










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa zoológica mexicana
versión On-line ISSN 2448-8445versión impresa ISSN 0065-1737
Acta Zool. Mex vol.24 no.2 Xalapa ago. 2008
Artículos originales
Distribución y solapamiento espacial de las aves acuáticas y ribereñas en un humedal de zonas semiáridas del NE de México
PALACIO-NÚÑEZ J.1, JIMÉNEZ-GARCÍA D.2, OLMOS-OROPEZA G.1 Y ENRÍQUEZ-FERNÁNDEZ J.3
1 Colegio de Postgraduados Campus San Luís Potosí, MÉXICO.
2 Departamento de Ecología, Universidad de Alicante, ESPAÑA.
3 Colaborador externo. e-mail: jpalacio@colpos.mx
Recibido: 18 de febrero de 2007
Aceptado: 3 de marzo de 2008
RESUMEN
La distribución espacial de una comunidad de aves y sus niveles de solapamiento dentro del humedal de la Media Luna fue el principal objetivo de este trabajo. Se detectó un uso amplio y heterogéneo del sistema y sus recursos, principalmente en relación a dos actividades: alimentación y descanso. Para descanso, la mayoría de las especies se mostraron gregarias y compartieron ramas de árboles o vegetación ribereña, mostrando solapamiento entre ellas; mientras que en actividad de alimentación hubo un solapamiento de nicho menos evidente. La competencia trófica fue evidente entre miembros de un mismo gremio, como algunas garzas (Ardeidae) piscívoras que mostraron solapamientos mayores al 75% entre sí, pero la repartición temporal de recursos de las especies nocturnas como Nycticorax nycticorax L, o la diferente dieta de Bubulcus ibis L redujeron la competencia. Algunas especies como Jacana spinosa L realizaron todas sus actividades en un sólo tipo de hábitat, con solapamiento menor al 10% con las demás especies. En general se observó poca agresividad interespecífica. Alteraciones antrópicas en el medio pueden favorecer la abundancia de algunas especies en posible decremento de otras. Los resultados mostraron que la repartición en el uso del hábitat es importante para actividades como descanso, además de la alimentación. Acciones de gestión mal llevadas a cabo tuvieron gran repercusión en el área de estudio, por lo que futuras acciones deben considerar información sobre el uso y la repartición de recursos por parte del grupo de las aves, para diferentes actividades.
Palabras clave: humedales, manantial de la Media Luna, solapamiento de nicho, heterogeneidad espacial, repartición de recursos, diversificación de uso de recursos.
ABSTRACT
Spatial distribution of bird community and their niche overlap on Media Luna wetland was the main objective of this work. A wide and heterogeneous use of the system resources were detected, mainly related to feeding and resting activities. By resting activity, most of the species were gregarious, sharing branches and riparian grassland, showing overlap among them. Whereas in feeding activity there was less evident overlap. Piscivorous egrets and herons (Ardeidae) showed high overlap among them, but temporal resources partitioning of nocturnal species such as Nycticorax nycticorax L, or different diet of Bubulcus ibis L, reduce the competence. Some species such as Jacana spinosa L realised all its activities in an only type of habitat with the lowest overlap with another species. Low interespecific aggressiveness was observed. Anthropic disturbances may favoured some species abundance and possible decrease of another species. Bad management actions had high consequences in the study area. So, future management actions must consider information of use and resource partitioning of bird group.
Key words: wetlands, Media Luna spring, niche overlap, spatial heterogeneity, resource partitioning, resource diversification use.
INTRODUCCIÓN
Ante la transformación y degradación de los humedales, ha sido necesario implementar programas y estrategias de conservación a nivel global. Para estos fines, se propuso la consideración de humedales prioritarios (Ramsar 1971). Considerando que en México los recursos económicos son limitados, se ha propuesto una cantidad mínima de estos, buscando aquellos capaces de albergar la mayor diversidad y abundancia de individuos, enfatizando en las especies relevantes en función de su estado de riesgo de extinción, importancia económica, etc. Para esto se seleccionaron áreas con características tales como gran tamaño, riqueza y relevancia, obteniendo así mayor ahorro y representatividad. Sin embargo, pequeños humedales naturales o artificiales constituyen un importante hábitat, al menos durante cortas temporadas, para gran número de aves acuáticas residentes y migratorias (Pérez-Arteaga et al. 2005).
