










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa zoológica mexicana
versión On-line ISSN 2448-8445versión impresa ISSN 0065-1737
Acta Zool. Mex vol.20 no.2 Xalapa ago. 2004
Variación estacional del uso y preferencia de los tipos vegetacionales por el venado cola blanca en un bosque tropical de Jalisco
S. Mandujano1, S. Gallina1, G. Arceo1 & L. A. Pérez-Jiménez2
1 Departamento de Ecología y Comportamiento Animal, Instituto de Ecología A. C., Apartado Postal 63, Xalapa 91070, Veracruz, MÉXICO. E-mail: mandujan@ecologia.edu.mx.
2 Estación de Biología Chamela, Universidad Nacional Autónoma de México. Apartado Postal 70-233, México, D. F. 04510. MÉXICO.
Recibido: 25 de febrero 2002
Aceptado: 8 de diciembre 2003
RESUMEN
El venado cola blanca (Odocoileus virginianus) habita el bosque tropical de Chamela en la costa Pacífica del estado de Jalisco, México. En esta región la vegetación dominante es el bosque tropical caducifolio el cual se establece en las colinas y se caracteriza porque en el estrato arbóreo y sotobosque dominan especies que pierden sus hojas durante seis a siete meses en la época seca. En contraste, la vegetación que se encuentra a lo largo de los arroyos y ríos es el bosque tropical subperennifolio en el cual dominan especies con hojas no caducifolias. Por lo tanto, en esta región existe un contraste notable en la disponibilidad de alimento, fuentes de agua, cobertura contra el clima y depredadores, tanto temporal como espacialmente. El objetivo del presente estudio fue analizar la variación estacional del uso y la preferencia que el venado cola blanca tiene por las dos principales comunidades o tipos vegetales en Chamela. Los resultados se discuten en función de dos hipótesis las cuales predicen mayor preferencia del bosque tropical caducifolio durante la época de lluvias, y mayor preferencia del bosque tropical subperennifolio durante la época de secas. El estudio se realizó de 1989 a 1993 y se emplearon los métodos de conteo directo de venados activos en el día a lo largo del año y el conteo de grupos fecales durante la época seca. Los sotobosques de ambos tipos vegetacionales fueron descritos para cada época del año (lluvias y seca) cuantificando el número total de especies arbóreas, arbustivas y herbáceas, y se expresó como densidad promedio de número de especies por metro cuadrado; también se estimó la producción de biomasa foliar de las partes potencialmente consumibles por el venado como hojas y ramas tiernas, y se expresó como gramo de peso seco por metro cuadrado; finalmente se estimó la disponibilidad relativa de alimento a partir de datos de disponibilidad de las plantas que consume el venado en este sitio. Como resultado se encontró que en la época de lluvias la densidad de especies y la biomasa foliar fue mayor en comparación a la época seca (prueba T de rangos de Mann-Whitney, P < 0.01). En particular, en la época de lluvias la densidad de especies fue mayor en el bosque tropical caducifolio, mientras que en la época seca tanto la densidad de especies como la biomasa fue mayor en el bosque tropical subperennifolio (T, P < 0.01). También se encontraron diferencias significativas en la densidad de especies y biomasa foliar entre años (prueba H de Kruskal-Wallis, P < 0.01 ). Por lo tanto, hubo correlación positiva significativa entre la cantidad de lluvia con la densidad de especies y biomasa foliar de las especies en ambos tipos de bosques (r2, P < 0.001). Durante la época seca la disponibilidad relativa de alimento fue mayor en el bosque tropical caducifolio que en el bosque tropical subperennifolio (X2, P < 0.01). Respecto al uso del hábitat, se observó mayor número de venados durante el período de crianza (época húmeda de julio a octubre), menor durante el período de apareamiento (época de transición de noviembre a febrero), y un aumento durante el período de gestación (época seca de marzo a junio) (X2, P < 0.01). El venado usó ambos tipos de comunidades vegetales todo el año, pero prefirió el bosque tropical caducifolio durante la época de lluvias (X2, P < 0.01) como área de forrajeo, descanso y crianza, debido a que ofrece una mayor disponibilidad de alimento de alta calidad nutricional, y mayor protección contra depredadores. Durante la época seca el venado no prefirió el bosque tropical subperennifolio (X2, P > 0.05) no obstante que este tipo vegetacional ofrece mayor densidad de especies y biomasa foliar en el sotobosque en comparación al bosque tropical caducifolio. Esto podría deberse a que el bosque tropical subperennifolio tiene menor disponibilidad de las plantas que consume el venado, a que probablemente las plantas tengan un mayor contenido de fibra, y a que tiene mayor presencia de depredadores y cazadores. Por el contrario, no obstante la baja densidad de especies y biomasa foliar de las plantas en el bosque tropical caducifolio, el venado lo usó preferentemente en la época seca (X2, P < 0.01) debido a que tiene mayor cantidad de alimento, los árboles del ciruelo (Spondias purpurea) se establecen principalmente en este tipo de vegetación y sus frutos representan un recurso muy importante que le provee con agua al venado durante este periodo, la cobertura del sotobosque y la topografía disminuyen los riesgos de depredación, y las laderas con exposición norte tienen mejor cobertura termal siendo más húmedas.
