










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa zoológica mexicana
versión On-line ISSN 2448-8445versión impresa ISSN 0065-1737
Acta Zool. Mex vol.22 no.3 Xalapa dic. 2006
Artículos originales
Informaciones ecológicas sobre Coprophanaeus (Coprophanaeus) telamon nevinsoni Arnaud & Gamez (Coleoptera: Scarabaeinae: Phanaeini) en un sector de la selva húmeda submontana en Mérida, Venezuela
Jorge Gámez, Enrique Mora y Raffaele Acconcia
Cátedra Libre de Estudios Ambientales «Profesor Francisco Tamayo» Facultad de Ciencias, Universidad de Los Andes, Mérida, Venezuela jogamez15@hotmail.com.
Recibido: 23 de mayo 2006
Aceptado: 14 de agosto 2006
RESUMEN
La subespecie Coprophanaeus (Coprophanaeus) telamon nevinsoni Arnaud & Gámez fue analizada tanto en selva como en una plantación mixta utilizándose necrotrampas para el estudio de la actividad anual, diaria y uso del espacio y cebos depositados artificialmente para observar cómo utiliza el recurso carroña. Fue recolectada durante todo el año concentrándose su mayor abundancia (86,63%) en los dos periodos de máxima precipitación. Utiliza indistintamente el ambiente selvático o agrícola en virtud de los corredores de vegetación existentes, sobre todo, los que disponen de elementos arbustivos o arbóreos. Presenta en los ambientes contrastantes evaluados actividad diaria bimodal circunscrita exclusivamente en el crepúsculo, con mayor amplitud temporal, dentro de la selva. Puede emplear el recurso carroña de diferentes tamaños, al contactar fragmentos, los individuos son capaces de realizar el empuje de alguno de ellos a cierta distancia del punto de agregación para luego enterrarlos en galerías de aprovisionamiento. En piezas grandes los escarabajos promueven el entierro parcial y trasladan porciones a cavidades y galerías. Ésta subespecie representa el coleóptero que ejerce mayor influencia en la utilización de la carroña en los ambientes estudiados. Es factible su extinción local debido a la acción antrópica fundamentalmente, por la eliminación de los relictos selváticos aún existentes.
Palabras Claves: Coleóptero necrófago, Scarabaeinae, actividad anual, uso del habitat, actividad diaria.
ABSTRACT
The subspecies Coprophanaeus (Coprophanaeus) telamon nevinsoni Arnaud & Gámez was analyzed in forest and in a mixed plantation using necrotraps in order to study the daily and annual activity, and the using of the space and the bait that was placed artificially in order to observe how it uses carrion resource. It was collected during the year. Its biggest abundance was registered in the two of maximum precipitation (83, 63%). It uses either the forest or agriculture environment, it doesn't matter which takes advantage of the vegetation corridors, mostly those which have arboreal and shrub elements. It shows bimodal daily activity exclusively circumscribed at the twilight in the contrasting environments evaluated. This happens with more temporal amplitude in the forest environment. It may use different sizes carrion resource, because they are capable to transfer it from the aggregation point to the supplying galleries where they bury the carrion. It big pieces, the beetles take the carrion from the parcial burial and take them to storage cavities and galleries. These subspecies represent the most influent beetle in terms of carrion disposition in the studied environments. Its extinction is feasible due to the antropic action, fundamentally for elimination of the relicts existent forest.
Key Words: Carrion beetle, Scarabaeinae, annual activity, habitat use, daily activity.