Los humedales suelen ser ricos en biodiversidad, de manera tal que pueden contener diversas comunidades (Allan & Flecker 1993; Cooperrider & Noss 1994). La distribución de los seres vivos depende de factores de diversa índole que abarcan escalas espaciales y temporales muy distintas, desde globales hasta locales (Nogués-Bravo 2003), o debidas a interacciones (Perry & Dixon 2002). Las comunidades ecológicas son inherentemente complejas (Winemiller & Pianka 1990) por la permanente existencia de un número importante de variables del hábitat que influyen sobre las especies, y la gran cantidad de especies que coexisten e interactúan (Gotelli & Graves 1996; Palmer et al. 2003). En general, las interacciones son amplias y diversas, involucran a gremios multiespecíficos de competidores que, en conjunto, son un mecanismo importante que estructura a las comunidades naturales (Albrecht & Gotelli 2001; Palmer et al. 2003). La interacción entre especies se refiere a la acción negativa activa o pasiva respecto al acceso a los recursos entre una especie u otra, y el término "gremio" se refiere a especies familiares o coordinadas en el uso común de un lote de recursos, tales como la preferencia de hábitat o microhábitat (Mac Nally 1983).
Para el estudio de especies interactuantes es fundamental el enfoque hacia el estudio de solapamiento de nicho. El término nicho es definido como la relación estrecha entre la subdivisión del hábitat y los recursos del ambiente, y se sugiere que al menos existen tres dimensiones principales que determinan el nicho: espacio, recurso trófico y tiempo (Pianka 1969).
Para el estudio de solapamiento de nicho usualmente se usa el término "eje de recursos" o "eje de nicho" para referirse al recurso particular por el cual dos o más especies compiten. Se considera que en todos los casos hay un número grande de estos ejes que las especies se reparten entre sí, siendo difícil saber cual o cuales deben ser medidos (Feisinger et al. 1981). Schoener (1974) argumenta que el microhábitat, la dieta y la actividad temporal son los ejes de nicho más importantes, y que la mayor diferenciación ocurre a lo largo de los dos primeros ejes. Los estudios basados en la dieta confían que la utilización de los diferentes tipos de alimentos es proporcional, pero esta condición reflejaría las interacciones entre el consumidor y su ambiente, más que similitudes entre pares de especies (Lawlor 1980), por lo que numerosos estudios son enfocados sobre gradientes ambientales, tales como microhábitat, y entonces los nichos quedan referidos como preferencias diferenciales del hábitat por parte de las especies (Dolédec et al. 2000).
La aparición de alteraciones en las conductas de uso de los recursos como consecuencia de la presencia de otra especie no indica por si mismo la existencia de competencia, dado que dos especies pueden coexistir cuando hay un adecuado reparto y uso de los recursos (Mac Nally 1983). Muchos autores apoyan la idea de que especies similares (ej. miembros de un gremio) reducen la competencia interespecífica por la subdivisión en distintos niveles de uso de los recursos disponibles (Winemiller & Pianka 1990).