Palabras Clave: bosque tropical caducifolio, bosque tropical subperennifolio, sotobosque, época de lluvias y de secas, uso y preferencia, variación temporal y espacial, venado cola blanca, Odocoileus virginianus, Chamela, México.
ABSTRACT
The white-tailed deer (Odocoileus virginianus) inhabits the tropical forest of Chamela in the Pacific coast of the state of Jalisco, Mexico. In this region the dominant vegetation is the tropical dry forest which settles down in the hills and it is characterized because in the arboreal stratum and understory dominate species that lose its leaves during six to seven months in the dry season. In contrast, the vegetation that is along the streams and rivers is the tropical semi-evergreen forest in which dominate species with leaves non deciduous. Therefore, in this region a remarkable contrast exists in the food availability, sources of water, covering against the climate and predators. The objective of the present study was to analyze the seasonal variation of the use and the preference that the white-tailed deer has for the two main communities in Chamela. The results are discussed in function of two hypothesis which predict bigger preference of the tropical dry forest during the rainy season, and bigger preference of the tropical semi-evergreen forest during the dry season. The study was carried out from 1989 to 1993, using the methods of direct count of active deer along the year and the count of fecal groups during the dry season. The understory of both plant communities were described for each season of the year quantifying the total number of arboreal species, shrubs and herbaceous, and it was expressed as density of the number of species for square meter; also was estimated the production of biomass to foliage potentially of the parts consumable for the deer like leaves and tender branches, and it was expressed as gram of dry weight by square meter; finally was estimated the relative availability of food in the dry season. As a result it was found that in the rainy season the density of species and the biomass of foliage was bigger in comparison to the dry season (T test of ranges of Mann-Whitney, P <0.01). In particular, in the rainy season the density of species was higher in the tropical dry forest, while in the dry season both the density of species and the biomass was higher in the tropical semi-evergreen (T, P <0.01). They were also significant differences in the density of species and biomass among years (H test of Kruskal-Wallis, P <0.01). Therefore, there was significant positive correlation among the quantity of rain with the density of species and biomass to foliage of the species in both types of forests (r2, P <0.001). During the dry season the relative availability of food was higher in the tropical dry forest than in the tropical semi-evergreen forest (X2, P <0.01). Regarding the use of the habitat, higher number of deer was observed during the period of fawning (July to October), smaller during the period of mating (November to February), and an increase during the period of gestation (March to June) (X2, P <0.01). The deer used both types of vegetable communities the whole year, but preferred the tropical dry forest during the rainy season (X2, P <0.01) as area of forage, rest and fawning, because it offers a higher availability of food of high quality nutritional, and bigger protection against predators. During the dry season the deer not prefer the tropical semi-evergreen forest (X2, P> 0.05) nevertheless that this type vegetacional offers higher density of species and biomass in the understory in comparison to the tropical dry forest. This could be due to that the tropical semi-evergreen forest has smaller availability of the plants that consumes the deer to that the plants probably have a high contained of fiber, and to that has bigger predators presence and hunters. On the contrary, nevertheless the drop density of species and biomass of the plants in the tropical dry forest, the deer used it preferably in the dry time (X2, P <0.01 ) because it has bigger quantity of food, the trees of the plum tree (Spondias purpurea) establish mainly in this type of vegetation and its fruits represent a important resource that provides with water to the deer during this period, the covering of the understory and the topography diminish the predation risks, and the hillsides with north exhibition have better thermal covering being more humid.