INTRODUCCIÓN
La conspicua variabilidad climática existente en Los Andes Venezolanos, derivada de la amplitud altitudinal, determina la presencia de variados habitats que han sido agrupados en siete unidades ecológicas, correspondiendo a cada una, un tipo de vegetación característico (Ataroff y Sarmiento 2004). Dentro de estas unidades se presenta la Selva Húmeda Submontana (Selva Húmeda Subandina en La Marca 1998), la cual concentra el mayor número de especies de coleópteros de la subtribu Phanaeini (Gámez 2004). Estos insectos, en conjunto con los demás integrantes del gremio de los Scarabaeinae coadyuvan en el reciclaje de nutrientes contenidos en el excremento o carroña de vertebrados, son fundamentales en la dispersión secundaria de semillas dentro de los ambientes selváticos, promueven la aireación e hidratación del suelo al realizar en el mismo galerías para aprovisionarse de alimento, nidificar o refugiarse, son utilizados en el monitoreo de la biodiversidad o como indicadores de salud ambiental (Halffter & Mathews 1966, Halffter & Favila 1993, Feer 1999, Andresen 2001, Davis et al. 2001, Andresen 2002, Vulinec 2002, Gámez 2004). Las investigaciones sobre Scarabaeinae, han permitido reflejar la clara segregación en torno a las tres dimensiones fundamentales del nicho, esto es, tiempo espacio y alimento (Halffter 1991). A nivel del nicho trófico se destacan los generalistas, copronecrófagos y necrocoprófagos y los especialistas coprófagos y necrófagos. En los Phanaeini, el género Coprophanaeus incluye especies con acusada especialización por la carroña (Edmonds 1972) estando representados en la Selva Húmeda Submontana por Coprophanaeus abas (MacLeay) y Coprophanaeus telamon telamon (Harold), hacia el piedemonte andino llanero y Coprophanaeus telamon nevinsoni Arnaud & Gamez hacia el piedemonte andino lacustre. Sólo es posible revisar información generalizada en relación con éstos coleópteros paracópridos y habiéndose localizado poblaciones bien establecidas de Coprophaneus telamon nevinsoni en un sector de Selva Húmeda Submontana en el Municipio Zea del Estado Mérida, se propuso la indagación de los siguientes aspectos:
1. Determinar el patrón de actividad anual y diaria de la subespecie.
2. Evaluar la preferencia de la subespecie por dos habitats contrastantes: Selva y plantación mixta.
3. Proporcionar información con relación a la forma como utilizan los escarabajos el recurso carroña.
MATERIALES Y MÉTODOS
Área de estudio: Se ubicó en el Municipio Zea del Estado Mérida, específicamente en la aldea Palmarito, sector Las Moras entre las coordenadas U.T.M. 195520 m E y 932800 m N (Fig. 1) a una altitud de 650 m s.n.m. La zona presenta una temperatura media anual de 27°C y una precipitación anual de 2500 mm con dos períodos de lluvia, uno entre abril y mayo y otro de agosto a noviembre. Ambos se alternan con dos períodos secos, uno de diciembre a marzo y otro entre junio y julio (Ministerio del Ambiente y de los Recursos Naturales Renovables. Sistema Nacional de Información Hidrológica y Meteorológica). El sector se corresponde con la Selva Húmeda Submontana la cual ha sido replegada en el área de estudio hacia los sectores en donde se registran pendientes superiores al 30% presentado además, afloramientos rocosos muy acentuados. Estas condiciones probablemente han evitado la utilización completa de ésta unidad ecológica en las labores agrícolas, sin embargo, en la actualidad está siendo fuertemente afectada debido a la extracción maderera sin ningún tipo de control.
En este relicto selvático, de aproximadamente 400 Ha, se evidencian cuatro estratos, con dos arbóreos bien definidos. El superior puede alcanzar alturas mayores a los 50 metros, incluso, se han medido árboles talados de casi 60 metros, presentando los mismos, fustes rectos con los característicos aletones o zancos. Estos árboles portan lianas, bromelias, orquídeas, y aráceas. Dominan en este tipo de selvas las Anacardiaceae, Sterculiaceae, Combretaceae, Moraceae, Burseraceae, Eaeocarpaceae y las Lauraceae, las palmas, son también destacables (Veillon 1989, Ataroff y Sarmiento 2004).
En función de la acción antrópica muchas áreas contiguas a la selva han sido utilizadas para el cultivo de café o café-cambural, observándose la permanencia intencional de ciertas especies arbóreas proporcionadoras de sombra para estos rubros, de igual forma, hay áreas que son cultivadas con cacao, frutales (limón, guanábana, parchita, entre otros) o en el asentamiento de pasturas para la explotación ganadera.
En cuanto a la textura del suelo es de tipo franco arcilloso limoso en la zona de cultivo y en la selva, arcilloso limoso. En ésta última se observa una densa capa orgánica de 08 cm de espesor la cual es inexistente o sólo se mantiene una muy delgada, derivada de los residuos generados por algunos cultivos o de la actividad de roza, en las plantaciones.
