










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa zoológica mexicana
versión On-line ISSN 2448-8445versión impresa ISSN 0065-1737
Acta Zool. Mex vol.20 no.3 Xalapa dic. 2004
Estructura anatómica e histológica de la lengua del murciélago frugívoro Centurio senex (Chiroptera: Phyllostomidae)
Cynthia Elizalde-Arellano1, Esther Uría-Galicia2 y Juan Carlos López-Vidal1
1 Laboratorio de Cordados Terrestres, Depto. Zoología, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional. Prol. de Carpio y Plan de Ayala s/n, Col. Casco de Santo Tomas, 11340, México D.F. MÉXICO.
2 Laboratorio de Histología Animal, Depto. Morfología, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional. Prol. de Carpio y Plan de Ayala s/n, Col. Casco de Santo Tomas, 11340, México D.F. MÉXICO. thiadeno@hotmail.com.
Recibido: 25 de agosto 2003
Aceptado: 6 de agosto 2004
RESUMEN
Se analizó la estructura anatómica e histológica de la lengua del murciélago frugívoro Centurio senex, y se comparó con la descrita para otras especies de quirópteros, para conocer si existen estructuras particulares que pudieran estar asociadas a su tipo de alimentación. La lengua de este murciélago es corta, redondeada y posee numerosas papilas filiformes unicúspides, las cortas se ubican en la parte anterior y las largas en la parte media, y cercano a la punta se encuentran pequeñas papilas filiformes con ápice bicúspide; carece de papilas fungiformes y circunvaladas. A nivel histológico, en la parte de la lengua hay un septo fibroso vertical y a ambos lados de él, vasos sanguíneos y nervios en posición simétrica, este arreglo indica que la lengua tiene función intraoral. Entre las fibras musculares estriadas de la parte posterior hay algunos vasos linfáticos, siendo éste el primer registro de la presencia de dichas estructuras en lenguas de murciélagos; también se encontraron nódulos linfoides como en Balantiopteryx plicata y Noctilio leporinus. Las papilas filiformes alargadas y los vasos linfáticos son las únicas estructuras especiales encontradas en Centurio senex.
Palabras Clave: Centurio senex, anatomía, histología, lengua, tejido linfoide.
ABSTRACT
The anatomy and histology of the tongue of Centurio senex, a frugivorous bat, were reviewed to look for any special structure associated to its feeding habit, and to compare it with other bat species with different alimentary habits, documented in other manuscripts. The tongue is short and rounded shape, on its surface there are numerous short single-pointed papillae on the first half, and large papillae at the middle of the surface; near the tip there is a small group of bicuspid papillae; fungiform and circumvallate papillae are absent. Histologically at the part of the tongue, there is a septum with one symmetrical pair of veins, arteries, and nerves at both sides of it, this particular arrangement indicates the intraoral use of this tongue. There are some lymphatic vessels among the muscle fibers, which are the first record of this kind of structures in the tongues of bats; also lymphatic nodules were found similar of those present in Balantiopteryx plicata. The large single-pointed papillae and the lymphatic vessels and nodules are the only specialized structures found in this bat species.
Key Words: Centurio senex, anatomy, histology, tongue, limphoid tissue.
INTRODUCCIÓN
La lengua ha sido objeto de algunos estudios en varias especies de mamíferos (Doran & Baggett 1971a, b y 1972, Doran 1975, Farbman 1970, Geneser 2000, Kubota et al. 1962, Reith 1963), incluido el orden Chiroptera (Arita & Martinez del Río 1990, Elizalde-Arellano et al. 2002 y en prensa, Forman et al. 1979, Gardner 1977, Greenhall et al. 1983, Greenbaum & Phillips 1974, Griffiths 1978 y 1982, Howell & Hodking 1976, Park & Hall 1951, Phillips et al. 1977, Suthers 1970, Tamar 1956, Uieda 1986, Wille 1954).