Key Words: tropical dry forest, tropical semi-evergreen forest, understory, wet and dry seasons, use-preference, temporal-spatial variation, white-tailed deer, Odocoileus virginianus, Chamela, Mexico.
INTRODUCCIÓN
Los ungulados seleccionan aquellas comunidades o tipos vegetacionales que les proveen con los recursos adecuados para satisfacer sus necesidades de manutención y reproducción, y que a su vez disminuyen el riesgo a condiciones climáticas severas y a la depredación (Sih 1993). En particular, el venado cola blanca (Odocoileus virginianus) está clasificado entre los rumiantes que seleccionan plantas con alto contenido celular de rápida fermentación y fácil digestión (Hanley 1982, 1997). Esto se debe a que el rumen de esta especie es pequeño en comparación a su cuerpo, por lo que debe compensar su menor capacidad ruminoreticular seleccionando plantas de alta calidad nutricional (Henke et al. 1988, Short 1963, Vangilder et al. 1982). Por lo tanto, el venado cola blanca prefiere consumir hojas y ramas jóvenes, plántulas, flores y frutos (Melchiors et al. 1985, Johnson et al. 1995). Como consecuencia de esta selección por determinadas especies y partes, este ungulado se caracteriza por usar determinadas comunidades vegetales y estados sucesionales (v.gr. Vangilder et al. 1982, Leslie et al. 1984, Ford et al. 1994). Además de estas restricciones en la selección del tipo de plantas para consumir y los tipos de comunidades vegetales donde tienen que buscar a éstas plantas, el venado debe resolver adecuadamente la variación estacional y espacial en la disponibilidad y calidad de las plantas (v.gr., Weckerly 1994). Esto es esencial para cubrir sus requerimientos nutricionales para manutención, crecimiento y reproducción los cuales cambian dependiendo de la edad, sexo, estado reproductivo y época del año (Hanley 1997). Por lo tanto, el venado modifica a lo largo del ciclo anual sus ritmos de actividad, desplazamientos y ámbito hogareño (Marchinton & Hirth 1984). Estos aspectos de la ecología del venado cola blanca, el uso y preferencia de hábitat, han sido ampliamente estudiados en hábitats templados y semiáridos tanto de Estados Unidos (v. gr., Larson et al. 1978, Armstrong et al. 1983, Loft & Menke 1984, Harestad 1985, Melchiors et al. 1985, Ordway & Krausman 1986), como de México (v. gr., Gallina 1994, Gallina et al. 1998a, 1998b, Soto-Werchitz 2000, Bello 2001). En contraste, son mínimos los estudios de estos aspectos en hábitats tropicales (v. gr., Irby & Calvopiña 1994). Esto es importante de conocer ya que el venado cola blanca habita en comunidades vegetacionales tropicales con una estacionalidad muy marcada en la disponibilidad de recursos.