Los pequeños humedales de las zonas semiáridas son especialmente importantes para las aves acuáticas (Pérez-Arteaga et al. 2005) donde tienen una estrecha relación con los recursos potencialmente limitados por el aislamiento. El sistema de la Media Luna, con un área de 8.62 ha de espejo de agua, inmerso en un ambiente semiárido, mantiene poblaciones de los cinco grupos de vertebrados, de los cuales las aves y los peces son los más importantes por su riqueza de especies y la abundancia de las mismas (Palacio-Núñez et al. 2007). El hecho de la coexistencia de al menos 20 especies de aves en este pequeño humedal puede reflejar a priori que hay competencia y una repartición de recursos en diferentes dimensiones (ej: Albrecht & Gotelli 2001). Este sistema se encuentra bajo constantes modificaciones para favorecer la actividad turística, las cuales han tenido repercusiones negativas en la distribución y la abundancia de algunas especies, mientras que otras no han sido afectadas (Palacio-Núñez et al. 2007). El objetivo de este trabajo fue determinar patrones de distribución espacial de una comunidad de aves, y el grado de solapamiento de las especies, dentro de un sistema dulceacuícola ubicado en un ambiente semiárido.
MATERIAL Y MÉTODOS
Área de estudio
El sistema de la Media Luna está localizado en el Valle de Rioverde, San Luis Potosí, en el NE de México (entre las coordenadas UTM: X: 393723 y 395317; Y: 2417647 y 2418070, de la zona 14 N). El clima en el área es seco, semicálido con invierno fresco y temperatura media anual de 21°C; régimen de lluvias de verano con una precipitación anual de 497 mm. La Media Luna se encuentra a una altitud de 1000 msnm (Labarthe et al. 1989).
Este sistema está formado por una laguna-manantial de donde parten una serie de canales, con influencia de humedad por infiltraciones laterales a distancias variables de la fuente constituyendo un humedal, que en época húmeda incluye a dos lagunas estacionales cercanas, a distancias de 800 y 1200 m al sur y sureste respectivamente. La variabilidad ambiental se representó por la porción acuática, el pastizal nativo con sus diferentes grados de antropización y zonas con arbolado ribereño, donde se establecieron 13 sectores cuya estructuración se muestra en la Figura 1. Este sistema tiene uso turístico que ha repercutido en la estructura y calidad de la vegetación acuática y ribereña de manera tal que actualmente algunos sectores están muy antropizados (Cuadro 1).

La plantación del arbolado con fines turísticos se llevó a cabo desde la década de 1950 hasta la de 1980 con las especies Casuarina cunninghamiana Miq y Taxodium sp.; otras acciones de gestión turística comenzaron en la década de 1970. La vegetación nativa característica de las riberas de los canales no alterados presenta una gran dominancia de las gramíneas Panicum bulbosum Kunth y Andropogon glomeratum (Walt) BSP asociadas entre sí y en menor proporción con otras especies. La vegetación acuática está representada básicamente por Nymphaea sp. con una gran cobertura en el sistema (Figura 1) presentando dos tipos morfológicos, uno de los cuales muestra hojas flotantes en algunos sectores (Palacio-Núñez 1997).
Caracterización de la estructura paisajística
La creación de cartografía base de la zona de la Media Luna se basó en la ortofoto digital (F14-C16f) suministradas por el Instituto Nacional de Estadística, Geografía e Informática (INEGI) y una ampliación adicional de fotografía aérea (F14-7) a escala de 1:7500. Esta foto se corrigió con el programa ER Mapper 6.4 a partir de la ortofoto digital georreferenciada. La distribución de la vegetación se realizó mediante la fotografía aérea y numerosos datos de campo. Para la obtención de los mapas de tipos de vegetación terrestre y acuática se emplearon 6 y 4 categorías respectivamente, así mismo se digitalizaron los límites de los sectores y la presencia de canales secundarios; estas capas se superpusieron para obtener la Figura 1.