En relación con la fauna vertebrada presente se pudo reconocer dentro de la selva, al mono nocturno Aotus trivirgatus (Humboldt), además del avistamiento de Dacypus novencinctus (Linneo) y Coendu sp Lacépéde. Por entrevista a los moradores de la región, señalan la presencia de Alouatta seniculus (Linneo), Marmosops fuscatus (Thomas), Sciurus granatensis Humboldt, Tamandua tetradactyla (Linneo), Choloepus didactylus (Linneo), Potos flavus (Schreber), Cerdocyon thous (Linneo), Leopardus wiedii (Schinz), Agouti paca (Linneo), Dasyprocta puntata (Gray) y Conepatus semistriatus (Boddaert). A estos se agregan algunas especies de quirópteros así como roedores de la familia Muridae. Como grandes reptiles se mencionan a Boa constrictor constrictor Linneo, Bothrops venezuelensis Sandner Montilla y Bothrops asper Garmar y dentro del grupo de los anfibios a Bufo marinus (Linneo). Claro esta, en la actualidad todas estas especies son observadas ocasionalmente ya que han sido afectadas, mucha de ellas, por la casería indiscriminada y por conversión del habitat como se ha señalado.
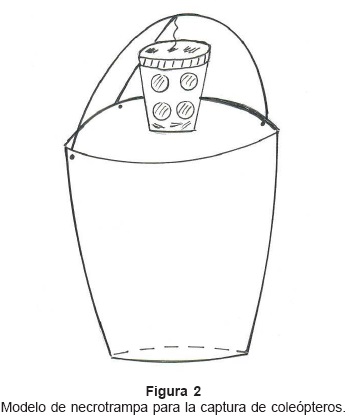
Muestreo: Para el análisis del patrón de actividad anual y preferencia de hábitat se efectuaron muestreos, una vez por mes, entre enero y diciembre de 2004 utilizándose cinco necrotrampas dentro de la selva e igual número en una plantación conformada por cacao-cítricos-pastizal con aproximadamente un kilómetro de separación entre estos habitats contrastantes. En ambos las trampas se instalaron en el suelo, dispuestas en transecta con 20 m de defección entre si. Cada necrotrampa consistió de un envase colector plástico de 11 cm de profundidad y 11,5 cm de diámetro; hacia el borde superior del envase se fijó en dos puntos, siguiéndose el diámetro del recipiente, un alambre de 21 cm de longitud dispuesto en arco. Otro alambre de 10 cm de longitud se fijó en un sólo punto del envase colector conectándose con el anterior hacia el centro y conformando un semiarco. De la conexión de los alambres (centro del envase colector) pende de un trozo de cuerda un recipiente plástico multiperforado de 50 cm3 de capacidad el cual sirve como contenedor del cebo (Fig. 2). Como carnada se utilizó sardina en descomposición y como líquido preservante, contenido dentro del envase colector, agua saturada con cloruro de sodio. Las trampas al instalarse permanecieron activas por 14 horas desde las 17:00 hasta las 08:00 horas del día siguiente. Al final de dicho período se colaba el contenido de las mismas depositándose la entomofauna en envases provisionales de traslado luego, los ejemplares fueron lavados con agua corriente, separadas las diferentes especies y conservadas en alcohol isopropílico al 70% v/v. Coprophanaeus telamón nevinsoni fue procesada para determinar el largo total de machos y hembras así como las proporciones en función de los habitats evaluados.
El análisis de la actividad diaria se efectuó tanto en la selva como en la plantación durante el novilunio utilizándose las necrotrampas de muestreos mensuales con revisiones cada hora desde las 17:00 horas hasta las 08:00 horas del siguiente día. Simultáneamente se registró la temperatura ambiental y la humedad atmosférica para establecer posibles relaciones entre estos parámetros y la actividad de la subespecie.
Para determinar cómo utilizan los individuos la carroña y cómo interactúan entre sí se colocó en el primer período lluvioso del año 2005, durante el novilunio y dentro de la selva, una pieza completa de pescado en descomposición (Prochilodus mariae) de 500 gramos. Con iguales condiciones pero en el segundo período lluvioso se colocaron diez trozos de pescado en descomposición con peso entre 10 y 15 gramos y en otro sector de la selva una pieza de 200 gramos. Las observaciones se efectuaron por 14 horas continuas desde las 17:00 hasta las 08:00 horas del siguiente día. El material recolectado y procesado se encuentra en las colecciones personales de los autores con duplicados en la colección de la Universidad de Los Andes.