La lengua de diferentes especies de murciélagos presenta estructuras anatómicas especiales que se relacionan con su tipo de alimentación, como en el caso de algunas especies insectívoras (Elizalde-Arellano et al. 2002), nectarívoras (Griffiths 1978 y 1982) y sanguívoras (Uieda 1986).
Los murciélagos de la especie Centurio senex pertenecen a la familia Phyllostomidae, y su característica más sobresaliente es el rostro achatado y ancho, con numerosos pliegues dérmicos y carente de pelo (Hill & Smith 1984). Su alimentación es frugívora y succionan la pulpa de frutas suaves como higos (Ficus sp.). C. senex habita en zonas tropicales como selvas y matorrales xerófitos, en México se distribuye por ambas vertientes costeras, desde Sinaloa y Tamaulipas hasta el norte de Sudamérica (Hall 1981, Snow et al. 1980), su colecta es poco frecuente por lo que se considera raro y se desconocen muchos aspectos de su biología.
Tomando en cuenta lo anterior, el objetivo del presente estudio es dar a conocer algunas observaciones anatómicas e histológicas de la lengua de Centurio senex con la finalidad de comparar características.
MATERIAL Y MÉTODOS
Se analizaron las lenguas de dos ejemplares de la especie Centurio senex, uno de ellos colectado en el poblado Bajos de Coyula (15° 42' 15" Latitud N, 96° 18' 22" Long W), Municipio de Santa María Huatulco, Oaxaca, y el segundo proveniente de Acaponeta (22° 29' 32" Latitud N, 150° 21' 38") en el municipio del mismo nombre, Nayarit, ambos en México. Las pieles y cráneos fueron procesados por los métodos convencionales (Ramíez-Pulido. et al. 1989) y se encuentran depositados en la colección Mastozoológica del laboratorio de Cordados Terrestres, de la Escuela Nacional de Ciencias Biológicas del IPN; los números de catálogo de éstos murciélagos son 39914 y 41391 respectivamente.
Las lenguas se extrajeron desde su base, tomando como referencia el orificio epiglótico, y se fijaron en formol al 10%. Se midieron con un vernier digital, con precisión de 0.01mm, se esquematizaron con ayuda de una cámara clara Zeiss® y se incluyeron en parafina (Uría & Mora, 1996). Se realizaron cortes longitudinales y transversales de 8 micras de grosor, teñidos con la técnica de Hematoxilina-Eosina (Uría & Mora, 1996).
RESULTADOS
La lengua de Centurio senex tiene forma redondeada y mide en promedio 9.8 mm de largo y 7.0 mm de ancho (Fig. 1). Tres cuartas partes de su superficie dorsal está cubierta por papilas de tipo filiforme, en la mitad anterior la mayoría de ellas son cortas y sus ápices unicúspides (Fig. 1 y 2); entre ellas se observa un pequeño grupo de papilas con ápices bicúspides cercano a la punta. En la parte media de la lengua las papilas filiformes son alargadas y muestran mayor grado de queratinización que las cortas (Fig. 1). En estos murciélagos no se presentan papilas fungiformes ni circunvaladas.