El venado cola blanca habita los bosques tropicales de la región de Chamela en la costa del Pacífico del estado de Jalisco en México, con densidades de 12 individuos/km2 (Mandujano & Gallina 1993, 1995a). Estos bosques se caracterizan por un notable contraste en la fenología de las plantas a lo largo del año causado principalmente por la marcada estacionalidad en el régimen de lluvias (Rzedowski 1978). En esta región, la variación anual en la precipitación es influenciada por los vientos Alisios durante el verano y los ciclones durante el invierno (García-Oliva et al. 1991), lo cual origina una época húmeda y una época seca de 6 a 7 meses de duración (Bullock 1986). La precipitación, junto con las características topográficas y condiciones del suelo, determinan que en las laderas y cimas de las colinas domine el bosque tropical caducifolio, mientras que en las zonas donde corren arroyos y ríos domine el bosque tropical subperennifolio. Estas comunidades vegetales difieren, entre otros aspectos, en su composición florística, fenología y biomasa arbórea (Lott et al. 1987, Bullock & Solís-Magallanes 1990, Martínez-Yrizar et al. 1992). Además, la composición bromatológica de las plantas en el sotobosque varía notablemente dependiendo la época del año y el tipo de comunidad vegetal (Silva-Villalobos et al. 1999). Otra característica importante de la región de Chamela es que no existen arroyos que lleven agua durante todo el año, sino que éstos están restringidos a la época de lluvias, por lo que el venado debe satisfacer sus necesidades de este líquido aprovechando el rocío y la humedad contenida en las plantas y en los frutos (Mandujano & Gallina 1995b). En particular, los frutos del árbol Spondias purpurea (Anacardiaceae) son un recurso muy importante que le provee con agua al final de la época seca (Mandujano et al. 1994) y constituyen el 14% de la dieta durante este período (Arceo 2003). Los cambios en la vegetación influyen no solo sobre la disponibilidad de alimento y agua, sino en la cobertura de protección contra las temperaturas ambientales extremas (Mandujano & Gallina 1995b), y contra la presencia constante de depredadores como el puma (Puma concolor), jaguar (Panthera onca), ocelote (Leopardus pardalis) y coyote (Canis latrans) (López-González et al. 1997, Nuñez et al. 2000), además de la cacería furtiva.
Como consecuencia, el venado cola blanca debe enfrentar una marcada variación espacial (entre el bosque tropical caducifolio y el bosque tropical subperennifolio) y estacional (entre la época de lluvias y la época de secas) en la disponibilidad de alimento, agua y cobertura de protección contra el clima y depredadores. En particular, las condiciones adversas durante la época seca influyen sobre la probabilidad de mortalidad de los ungulados en hábitats tropicales (Owen-Smith 1990). Datos acerca de la dinámica poblacional del venado en Chamela indican que la tasa de fecundidad es relativamente baja debido a que no todas las hembras adultas tienen crías todos los años, y también debido a una menor sobrevivencia en la categoría de crías y juveniles (Mandujano & Gallina 2004). Por lo tanto, para incrementar su probabilidad de sobrevivencia los individuos deben seleccionar aquellos tipos de hábitats o comunidades vegetales que ofrezcan mejor disponibilidad y calidad de alimento y agua, y menor exposición a condiciones climáticas adversas y riesgo a ser depredados. El objetivo del presente estudio fue analizar la variación estacional del uso y la preferencia que el venado cola blanca tiene por las dos principales comunidades o tipos vegetales en Chamela. En particular, se analizan los resultados para tratar de probar dos hipótesis. Primero, se espera que durante la época húmeda los venados usen preferentemente el bosque tropical caducifolio debido a su mayor producción de plantas (potencialmente mayor alimento de alto valor nutritivo). Segundo, se espera que durante la época seca los venados usen preferentemente el bosque tropical subperennifolio debido a que presenta mayor biomasa vegetal (potencialmente mayor alimento y cobertura) en comparación al bosque tropical caducifolio. Para probar estas hipótesis, en el presente trabajo se analiza la variación estacional del uso y la preferencia que el venado tiene por ambas comunidades vegetales en relación con la variación en la riqueza de especies, la producción foliar, y el porcentaje de alimento relativo del sotobosque. Adicionalmente, se discute esto en relación a datos publicados acerca de la producción de frutos del S. purpurea (Mandujano et al. 1994, Mandujano, 200-), la disponibilidad de fuentes de agua durante la época seca (Mandujano & Gallina 1995b), la variación en el tamaño de los grupos del venado (Mandujano & Gallina 1996), el radioseguimiento de dos venados (Sánchez-Rojas et al. 1997), el papel de algunos depredadores (Nuñez et al. 2000) y de la chachalaca (Ortalis poliocephala) (Mandujano & Martínez-Romero 1997), el valor nutricional de las plantas (Silva-Villalobos et al. 1999), y los hábitos alimentarios del venado (Arceo 2003).