RESULTADOS Y DISCUSIÓN
Patrón de actividad anual y uso del espacio
La subespecie fue recolectada durante todo el año exhibiendo dos picos de abundancia que coincidieron con los dos períodos de máxima precipitación (Fig. 3) totalizándose en ellos el 83,63% de las capturas. Fueron recolectados 117 individuos en la plantación y 109 en la selvática, de éstos, 61 machos y 48 hembras en la selva y 59 machos y 58 hembras en el cultivo. La información permite destacar: la presencia permanente de Coprophanaeus telamon nevinsoni durante el año teniendo preponderancia dentro de los invertebrados degradadores en los habitats evaluados; la relación entre la abundancia del escarabajo y el régimen de precipitaciones locales y la plasticidad bioecológica que exhibe la subespecie en función de la conectividad biótica que se presenta entre los habitats, en especial, entre la selva, zona de cultivo y áreas de vegetación secundaria donde se conserven elementos de tipo arbóreo, arbustivo o una mezcla de ambos. En éste escenario se le posibilitaría explorar y explotar los recursos que potencialmente les serian ofrecidos en las zonas antrópicas principalmente, recursos tróficos adicionales derivados de la fauna doméstica o aquella silvestre que utiliza los ambientes modificados. Este último aspecto ha sido señalado a nivel comunitario en donde especies de escarabajos de la subfamilia Scarabaeinae pueden ocupar una gran variedad de habitats alterados por el hombre reduciéndose el aislamiento poblacional, así como los efectos por la pérdida de las zonas selváticas o fragmentación de la misma (Estrada et al. 1998, Escobar y Chacón de Ulloa 2000, Estrada & Coates - Estrada 2002). Para la subespecie en estudio es factible su extinción local si se consolida la desaparición de los relictos selváticos que aún se observan. El suelo, aparentemente no imposibilita la distribución de la subespecie entre los habitats a pesar de presentar diferencias en la textura, materia orgánica y exposición a los elementos climáticos.

Patrón de actividad diaria
En función de los avistamientos y recolecciones efectuadas, Coprophanaeus telamon nevinsoni corresponde a una subespecie que presenta conspicua actividad diaria bimodal circunscrita en el crepúsculo. Dentro de la Selva y en el alba inicia actividad de vuelo a las 06:00 horas registrándose su finalización hacia las 06:45 horas (promedio: 41 minutos). En el ocaso desde las 18:30 hasta las 19:30 horas (promedio: 50 minutos). En la zona de cultivo comienza actividad, de igual forma, a las 06:00 horas hasta las 06:25 horas (promedio: 19 minutos) y en el ocaso por lo general inicia actividad a las 18:40 horas finalizando alrededor de las 19:15 horas (promedio: 30 minutos). De la información se deduce la coincidencia en el inicio de las acciones en los habitats contrastantes por parte de la subespecie presentando mayor amplitud temporal dentro de la selva, la cual podría constituir el habitat de preferencia ecológica primario y desde donde extendería sus actividades a otros lugares, en especial, aquellos que conservan cobertura de tipo arbóreo o arbustivo, como se ha informado. En los dos habitats evaluados el tiempo de actividad es mayor en el ocaso, de igual forma, las mayores recolecciones y avistamientos ocurrieron en éste período de tiempo. Aunque no se registraron variaciones apreciables con relación a la temperatura y humedad poco antes o después del crepúsculo si es factible cambios con relación a la intensidad de la luz tanto diaria como mensual pudiendo afectarse el tiempo de actividad de la subespecie al permitir que los escarabajos prolonguen o acorten la misma. La intensidad de la luz, en particular, se ha mencionado como factor climático que podría modelar o influenciar el patrón de actividad diaria en los coleópteros Scarabaeinae (Martín - Piera et al. 1994) por lo que se haría necesario estudios adicionales con relación a Coprophanaeus telamon nevinsoni para este factor. La condición crepuscular se ha registrado (observaciones puntuales) para otras dos especies en Venezuela recolectadas exclusivamente dentro de la selva en el estado Amazonas: Coprophanaeus (Coprophanaeus) parvulus (Olsoufieff) y Coprophanaeus (Coprophanaeus) dardanus (MacLeay) reflejaron en el alba 39 minutos de actividad y en el ocaso 23 minutos (Jorge Gámez, observaciones personales).