Toma de datos de las aves en campo
Los datos de campo de la aves se obtuvieron de visitas mensuales en el periodo comprendido entre noviembre de 1996 y mayo de 1997, en el cual se llevaron a cabo un total de 39 repeticiones de un único transecto recorriendo en balsa inflable en cada repetición los 2498 m de la longitud total de los 13 sectores de la Media Luna. Durante cada muestreo se realizó un conteo del total de las aves observadas relacionadas estrechamente al humedal; las especies fueron reconocidas con la ayuda de la guía de campo de Peterson & Chalif (1989) y binoculares. Además de la especie, la abundancia, y el mes de muestreo, se registró la actividad que realizaban los individuos (alimentación, descanso u otra actividad), así como la forma en que se alimentan (por acecho desde la orilla, una percha o desde el aire, captura por buceo, u otro tipo de forrajeo) y la porción particular del hábitat o microhábitat (aguas profundas o someras, vegetación acuática, pastizal o arbolado ribereño). En la zona de la Media Luna habitan numerosas especies de aves acuáticas, sin embargo, se observó que la mayoría de ellas prefieren las lagunas estacionales cercanas a la laguna-manantial, por lo que para el presente trabajo se consideraron 20 especies recurrentes que utilizaron diferentes recursos exclusivamente en la laguna y los canales propios de la Media Luna. Las aves se distribuyeron en ocho órdenes taxonómicos y 10 familias (Cuadro 2).
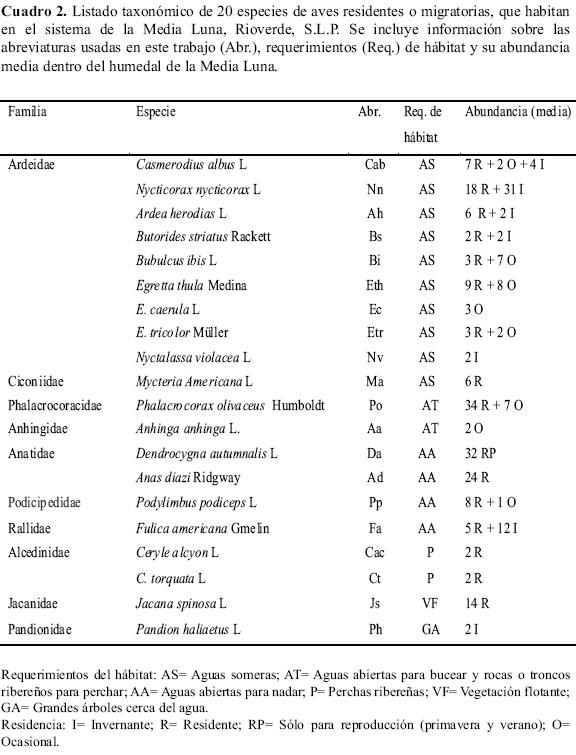
Distribución de las especies
Las especies registradas como R (residentes) en el Cuadro 2 se reprodujeron en la zona en diferentes variantes del hábitat, algunos como M. americana L. anidan en una arboleda a aproximadamente 10 kilómetros de distancia de la Media Luna, siendo el sitio de anidación más lejano registrado (Palacio-Núñez 1997). No se observó competencia interespecífica por los sitios de anidación. Se observó que las aves dividieron su tiempo básicamente entre sesiones de alimentación y descanso. Estas dos actividades fueron consideradas las variables de agrupación para analizar la distribución de las especies entre los sectores de la Media Luna, para esto se realizaron pruebas de análisis factorial de correspondencias (AFC) tomando la abundancia de cada una de las especies en cada circunstancia; usando el programa Statistica 6.0 (StatSoft 2001).
Solapamiento de nicho entre las especies
Una de las mejores herramientas actuales para estimar solapamiento de nicho es mediante el módulo "Niche Overlap" del programa informático EcoSim 7.0. Este modelo se basa en una tabla de medias que muestra la prueba de probabilidad en la cual el solapamiento de nicho medio observado es comparado con el solapamiento de nicho en las comunidades simuladas (Gotelli & Graves 1996).