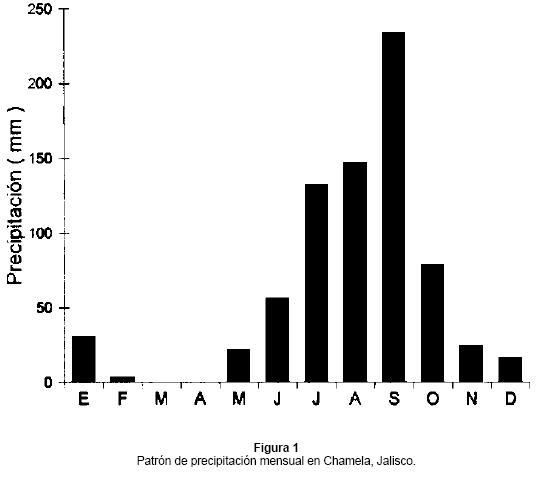
Área de estudio: El trabajo se realizó en la Estación de Biología "Chamela" de la UNAM, situada en la costa de Jalisco, México (19° 30' N y 105° 00' W). La Estación tiene una extensión de 3,200 ha y se caracteriza por una topografía irregular, con pendientes de 21° a 34°; el intervalo altitudinal es de 30 a 580 m, aunque la mayor parte del terreno no sobrepasa los 150 m. El clima es tropical con una marcada estacionalidad. La temperatura media anual es 25°C, con los meses más calientes entre Mayo y Septiembre (Bullock 1986). La precipitación promedio anual de 1977-1997 ha sido de 740 mm (SD = 196). La temporada de lluvias inicia la última semana de Junio, y de Julio a Octubre cae el 80% de la lluvia anual y tiene una duración media de 126 días (Fig. 1). Durante la época seca ocurre el 20% de la lluvia anual y tiene una duración media de 158 días (Bullock 1986). La precipitación mensual durante la época seca es muy variable, lo que indica que en el mismo mes se puede presentar una cantidad de precipitación diferente en distintos años. La variación anual en el patrón de precipitación está influenciado principalmente por los vientos Alisios y los ciclones del Pacífico (García-Oliva et al. 1991). Los primeros explican parte de la variabilidad anual de la lluvia que cae en Junio y Julio; mientras que los ciclones explican la variación de la lluvia entre Agosto y Octubre, además de tener gran influencia en la probabilidad de precipitación de Diciembre a Marzo. La vegetación dominante es el bosque tropical caducifolio el cual se localiza en lomeríos con suelos someros, el estrato arbóreo tiene una altura de 4 a 15 m, presenta un sotobosque bien desarrollado, y numerosas especies arbóreas y arbustivas pierden sus hojas durante la época seca. Algunas de las especies más comunes son el botoncillo (Cordia alliodora [Ruiz y Pav.] Oken), vara blanca (Lonchocarpus lanceolatus Benth.) e iguanero (Caesalpinia eriostachys Benth.). También se encuentra el bosque tropical subperennifolio el cual se desarrolla a lo largo de los arroyos principales, en los lugares protegidos sobre suelos profundos, y el estrato arbóreo tiene una altura de 10 a 25 m. Las especies más comunes son el culebro (Astronium graveolens Jacq.), mojote (Brosimum alicastrum Sw.) y tempisque (Sideroxylon capiri A. DC.) (Lott et al. 1987).
