier, New York. [ Links ]
Leppla, N.C., M.D. Huettel, D.L. Chambers, T.R. Ashley, D.H. Miyashita, T.T.Y. Wong & E.J. Harris. 1983. Strategies for colonization and maintenance of the Mediterranean fruit fly. Entomologia Experimentalis et Applicata 33:89–96. [ Links ]
Mack, P.D., V.K. Lester & D.E.L. Promislow. 2001. Age–specific effects of novel mutations in Droso–phila melanogaster: II. Fecundity and male mating ability. Genetica 110:31–41. [ Links ]
Meats, A., H.M. Holmes & G.L. Kelly. 2004. Laboratory adaptation of Bactrocera tryoni (Diptera: Tephritidae) decreases mating age and increases protein consumption and number of eggs produced per milligram of protein. Bulletin of Entomological Research 94:517–524. [ Links ]
Norry, F.M., G.E. Calcagno, M.T. Vera, F. Manso, J.C. Vilardi. 1999. Sexual selection on morphology independent of male–male competition in the Mediterranean fruit fly (Diptera: Tephritidae). Annals of Entomological Society of America 92:571–577. [ Links ]
Orozco, D. & R.O. Lopez. 1993. Mating competiveness of wild and laboratory mass–reared medflies: effect of male size. Pp. 185–188. In: M. Aluja and P. Liedo (eds). Fruit Flies: biology and management. Springer, New York. [ Links ]
Prokopy, R.J., G.E. Haniotakis & A.P. Economopoulos. 1975. Comparative behavior of laboratory cultured and wild–type Dacus oleae flies in the field. Pp. 101–108. In: Panel Proceedings Series (IAEA). Panel and Research Co–ordination Meeting on the Sterile–Male Technique for Control of Fruit Flies, Vienna (Austria), 12 Nov 1973 / FAO, Vienna (Austria). Joint FAO/IAEA Div. of Atomic Energy in Food and Agriculture. [ Links ]
Promislow, D.E.L. & M. Tatar. 1998. Mutation and senescence: where genetics and demography meet. Genetics 103:299–314. [ Links ]
Rodriguero, M.S., J.C. Vilardi, M.T. Vera, J.P. Cayol & E. Rial. 2002. Morphometric traits and sexual selection in medfly (Diptera: Tephritidae) under field cage conditions. Florida Entomologist 85:143–149. [ Links ]
Rull, J. & A. Barreda–Landa. 2007. Colonization of a hybrid strain to restore male Anastrepha ludens (Diptera: Tephritidae) mating competitiveness for sterile insect technique programs. Journal of Economic Entomology 100:752–758. [ Links ]
Silva–Neto, A.M. 2007. Efeito do tamanho, da alimentação na fase adulta e da idade no sucesso de cópula de machos de Ceratitis capitata (Diptera: Tephritidae). Dissertation Masters – Instituto de Biologia da Universidade Federal da Bahia. 106 pp. [ Links ]
Sgro, C.M. & L. Partridge. 2000. Evolutionary responses of the life history of wild–caught Drosophila melanogaster to two standard methods of laboratory culture. America Naturalist 156:341–353. [ Links ]
Shimoji, Y. & T. Miyatake. 2002. Adaptation to artificial rearing during successive generations in the west indian sweetpotato weevil, Euscepes postfasciatus (Coleoptera: Curculionidae). Annals of Entomological Society of America 95:735–739. [ Links ]
Sivinski, J.M. 1993. Longevity and fecundity in Caribbean fruit fly (Diptera: Tephritidae): effects of mating, strain and body size. Florida Entomologist 76:635–644. [ Links ]
Vargas, R.I. & J.R. Carey. 1989. Comparison of demographic parameters for wild and laboratory–adapted Mediterranean fruit fly (Diptera: Tephritidae). Annals of Entomological Society of America 82:55–59. [ Links ]
Wong, T.T.Y. & L.M. Nakahara. 1978. Sexual development and mating response of laboratory–reared and native Mediterranean fruit flies. Annals of Entomological Society of America 71:592–596. [ Links ]
Zucoloto, F.S. 1987. Feeding habits of Ceratitis capitata: Can larvae recognize a nutritional effective diet? Journal of Insect Physiology 33:349–353. [ Links ]
Zucoloto, F.S. 2000. Alimentação e nutrição de moscas–das–frutas. Pp. 67–80. In: A. Malavasi & R.A. Zucchi (Eds). Moscas–das–Frutas de importância econômica no Brasil: conhecimento básico e aplicado. Holos, Ribeirão Preto. [ Links ]
Zucoloto, F.S., S. Pushel, C.M. Message. 1979. Valor nutritivo de algumas dietas para Anastrepha obliqua. Boletim de Zoologia da Universidade de São Paulo 4:75–80. [ Links ]