La amplitud del nicho puede ser retenida o reasignada, cuando es retenido se preserva el grado de especialización de cada especie y cuando es reasignado usualmente conduce a un espectro mucho más amplio de utilización y, por lo tanto, un mayor solapamiento de nicho en la comunidad simulada contra la comunidad real. De la misma manera los estados en cero son retenidos o reasignados: la participación de los ceros en la matriz observada es mantenida o no en cada matriz simulada. Para el presente trabajo se realizaron comparaciones entre los algoritmos RA2, RA3 y RA4 utilizando el índice de Pianka. El algoritmo RA2 flexibiliza la amplitud del nicho observado y retiene la condición de ceros; RA3 retiene la amplitud y reasigna la condición de ceros mientras RA4 retiene ambas condiciones. Cada prueba fue aleatorizada 1000 veces usando el programa informático EcoSim 7.0 (Gotelli & Entsminger 2001). Para este análisis se usó la abundancia total de aves (adultas o inmaduras), en todos los casos las filas contuvieron a las especies y las columnas a las diferentes porciones particulares del hábitat.
RESULTADOS
Distribución espacial de las especies
La gráfica principal del AFC para las aves alimentándose (Figura 2) explica el 69.31% de la varianza. La variabilidad explicada por el eje 1 muestra tres agrupaciones: un primer grupo con las especies Anas diazi Ridgway, Fulica americana Gmelin y Bubulcus ibis L. en condiciones ambientales no alteradas: el segundo grupo con Ardea herodias L., Casmerodius albus L. y Ceryle alcion L. en condiciones similares a las anteriores, pero con arbolado disperso y, el tercer grupo las especies remanentes, más flexibles ante los sectores modificados por presencia de arbolado u otros impactos; Phalacrocorax olivaceus Humboldt y Podylimbus podiceps L. son las especies buceadoras que buscan los espacios de aguas abiertas aún cuando estén en los sectores más antropizados. En este último grupo el eje 2 separa especies como Egretta thula Medina y E. tricolor Müller que se alimentan en aguas muy someras de los sectores 2, 3 y 9.

Para las aves en descanso (Figura 3), la gráfica principal del AFC explica el 51.47% de la varianza; el panorama cambia sensiblemente, formándose dos grupos con respecto al eje 1. El primer grupo suma los dos primeros grupos mostrados en la condición de alimentación, ahora se observa a A. diazi, A. herodias, C. albus, C. alcion y F. americana descansando en los mismos sectores donde se alimentan; pero B. ibis prefiere descansar en los sectores más arbolados. El segundo grupo contiene a las especies y sectores remanentes, P. olivaceus ahora muestra un claro alejamiento del sector 1. Los sectores 1 y 13 se manifiestan como los menos elegidos por las aves para descansar.

Análisis de solapamiento de nicho
Algunas especies como M. americana, Dendrocygna autumnalis L., A. anhinga, Pandion haliaetus L. y N. violacea nunca fueron observadas comiendo dentro del área de estudio y se les dejó fuera de la prueba para solapamiento por sitios de alimentación. Para la actividad de alimentación (Cuadro 3), sólo RA3 mostró significancia en la prueba.

En el Cuadro 4 se muestra el solapamiento de nicho por pares de las 12 especies más representativas que se alimentan en este sistema. Se encontraron valores de solapamiento mayores al 60% para las aves que comparten hábitat alimentario, independientemente de si son competidores tróficos. Sólo las especies C. albus, A. herodias, E. thula y E. tricolor mostraron solapamiento mayor al 75% entre sí. De igual manera, hubo solapamiento de 90% entre las especies buceadoras: P. olivaceus, F. americana y P. podiceps.

Dado que aspectos como la reproducción no reflejaron competencia entre las especies, el solapamiento por el uso del hábitat estuvo dado al menos por los sitios requeridos por las aves para descanso y para alimentación, y entre estas dos variables quedó representado todo el uso espacial del hábitat.
Para el análisis de solapamiento en actividad de descanso se dejaron fuera las especies que aparecieron de manera muy esporádica y con un número muy escaso de individuos, tales como: E. caerula, Nyctalassa violácea L., Anhinga anhinga L., y Ceryle torquata L., realizándose el análisis con las 15 especies remanentes. Los resultados altamente significativos para las medias y para las varianzas muestran que el solapamiento de nicho medio observado fue mayor que el simulado para los tres algoritmos (Cuadro 5). Los tres algoritmos mostraron solapamiento en esta actividad; la mayor diferencia entre las medias estuvo dada por RA3.