MÉTODOS
1. Descripción del sotobosque
Densidad de especies y producción foliar neta. Para describir la variación temporal y espacial del sotobosque, se obtuvieron índices de la densidad de especies (número de especies/m2) y de la producción foliar (gramos de peso seco/m2) del bosque tropical caducifolio y del subperennifolio, durante las épocas húmeda y seca. Los muestreos se hicieron al final de época seca (Mayo-Junio) de 1990 a 1993, y de la época húmeda (Octubre-Noviembre) de 1989 a 1992. En Octubre de 1989 se establecieron 50 parcelas permanentes (10 x 10 m cada una). Para seleccionarlas, primero se ubicaron en un mapa 120 puntos separados cada 50 m a lo largo de 6 km de caminos dentro del área de estudio. Luego se seleccionaron de manera aleatoria 25 puntos en el bosque tropical caducifolio y 25 en el subperennifolio. Con el fin de evitar posibles efectos de borde, las parcelas se ubicaron entre 10 y 20 m perpendicular a los caminos. Durante cada periodo de muestreo, en cada parcela se colocó una subparcela de 1 m2 en diferentes sitios dentro de la parcela permanente. En cada subparcela se cosecharon todas las plantas del sotobosque hasta 1.8 m lo cual representa la altura máxima que un venado puede alcanzar. De cada planta se cortaron únicamente las hojas y ramas jóvenes y se obtuvo el peso fresco para cada especie. Posteriormente las plantas se secaron durante 72 horas a temperatura constante (50 °C) y se obtuvo el peso seco y el porcentaje de humedad.
La densidad de especies y la producción foliar neta se expresaron y analizaron de tres maneras. Primero, la densidad se expresó como el promedio de especies por metro cuadrado, mientras que la biomasa como el promedio de gramos por metro cuadrado. En ambos casos se calculó para cada tipo vegetacional, cada época y año. Para detectar diferencias se aplicaron análisis de varianza de rangos de una vía no paramétricos empleando el estadístico H de Kruskal-Wallis considerando como factores a los tipos de vegetación (bosque tropical caducifolio y subperennifolio), épocas del año (húmeda y seca) y años (1989 a 1993). Se aplicó la prueba a posteriori de Dunn para comparaciones múltiples de las medias (Sokal & Rohlf 1995). En caso de comparaciones de dos grupos se utilizó el estadístico T para la prueba de suma de rangos de Mann-Whitney. Segundo, se analizó la distribución de frecuencia de parcelas en distintas categorías de densidad de especies y biomasa foliar para cada tipo vegetacional en ambas épocas del año. Tercero, se calculó la producción de biomasa foliar a partir de la biomasa en pie al final de la época seca y la biomasa al final de la época húmeda, y se expresó como gramos de peso seco por metro cuadrado producidos en cada tipo vegetacional. Por otro lado, para conocer si la densidad de especies y la biomasa foliar estacional dependen de la cantidad de lluvia, se realizaron análisis de regresión lineal simple para cada tipo de bosque. Los datos de precipitación mensual se obtuvieron de la estación meteorológica del área de estudio. Los datos mensuales se agruparon por época húmeda considerando de Junio a Octubre, mientras que los de la época seca corresponde de Noviembre a Mayo.
Disponibilidad relativa del alimento durante la época seca. Para conocer la cantidad de alimento que potencialmente puede ofrecer cada tipo vegetacional, se procedió a estimar la biomasa de las especies que consume el venado en este sitio. La información sobre la dieta se obtuvo a partir de la observación directa de venados cuando estaban comiendo y del análisis microhistológico de las heces (Arceo 2003). A partir de la información de biomasa foliar total, se separó la de las especies que consume el venado. Esta disponibilidad se expresó en cantidad absoluta (gramos de peso seco/m2) y cantidad relativa (biomasa consumible/biomasa total x 100). Estas estimaciones se hicieron para cada tipo vegetacional únicamente para las épocas secas de 1990 a 1993.
2. Uso y preferencia del hábitat por el venado
Obtención de datos. La información para analizar este aspecto se obtuvo a partir de los métodos de conteo directo de venados y del conteo de excrementos. Para el primer caso se colocaron de cinco a ocho transectos a lo largo de los caminos de terrecería (Mandujano & Gallina 1993). El largo total de los transectos varió entre 6 y 11 km (x = 9.1 km) durante cada mes de muestreo. Del largo total de los caminos, el 74% cruza por el bosque tropical caducifolio y el resto por el bosque tropical subperennifolio. Los transectos fueron recorridos (1 a 2 km/h) por una persona entre las 0700 y 1900 horas, de dos a cuatro veces cada mes. Cuando un venado era detectado se midió su distancia perpendicular al transecto. Posteriormente, todas las observaciones se incluyeron en cuatro clases de distancia perpendicular: 0-10, 11-20, 21-30 y 31-40 m. Cada vez que se detectaba un venado se tomaron los siguientes datos: sexo, edad (adulto, joven o cervatillo), hora, actividad, y tipo vegetacional (caducifolio o subperennifolio). Los datos se colectaron durante la época húmeda de 1989 a 1992, y la época seca de 1990 a 1993.