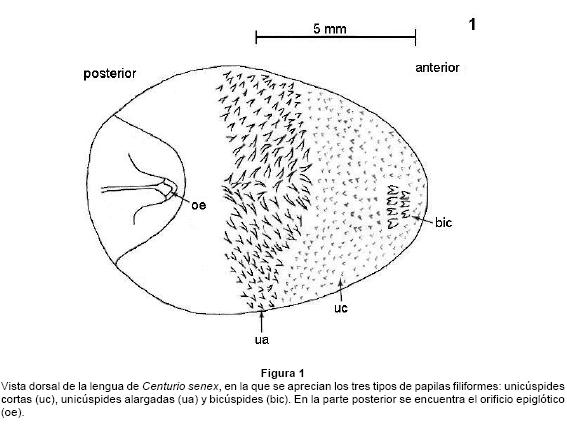
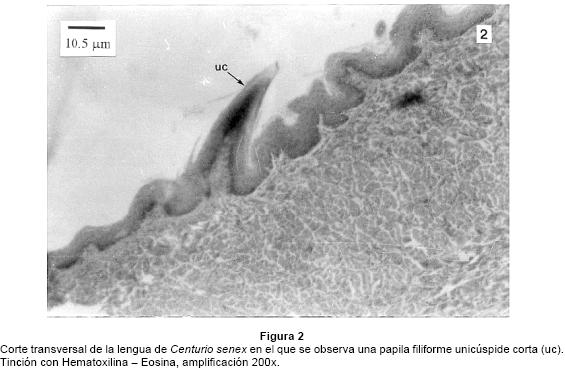
El análisis histológico mostró que el epitelio dorsal es de tipo plano estratificado córneo y el ventral es plano estratificado mucoso. Las fibras musculares estriadas se orientan en direcciones horizontal, vertical y longitudinal. En la parte central de la lengua, hay un septo vertical delgado formado por fibras de tejido conjuntivo que la recorre longitudinalmente, y a ambos lados de él y de manera simétrica hay arterias, venas y nervios.
Entre las fibras musculares cercanas a la zona posterior de la lengua se encuentra una glándula mixta (con acinos serosos y mucosos), así como algunos vasos linfáticos (Figs. 3 y 4). En la parte posterior de la lengua se observaron tres nódulos de tejido linfoide rodeados por una glándula de tipo mucoso, uno en la zona ventral y dos en la parte intermedia (Fig. 5).

DISCUSION
La forma redondeada y el tamaño corto de la lengua de Centurio senex corresponde claramente a la anatomía de su rostro corto y ancho. Las papilas alargadas con mayor queratinización en la parte media de la lengua son hasta ahora una particularidad de Centurio senex, y junto con las papilas bífidas probablemente facilitan la destrucción de los tejidos vegetales al momento de succionar la pulpa de las frutas. Las papilas bífidas también se presentan en otros murciélagos como Balantiopteryx plicata, Macrotus waterhousii (insectívoros), Noctilio leporinus (piscívoro), Phyllostomus hastatus, Brachyphylla cavernarum, Phyllonycteris poeyi (omnivoros), Carollia perspicillata y Artibeus jamaicensis (frugívoros) (Elizalde-Arellano et al. 2002 y en prensa, Griffiths 1982), por lo que éstas no son estructuras asociadas particularmente a la alimentación frugívora.
La ausencia de papilas fungiformes y circunvaladas en Centurio senex, indica que esta especie es incapaz de percibir sabores con la lengua, al igual que Desmodus rotundus (Brightman 1976, Uieda 1986). Sin embargo Centurio senex podría percibir sabores en caso de que existan botones gustativos en el paladar, como ocurre en el caso de los humanos (Geneser 2000), pero para verificar esta característica es necesario realizar estudios adicionales.
El septo fibroso central en la lengua de Centurio senex señala que ésta tiene una función principalmente intraoral (Tipo I) (Doran & Baggett 1971b), como la de los murciélagos insectívoros como Tadarida brasiliensis y Balantipoteryx plicata (Elizalde-Arellano et al. 2002), pescadores en el caso de Noctilio leporinus (Elizalde-Arellano et al. en prensa), y frugívoros como Artibeusjamaicensis (Griffiths 1982). Lo anterior constituye una diferencia fundamental con los murciélagos nectarívoros Leptonicterys nivalis y L. curasoe, cuya lengua carece de septo fibroso central, y las fibras musculares, nervios y vasos sanguíneos se arreglan de manera concéntrica (Griffiths 1982), y su función es extraoral (Tipo II) ya que su elongación es igual o mayor al 100% de su longitud en reposo (Doran & Baggett, 1971b).