En el Cuadro 6 se muestran los resultados del solapamiento de nicho para los pares de especies en actividad de descanso. La mayoría de las aves (67%) comparten sitios de descanso en los árboles y muestran solapamiento elevado (>70 %) entre ellas, con gran contraste respecto a las que no usan los árboles.

DISCUSIÓN
Con base en los resultados del AFC (Figuras 2 y 3), las aves utilizan de manera heterogénea los sectores de la Media Luna y sus recursos, donde se observan concentraciones de especies en algunos puntos, evidenciando preferencias particulares de hábitat, y por tanto repartición espacial, y solapamiento en al menos dos actividades principales: descanso y alimentación. La abundancia de aves varió numéricamente entre las repeticiones del transecto (133.3 ± 27.1), con un mínimo de 69 aves y un máximo de 190.
Para las aves en actividad de alimentación, en términos generales, se observó un solapamiento de nicho menor que en la actividad de descanso para los pares de especies (Cuadro 4). En algunos casos el solapamiento real es menor, ya que especies como P. olivaceus mostraron solapamiento con P. podiceps (90%), F. americana (90%) y A. diazi (61%), sin embargo P. olivaceus es completamente piscívora (Peterson & Chalif 1989); P. podiceps es bastante piscívora, depredando sobre tallas más pequeñas (Smith 2003); las otras dos especies no suelen comer peces por lo que comparten hábitat pero no recursos tróficos.
Todas las especies fueron de actividad diurna, salvo dos garzas (Ardeidae): N. nycticorax y N. violacea que son de hábitos nocturnos (Peterson & Chalif 1989). Según Kronfeld-Schor & Dayan (2003), las especies han desarrollado diferentes ritmos de actividad temporal como un valor adaptativo, sugiriendo la teoría que la repartición temporal puede facilitar la coexistencia entre competidores y entre depredador - presa, por lo que se asume que estas especies redujeron la competencia con las aves diurnas. Otra garza que nunca fue observada alimentándose en el agua fue B. ibis, que se alimenta de insectos terrestres (Peterson & Chalif 1989), por lo que este hábitat alimentario redujo la competencia con las demás garzas que fueron observadas alimentándose preferentemente en el agua.
Las aves de la Media Luna alternan sus periodos de alimentación y descanso en patrones no definidos y en circunstancias diversas. J. spinosa realiza todas sus actividades en el mismo uso espacial (sobre la vegetación flotante), aunque la mayoría de las especies diversifica los usos; aves piscívoras como P. olivaceus. C. albus o Butorides striatus Rackett descansan en árboles o en la vegetación ribereña pero se alimentan en el agua.
Las zonas arboladas de los sectores 4 y 9 (ver Figura 1) constituyen la mayor predilección del conjunto de aves; ocho de las 15 especies se mostraron gregarias y compartieron ramas de árboles o vegetación ribereña para descansar, mostrando solapamiento entre ellas mayor a 70%. Otras especies como J. spinosa, F. americana, P. podiceps o A. diazi no descansan en esta vegetación y mostraron un solapamiento menor al 10% con las especies anteriores, salvo A. diazi con un solapamiento de 31% con C. albus y A. herodias, dado que ambas suelen descansar también en aguas poco profundas. J. spinosa prácticamente no se solapa con ninguna otra especie, sin embargo las aves que suelen descansar en aguas medias: F. americana, A. diazi y P. podiceps presentan un solapamiento mayor al 65% entre ellas (Cuadro 6). Sólo se observó agresividad por sitios de descanso en algunos ejemplares de A. herodias con las demás especies, salvo E. tricolor.
Se observaron algunos aspectos contrastantes: C. albus es gregaria para sus sitios de descanso y de anidamiento, pero para sus sitios de pesca son intolerantes (Jones 2002), conducta que fue observada con algunos incidentes de agresividad por invasión de territorio. A. herodias intolerante en su sitio de percheo fue muy tolerante con C. albus en sus sitios de pesca.