Para el conteo de grupos fecales en parcelas se establecieron aleatoriamente seis transectos de 400 m de largo con 40 parcelas de 9.3 m2 colocadas a intervalos de 10 m en cada parcela. El tiempo de depósito de los excrementos fue controlado limpiando previamente cada parcela de cualquier excremento viejo, y vuelta a revisar por dos o más personas 50 a 90 días después. Este método solo se aplicó durante la época seca de 1990 a 1993.
Análisis preliminar. Para poder emplear la información de los conteos directos como un indicador del uso del hábitat, primero se hicieron análisis de datos con la finalidad de eliminar posibles fuentes de error en la interpretación del uso de los tipos vegetacionales. Esto se debe a que la tasa mensual de observación de venados (n) entre el número total de kilómetros recorridos (L) ese mismo mes, puede variar dependiendo de dos factores: 1) no se recorrieron exactamente el mismo número de kilómetros cada mes; y 2) las distancias perpendiculares de los venados al transecto variaron cada mes dependiendo de la visibilidad de la vegetación la cual varía dependiendo el estado fenológico de las plantas el cual cambia de la época de lluvias a la época de secas.
Para el primer factor, los datos de observación mensual de venados se agruparon en tres épocas: lluvias (Julio a Octubre), transición (Noviembre a Febrero) y seca (Marzo a Junio). Estas tres épocas corresponden aproximadamente a los períodos de crianza, apareamiento y gestación (en el caso de hembras), respectivamente (Mandujano 1999). Para cada época, se empleó un análisis de regresión lineal simple para conocer si el número de venados detectados dependió del número de kilómetros recorridos. En caso de no significancia se puede concluir que el largo del transecto no influye en la cantidad de animales observados y por lo tanto se puede emplear las observaciones como un indicador del uso del hábitat por los venados. Para el segundo factor, se prosiguió a conocer si existían diferencias en la frecuencia de venados observados en cada clase de distancia perpendicular (0-10, 11-20, 2130 y 31-40 m) entre las tres épocas del año, empleando un análisis de heterogeneidad con el estadístico X2 (Sokal & Rohlf 1995). Diferencias significativas entre las épocas podrían implicar variación en la visibilidad impuesta por cambios fenológicos de la vegetación en el sotobosque. En este caso, es necesario solo incluir en el análisis de uso del hábitat a los venados detectados a determinada distancia. La visibilidad del sotobosque se ha estimado en un promedio de 9.9 metros en la época de lluvias y de 23.4 m en la época seca (Mandujano 1999).
Uso de los tipos vegetacionales. El uso de hábitat fue definido como el número de venados observados en cada tipo vegetacional durante cada época del año. Con los resultados de los análisis previos se decidió a emplear únicamente las observaciones de venados detectados entre 0 y 10 metros perpendiculares al transecto. Además, las observaciones se agruparon en períodos de 0900-1200 y 1600-1900 horas. A partir de esto se analizó la frecuencia de venados activos en cada tipo vegetacional, en cada período del día y durante cada época del año, por medio de una prueba de independencia de tres factores (Sokal y Rohlf 1995). Por otra parte, la información obtenida con el método de conteo de excrementos, se analizó considerando que el 73% de las parcelas fueron establecidas en el bosque tropical caducifolio y el 27% en el subperennifolio, lo cual corresponde aproximadamente a la superficie que abarca cada tipo vegetacional en el área de estudio. Además, las parcelas establecidas en el bosque caducifolio fueron clasificadas en función de su fisionomía (cima o ladera) y exposición (NE-NO o SE-SO). Se consideró como ladera si la parcela tenía más de 10 E de inclinación. Para estimar la preferencia, se empleó el análisis de uso-disponibilidad empleando la prueba X2 y los intervalos de Bonferroni al 95% de confianza (Byers et al. 1984).