De acuerdo a las estructuras del sistema inmune en la lengua, la presencia de los vasos linfáticos observados en Centurio senex además de los nódulos linfoides, indican un grado de desarrollo del sistema inmunológico similar al de Balantiopteryx plicata y Tadarida brasiliensis en los cuales también se registraron nódulos linfoides (Elizalde-Arellano et al. 2002), y al de Noctilio leporinus en el cual se registró un par de amígdalas (Elizalde-Arellano et al. en prensa). No se ha encontrado registro de alguna estructura del sistema inmune en lenguas de otras especies de murciélagos, por lo que en el presente escrito los vasos linfáticos corresponden al primer registro de su presencia para el orden Chiroptera, y los nódulos linfoides son el tercero de estructuras con función inmunológica en la lengua.
La importancia y función precisa que desempeñan algunas de las estructuras mencionadas para la alimentación de Centurio senex, así como su posible significado evolutivo o relación taxonómica con otras especies de murciélagos no puede conocerse con las observaciones con las que se cuenta en el presente escrito, además de estar más allá de los límites del mismo, e intentar descifrar o mencionar alguna de ellas sería caer en el terreno de la especulación. Para establecer con seguridad las diversas funciones o relaciones ecológicas y/o taxonómicas mediante las características linguales de los murciélagos de esta especie es necesario llevar a cabo más estudios con una mayor cantidad de ejemplares, incluyendo también los de otras especies que guarden diferentes grados de parentesco con Centurio senex.
AGRADECIMIENTOS
Agradecemos el apoyo brindado por Hortensia Montellano Rosales del Lab. de Embriología, Olga Ixta Rodriguez y Benjamin Nogueda Torres del Depto. de Inmunología, de la ENCB del IPN, así como también a la Coordinación General de Posgrado e Investigación (CGPI) del Instituto Politécnico Nacional a través del proyecto 980381.
LITERATURA CITADA
Arita T. H. & C. Martinez del Río. 1990. Interacciones flor-murciélago, un enfoque zoocéntrico. Pub. esp. UNAM, 4:1-35. [ Links ]
Brightman V. J. 1976. The vallate-foliate papilla complex and suckling behaviour. Anat. rec. 184(3):363-364. [ Links ]
Doran G. A. & H. Baggett, 1971a. The specialized lingual papillae of Tachiglossus aculeatus I.Gross and light microscopic features. Anat. rec. 172:157-166. [ Links ]
----------. 1971b. A structural and functional classification of mammalian tongues. J. Mamm. 52:427-429. [ Links ]
----------. 1972. The genioglossus muscle: a reassessment of its anatomy in some mammals, including man. Acta anat. 83:403-410. [ Links ]
Doran G. A. 1975. Review of the evolution and phylogeny of the mammalian tongue. Acta Anat. 91:118-129. [ Links ]
Elizalde-Arellano C., E. Uría-Galicia & J. C. López-Vidal. 2002. Morfología comparada de lenguas de Tadarida brasiliensis (I.Geoffroy St. Hilaire, 1824) y Balantiopteryx plicata (Peters, 1867) (Mammalia, Chiroptera). Acta Zool. Mex. (n.s.), 86:103-117. [ Links ]
----------. (En prensa). Morfología lingual del murciélago piscivoro Noctilio leporinus (Chiroptera: Noctilionidae). Acta Zool. Mex. (n.s.).