Para cinco especies del género Coprophanaeus, en la Guayana Francesa, Feer y Pincebourde (2005) las registran de condición crepuscular con vuelos entre las 06:00 y 07:00 horas y entre las 18:00 y las 19:00 horas. Para esta delimitación temporal de las especies del género se ha conjeturado que el funcionamiento de las mismas en cortos períodos de tiempo estaría asociado con la menor disponibilidad y predecibilidad de la carroña en comparación con la presencia del estiércol, en consecuencia, cortos períodos de vuelo a alta velocidad podría ser suficiente para determinar si la carroña está localmente disponible reduciéndose concomitantemente el gasto de energía (Gill 1991). Esta conspicua presencia de las especies del género Coprophanaeus en el crepúsculo resulta destacable, si las mismas originariamente habitaban áreas abiertas y posteriormente derivaron como habitantes de selva y de hábitos alimenticios necrófagos ,en parte, debido a la extinción de los herbívoros en el Pleistoceno (Camberfort 1991, Gill 1991, Halffter 1991) y asumiendo bien sea actividad diurna o nocturna, es posible, que se hayan adaptado y se han mantenido así, a ser activas hacia los momentos en los que en la selva se mantendrían condiciones de iluminación que no caracterizan completamente ni a la noche ni al día, es decir, al crepúsculo; siendo éste el momento de actividad y en donde serían competitivamente exitosos. Sin embargo, la adaptación a la selva podría considerarse no estricta, por ejemplo, Coprophanaeus jasius (Oliver) se presenta en habitats abiertos en Venezuela (Martínez y Clavijo 1990, Gámez 2004). Lógicamente, en estas zonas podría existir, de igual forma, carroña potencial pudiendo ser utilizada por especies adaptadas a estos ambientes. Estas características ecológicas exhibidas específicamente por el género Coprophanaeus coadyuvaría en la coexistencia e incremento de la diversidad de coleópteros de la subfamilia Scarabaeinae en un ambiente, como el trópico americano, el cual ha mantenido alta heterogeneidad incluso en el último periodo glacial (Hooghiemstra et al. 2002). Tal heterogeneidad puede haber permitido la gran diversidad biótica que existe en Suramérica la cual ha sido observada para la tribu Phanaeini en ciertas regiones en donde pueden confluir en habitats contrastantes pero cercanos hasta 18 especies en las denominadas intersecciones biogeográficas (Spector 2002).
Utilización del recurso carroña
Dentro de los Scarabaeinae registrados en el área de estudio, Coprophanaeus telamon nevinsoni resultó el necrófago de mayor tamaño ( 24 mm). Las especies necrófilas restantes no superan los 10mm y entre las comparativamente similares en tamaño están Phanaeus hermes (Harold), Oxysternon conspicillatum (Weber) y un cantonino del género Deltochilum. Los dos primeros son de hábitos diurnos y coprófagos, no se constituirían en competidores potenciales, el Deltochilum, aunque puede concurrir en el mismo período de actividad de Coprophanaeus telamon nevinsoni, es copronecrófago y su condición de rodador hace que utilice preponderantemente la parte expuesta del recurso aprovisionándose de bolas del mismo las cuales son alejadas a cierta distancia donde son enterradas, en si, la interacción es lábil.
Coprophanaeus telamon nevinsoni realiza exploración con vuelo en zig-zag y luego en círculos que van cerrándose sobre la carroña a menos de treinta centímetros sobre el suelo. Al contactar trozos de carroña agregados (diez fragmentos de aproximadamente 10 gramos cada uno), los individuos realizan el «reconocimiento» de los mismos y proceden al empuje de alguno de ellos utilizando el clípeo y la parte anterior del tórax, ejerciendo el conjunto de patas, tracción e igualmente empuje. En promedio pueden alejar el fragmento desde el punto de agregación unos 35 centímetros, luego el escarabajo (macho o hembra) realiza una cavidad bajo el recurso y una galería contigua a la misma. Posteriormente procede a halar el trozo utilizando preponderantemente los espolones y partes dentadas de las tibias delanteras tapando finalmente la cavidad. Este proceso lo realizan en aproximadamente 34 minutos. Dentro de las oquedades el escarabajo fragmenta el trozo de carroña y los introduce en la galería.