La competencia interespecífica suele ser mayor que la intraespecífica (Vandermeer et al. 2002), sin embargo para las aves de la Media Luna se observó muy poca competencia manifestada mediante agresividad entre especies, pero a nivel intraespecífico se observaron aspectos de territorialidad para J. spinosa y P. podiceps en todas sus actividades, incluyendo reproducción.
La estrecha relación de las especies con los recursos pueden ser difíciles de detectar (Vandermeer et al. 2002). Evidencias de esta relación fueron dadas por malas acciones de manejo en la Media Luna; en 1992 se observó una eliminación de la vegetación acuática y ribereña de varios sectores, con lo que desaparecieron de ahí peces, aves y tortugas. De la misma manera, para enero de 2006 se observó una casi total eliminación de peces, por lo que desaparecieron de la zona la mayoría de las aves piscívoras, siendo muy evidente la total ausencia de las dos especies piscívoras más abundantes P. olivaceus y N. nycticorax (ver Cuadro 2).
Los sistemas dulceacuícolas de zonas áridas como la Media Luna son fundamentales para varias especies de aves tales como A. diazi, amenazada y de distribución restringida básicamente a tierras altas del interior de México, dependiente de pequeños cuerpos de agua como este (Pérez-Arteaga et al. 2002; Pérez-Arteaga et al. 2005); son muy importantes como sitios de dispersión para muchas otras especies migratorias o residentes. Los humedales en general son ricos en biodiversidad y especialmente susceptibles (Cooperrider & Noss 1994), por lo que acciones de manejo mal llevadas pueden ser desastrosas para las especies que lo habitan. Algunas de las modificaciones antrópicas han favorecido la presencia de aves para descansar, pero las especies compiten o se reparten los recursos y sus poblaciones suelen autorregularse sin intervención humana, mientras un estado de hábitat modificado puede favorecer a algunas especies en detrimento de otras, creando un desajuste en la comunidad natural local y el sistema de la Media Luna ha estado sujeto a modificaciones en las últimas décadas (Palacio-Núñez 1997). Estos resultados aportan información útil a considerar sobre el uso y la repartición de recursos por parte del grupo de las aves para dos actividades diferentes, aplicable a este y otros humedales similares, dado que cada vez es más necesario incorporar la información documentada a las acciones de manejo.
AGRADECIMIENTOS
Agradecemos la colaboración en los muestreos de campo a Oscar Nieto Núñez, así como a la Agencia Española de Cooperación Internacional (AECI), Departamento de Becas MAE, por la concesión de la beca No. 32342 y al Consejo Nacional de Ciencia y Tecnología (Beca CONACyT No. Ref: 187777).