Farbman, A. I. 1970. The dual pattern of queratinization in filiform papillae on rat tongue. J. Anat. Lond. 106:233-242. [ Links ]
Forman, G. L., C. J. Phillips, & C. S. Rouk. 1979. Alimentary tract. Pp. 205-227, In: R. J. Baker, J.K. Jones Jr. and D. C. Carter, (Eds.) Biology of bats of the New World family Phyllostomatidae. Part III, Spec. Publ. Mus. Texas Tech Univ., 16. [ Links ]
Gardner, A. L. 1977. Feeding habits. Pp, 293-350, In: R. J. Baker, J.K. Jones Jr. and D. C. Carter (Eds.) Biology of bats of the New World family Phyllostomatidae. Part II, Spec. Publ. Mus. Texas Tech Univ., 13. [ Links ]
Geneser, F. 2000. Histología. 3ª edición. Panamericana, México D. F. xvi + 813pp. [ Links ]
Greenbaum, I. F. & C. J. Phillips. 1974. Comparative anatomy and general history of tongues of long nosed bats (Leptonycteris sanborniy L. nivalis) with reference to oral mites. J. Mamm. 55:489-504. [ Links ]
Greenhall, A. M., G. Joerman, & J. Smith. 1983. Desmodus rotundus, Mamm. Species, 202: 1-6. [ Links ]
Griffiths T. A. 1978. Muscular and vascular adaptations for nectar feeding in the Glossophaginae bats, Monophyllus and Glossophaga, J. Mamm. 59: 414.418. [ Links ]
Griffiths T. A. 1982. Systematics of the New World nectar feeding bats (Mammalia: Phyllostomidae) based on morphology of the hioid and lingual regions. Amer. Mus. Novitates, 2742:1-45. [ Links ]
Hall, E. R. 1981. The mammals of North America. 2nd ed. John Wiley and Sons. New York, vol. 1xviii + 1-600+90; vol. 2:xi+601-1181+90. [ Links ]
Hill J.E. & J. D. Smith, 1984. Bats, a natural history. University of Texas Press Austin. 243 pp. [ Links ]
Howell D. & J. Hodking. 1976. Feeding adaptations in the hair and tongue of nectar feeding bats. J. Morph. 148:329-336. [ Links ]
Kubota K., J. Kubota, T. Nakamura, N. Fukuda, S. Asakura, S. Nakagawa & M. Masui. 1962. Comparative anatomical and neurohistological observations on the tongue of the pangolin (Manis pentadactyla, Linneus). Anat. Rec.144:43-45. [ Links ]
Park, H. & E. R. Hall. 1951. The gross anatomy of the tongues and stomachs of eight New World bats. Trans. Kansas Acad. Sci. 54:64-72. [ Links ]
Phillips C. J., G. W. Grimes & L. G. Forman. 1977. Oral Biology, Pp. 121-246, In: R. J. Baker, J.K. Jones Jr. and D. C. Carter, (Eds.) Biology of bats of the New World family Phyllostomatidae. Part II, Spec. Publ. Mus. Texas Tech Univ., 13. [ Links ]
Ramirez-Pulido J. I. Lira, S. Gaona, C. Müdespacher & A. Castro. 1989. Manejo y mantenimiento de colecciones mastozoológicas. Universidad Autónoma Metropolitana. 127 pp. [ Links ]
Reith, E. J. 1963. Type and formation of keratin in the rat's filiform papilla. Anat. Rec. 145:346. [ Links ]
Snow L. J., J. K. Jones Jr. & W. D. Webster, 1980. Centurio senex, Mammalian species, 138:1-3. [ Links ]
Suthers, R. A. 1970. Vision, olfaction and taste. Pp. 265-309, In: W. A. Wimsat (Ed.) Biology of bats Academic press. New York, 2:xii + 477. [ Links ]
Tamar, H. 1956. Taste responses at opossum and bat. Amer. J. Physiol. 187:636. [ Links ]
Uieda, W. 1986. Aspectos da morfología lingual das tres especies de morcegos hematofagos (Chiroptera, Phyllostomidae). Rio de Janeiro R. J., Rev. Brasil Biol. 46:581-587. [ Links ]
Uría, G. E. & M. C. Mora, 1996. Apuntes para el curso teórico-práctico de histología animal. Instituto Politécnico Nacional. México 1-277pp. [ Links ]
Wille, A. 1954. Muscular adaptation of the nectar eating bats (subfamily Glossophaginae). Trans. Kansas Acad. Sci., 57:315-325. [ Links ]