Al tratarse de piezas de carroña de aproximadamente 200 gramos los individuos, después de un breve reconocimiento, estructuran una cavidad bajo el recurso y una galería diagonal a la misma. Por lo general, entierran parcialmente la carroña y utilizan inicialmente los dientes del clípeo para perforar la piel y las tibias delanteras para rasgar y acumular porciones del recurso que son llevadas dentro de las galerías. Se ha observado en esos trozos individuos realizando el mismo trabajo descrito pero en cavidades y túneles independientes. Al tiempo de trabajado el recurso, cuando se les levanta la pieza, sorprende dos hechos: primero, el tejido muscular y visceral ha sido prácticamente removido quedando expuesto al exterior la piel y debajo la parte esquelética y segundo, la acción de escape violento hacia el fondo de la galería al verse expuesto el escarabajo. El comportamiento «agitadizo» también es observado desde el mismo momento en que el animal aterriza, busca disponer del recurso y enterrarlo de inmediato con ello, evitaría la competencia inter e intraespecífica y el ser depredados sobre todo por quirópteros los cuales fueron frecuentemente observados en el período de actividad de los escarabajos sobrevolando la carroña con movimientos quebradizos.
Ante la pieza de carroña de 500g, el comportamiento del escarabajo es similar al descrito anteriormente. No se observó, en ambos casos, competencias fuera del recurso, ni comportamiento subsocial como ha sido descrito para otros Phanaeini (Halffter & Edmonds 1982). Llegado el período de actividad, los individuos van aterrizando uno a uno, muy rara vez lo hacen simultáneamente más de dos coleópteros. La acción primaria siempre es la de cavar inmediatamente bajo el recurso en trabajo individual como se ha señalado. En una oportunidad, se observó un macho el cual estructuró una galería al lado de una trampa y se colocó de tal forma que asomaba al exterior, las patas traseras y el abdomen. En esta posición produjo frecuentes y fuertes contracciones del abdomen contra los élitros, no se determinó si la conducta del insecto era la de producir algún sonido o feromona para atraer al sexo opuesto.
Las caracterizaciones anteriores se registraron dentro del tiempo de actividad de la subespecie, hacia el final del mismo, tanto en el alba como en el ocaso, los individuos generalmente sobrevuelan el recurso y de repente se alejan rápidamente, sin procurar su utilización.
Lo que ocurra dentro de las galerías, si se dan eventos reproductivos o interacciones antagónicas o beneficiosas son de interés por lo que se propone la realización de estudios complementarios a través de terrarios para dilucidar estas y otras pautas etológicas.
AGRADECIMIENTOS
A W. D. Edmonds (Marfa, Texas) quien realizó comentarios que contribuyeron a mejorar una versión preliminar del manuscrito. De igual forma, el trabajo se benefició de los comentarios editoriales y de dos árbitros anónimos. A Pedro Acuña, por la colaboración desinteresada en los trabajos de campo.
LITERATURA CITADA
Andresen, E. 2001. Effects of dung presence, dung amount, and secondary dispersal by dung beetles on the fate of Micropholis guyanensis (Sapotaceae) seeds in Central Amazonia. J. Trop. Ecol. 17: 61-78. [ Links ]
---------- 2002. Dung beetles in a Central Amazonian rainforest and their ecological role as secondary seed dispersed. Ecol. Entomol. 27: 257-270. [ Links ]
Ataroff, M. & L. Sarmiento. 2004. Las unidades ecológicas de Los Andes de Venezuela. Pp. 11-26. In: La Marca, E. y P J. Soriano. Reptiles de los Andes de Venezuela. Fundación Polar, Conservation International, Codepre-Ula, Biogeos, Mérida, Venezuela. [ Links ]
Cambefort, Y. 1991. Biogeography and evolution. Pp. 51-67. In: I. Hanski & Y Cambefort (eds). Dung Beetle Ecology. Princeton University Press, Princeton, NY. [ Links ]