LITERATURA CITADA
Allan J.D. & A.S. Flecker. 1993. Biodiversity conservation in running waters. BioScience 43(1): 32-44. [ Links ]
Albrecht M. & N.J. Gotelli. 2001. Spatial and temporal niche partitioning in grassland ants. Oecologia 126: 134-141. [ Links ]
Cooperrider A. & R. Noss. 1994. Saving aquatic biodiversity. Wild Earth Spring: 54-64. [ Links ]
Dolédec S., Chessel D. & C. Gimaret-Carpenter. 2000. Niche separation in community analysis: a new method. Ecological Society of America 81 (10): 2914-2927. [ Links ]
Feinsinger P., Spears E.E. & R.W. Poole. 1981. A simple measure of niche breadth. Ecology 62: 69-76. [ Links ]
Gotelli N.J. & G.R. Graves. 1996. Null models in ecology. Smithsonian Institution Press, Washington. USA. [ Links ]
Gotelli N.J. & G.L. Entsminger. 2001. EcoSim: Null models software for ecology. Version 7.0. Acquired Intelligence Inc. & Kesey-Bear. http://homepages.together.net/~gentsmin/ecosim.htm. [ Links ]
Jones J. 2002. "Ardea alba". Animal Diversity Web. Disponible en: <http://animaldiversity.ummz.umich.edu/site/accounts/information/Ardea_alba.html.> Accedido el 03/05/2006. [ Links ]
Kronfeld-Schor N. & T. Dayan. 2003. Partitioning of time as an ecological resource. Annu. Rev. Ecol. Evol. Syst. 34: 153-181. [ Links ]
Labarthe G., Tristán M., Aguillón R., Jiménez L.S. & A. Romero. 1989. Cartografía Geológica 1:50 000 de las hojas El Refugio y Mineral El Realito, Estados de San Luis Potosí y Guanajuato. U.A.S.L.P., Instituto de Geología. Folleto Técnico Nº 12. S. L. P., México. [ Links ]
Lawlor L.R. 1980. Overlap, similarity, and competition coefficients. Ecology 61 (2): 245-251. [ Links ]
Mac Nally R.C. 1983. On assessing the significance of interespecific competition to guild structure. The Ecological Society of America 64 (6): 1646-1652. [ Links ]
Nogués Bravo D. 2003. El estudio de la distribución espacial de la biodiversidad: conceptos y métodos. Cuadernos de Investigación Geográfica 29: 67-82. [ Links ]
Palacio Núñez J. 1997. Caracterización del ecosistema de la laguna Media Luna, Rioverde, S. L. P. y su influencia en la ornitofauna acuática y ribereña. Tesis de Maestría. Colegio de Postgraduados. Montecillo, México. 140 pp. [ Links ]
Palacio-Núñez J., Verdú J.R., Galante E., Jiménez-García D. & G. Olmos-Oropeza. 2007. Birds and fish as bioindicators of tourist disturbance in springs in semi-arid regions in Mexico: a basis for management. Animal Biodiversity and Conservation 30 (1): 29-41. [ Links ]
Palmer T.M., Stanton M.L. & T.P. Young. 2003. Competition and coexistence: exploring mechanisms that restrict and maintain diversity within mutualist guilds. The American Naturalist. 162: S63-S79. [ Links ]
Pérez-Arteaga A., Gaston K.J. & M. Kershaw. 2002. Population trends and priority conservation for Mexican Duck Anas diazi. Bird Conservation International 12: 35-52. [ Links ]
Pérez-Arteaga A., Jackson S.F. & K.J. Gaston. 2005. Priority sites for wildfowl conservation in Mexico. Animal Conservation 8: 41-50. [ Links ]
Perry J.N. & P. Dixon. 2002. A new method for measuring spatial association in ecological count data. Ecoscience 9: 133-141. [ Links ]
Peterson R. T. & E. L. Chalif, 1989. Aves de México. Guía de Campo. Ed. Diana. México. 473 pp. [ Links ]
Pianka E.R. 1969. Sympatry of desert lizards (Ctenotus) in western Australia. Ecology 50: 1012-1030. [ Links ]
Ramsar. 1971. Accedido en 12/09/07.
Schoener T.W. 1974. Resource partitioning in ecological communities. Science 185: 27-39. [ Links ]
Smith A. 2003. "Podilymbus podiceps". Animal Diversity Web. Disponible en: information/Podilymbus_podiceps.html. Accedido el 31/08/06. [ Links ]
StatSoft, Inc. 2001. STATISTICA (Data Analysis Software System) Version 6. [ Links ]
Vandermeer J., Lawrence D., Symstad A. & S. Hobbie. 2002. Effect of biodiversity on ecosystem functioning in managed ecosystems. In: Loreau M., Naeem S. & P. Inchausti (Eds). Biodiversity and Ecosystem Functioning. Oxford University Presss, UK. Pp. 221-233. [ Links ]
Winemiller K.O. & E.R. Pianka. 1990. Organization in natural assemblages of desert lizards and tropical fishes. Ecol. Monogr. 60: 27-56. [ Links ]