Davis, A.J., J.D. Holloway, H. Huijbregts, J. Krikken, A. Kirk - Sprigg & S. Sutton. 2001. Dung beetles as indicator of change in the forest of northern Borneo. J. Appl. Ecol. 38: 593-616. [ Links ]
Edmonds, W. D. 1972. Comparative skeletal morphology, systematics and evolution of the Phanaeine dung beetles (Coleoptera: Scarabaeidae). The University of Kansas Science Bulletin 49: 731-874. [ Links ]
Escobar, F. & P. Chacón de Ulloa. 2000. Distribución espacial y temporal en un gradiente de sucesión de la fauna de coleópteros coprófagos (Scarabaeinae, Aphodiinae) en un bosque tropical montano, Nariño - Colombia. Rev. Biol. Trop. 48(4): 961-975. [ Links ]
Estrada, A & R. Coates-Estrada. 2002. Dung beetles in continuos forest, forest fragments and in an agricultural mosaic habitat island at Los Tuxtlas, Mexico. Biodiversity and Conservation. 11: 1903-1918. [ Links ]
Estrada, A; R. Coates-Estrada, A. Anzures Dadda & P. Cammarano. 1998. Dung and carrion beetles in tropical forest fragments and agricultural habitats at Los Tuxtlas, Mexico. J. Trop. Ecol. 14: 577-593. [ Links ]
Feer, F. 1999. Effects of dung beetles (Scarabaeidae) on seeds dispersed by howler monkeys (Alouatta seniculus) in the French Guianan rain forest. J. Trop. Ecol. 15: 1-14. [ Links ]
Feer, F. & S. Pincebourde. 2005. Diel flight activity and ecological segregation within an assemblage of tropical forest dung and carrion beetles. J. Trop. Ecol. 21: 21-30. [ Links ]
Gámez, J. 2004 («2002»). Phanaeini (Coleoptera: Scarabaeinae) de la cordillera de los Andes, depresión de Maracaibo y Llanos de Venezuela. Mem. Soc. Cien. Nat. La Salle. 158: 43-60. [ Links ]
Gill, B. D. 1991. Dung Beetle in Tropical American Forest. Pp. 211-229. In: I. Hanski & Y Cambefort (Eds.). Dung Beetle Ecology. Princeton University Press, New Jersey. [ Links ]
Halffter, G. 1991. Historical and ecological factors determining the geographical distribution of dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae). Folia Entomol. Mex. 82: 195-238. [ Links ]
Halffter, G. & W.D. Edmonds. 1982. The nesting behavior of dung beetles (Scarabaeinae). An ecological and evolutive approach. Instituto de Ecología, Mexico D.F. [ Links ]
Halffter, G & M.E. Fávila. 1993. The Scarabaeinae (Insecta: Coleoptera), an animal group for analyzing, inventorying and monitoring biodiversity in tropical rain forest and modified landscapes. Biol. Int. 27: 15-21. [ Links ]
Halffter, G. & E.G. Matthews. 1966. The natural history of dung beetles of the subfamily Scarabaeinae (Coleoptera, Scarabaeidae). Folia Entomol. Mex. 12-14: 1-312. [ Links ]
Hooghiemstra, H., T. Van der Hammen & A. Cleef. 2002. Paleoecología de la flora boscosa. Pp.43-58. In: Guariguata, M.R. y Kattan, G. (editores compiladores). Ecología y Conservación de Bosques Neotropicales. Editorial Tecnológica de Costa Rica, Cártago, Costa Rica. [ Links ]
La Marca, E. 1998. Biogeografía de los anfibios de la cordillera de Mérida, Andes de Venezuela. Tesis Doctoral, Centro de investigaciones Ecológicas en Los Andes Tropicales. (CIELAT), Universidad de los Andes. [ Links ]
Martín-Piera, F., I. Sanmartín & J.M. Lobo. 1994. Observaciones sobre el ritmo de actividad diaria en Escarabaeidos Telecópridos (Coleóptera, Scarabaeidae). Bull. Soc. Ent. Fr. 99(5): 463-470. [ Links ]
Martínez, A. & J. Clavijo. 1990. Notas sobre Phanaeinas venezolanos, con descripción de una nueva especie de Diabroctis (Coleóptera, Scarabaeidae, Coprini). Bol. Entomol. Venez. 5 (20): 147-157. [ Links ]
Spector, S. 2002. Biogeographic crossroads as priority areas for biodiversity conservation. Conserv. Biol. 16 (6): 1480-1487. [ Links ]
Veillon, J. P. 1989. Los bosques naturales de Venezuela. Parte I. Instituto de Silvicultura, Universidad de los Andes, Mérida, Venezuela. [ Links ]
Vulinec, K. 2002. Dung beetle communities and seed dispersal in primary forest and disturbed land in Amazonia. Biotropica. 34 (2): 297-309. [ Links ]