RESULTADOS
1. Descripción de los tipos vegetacionales
Densidad de especies. El promedio de densidad de especies en el bosque tropical caducifolio fue de 9 ± 1 y 2 ± 0.2 especies/m2 durante las épocas húmeda y seca, respectivamente. Esta diferencia fue estadísticamente significativa (T = 2478, P = 0.0001). El 87% de las parcelas tuvieron entre 4 y 11 especies/m2 durante la época húmeda, y el 93% tuvieron entre 1 y 3 especies/m2 en la época seca (Fig. 2). Por su parte, el promedio de la densidad especies en el sotobosque del bosque tropical subperennifolio fue de 8 ± 1 y 4 ± 0.5 especies/m2. Esta diferencia fue significativa (T = 2867, P = 0.0001). El 90% de la parcelas tuvieron también de 4 a 11 especies/m2 en la época húmeda, y el 92% de 1 a 6 especies/m2 en la época seca (Fig. 2). La densidad de especies fue significativamente mayor en el bosque tropical caducifolio durante la época húmeda (T = 10937, P = 0.01); mientras que en la época seca la riqueza fue mayor en el bosque tropical subperennifolio (T = 6192, P = 0.0001) (Fig. 3). Por otro lado, hubo correlación positiva significativa entre la lluvia y la densidad de especies del bosque tropical caducifolio (r2 = 0.65, F = 11.1, P = 0.02) y del bosque tropical subperennifolio (r2 = 0.81, F = 25.1, P = 0.002). Del total de especies registradas en las parcelas de muestreo, se encontró que durante la época húmeda el 39% de las especies fueron exclusivas del bosque tropical caducifolio, el 43% del bosque tropical subperennifolio y el 18% se encontraron en ambos tipos de bosque.

Biomasa foliar en el sotobosque. En el bosque tropical caducifolio el promedio fue de 41.2 ± 1.9 y 7.1 ± 2.3 g/m2 durante las época húmeda y seca, respectivamente. Estas diferencias fueron altamente significativas (T = 3160, P = 0.0001). El 87% de las parcelas tuvieron entre 5 y 70 g/m2 en la época húmeda, y el 91% entre 0 y20 g/m2 en la época seca (Fig. 4). Las familias que contribuyeron con el 52% del total de biomasa en la época húmeda fueron Euphorbiaceae, Leguminosae, Graminae, Sapindaceae, Acantáceas y Malphigiaceae. De estas, Euphorbiaceae contribuyó con el 28.9% de la biomasa en el sotobosque, específicamente el género Acalypha fue el más importante. Durante la época seca Capparidaceae y Acanthaceae fueron las familias más importantes. La biomasa foliar neta en el bosque tropical subperennifolio se estimó en 41.0 ± 1.9 y 18.0 ± 2.3 g/m2 durante las épocas húmeda y seca respectivamente. Estas diferencias fueron significativamente diferentes (T = 3161, P = 0.0001). El 89% de las parcelas tuvieron entre 11 y70 g/m2 en la época húmeda, y el 94% entre 10 y 40 g/m2 en la época seca (Fig. 4). Las familias que contribuyeron con el 38% de la biomasa en la época húmeda fueron Leguminosae, Euphorbiaceae, Nyctaginaceae, Bignoniaceae, Phytolacaceae, Sapindaceae, Rutaceae y Graminae. Mientras que en la época seca más del 50% de la biomasa fue de Bignonaceae, Capparidaceae y Sapindaceae. Por otro lado, durante la época húmeda no hubo diferencias en la biomasa de ambos tipos de bosques (T= 9848, P= 0.89) (Fig. 5). En contraste, durante la época seca la biomasa del bosque tropical subperennifolio fue significativamente mayor a la del bosque tropical caducifolio (T = 5783, P = 0.0001). De finales de la época seca a la época húmeda hubo un incremento promedio de 341 kg/ha en el bosque tropical caducifolio y de 230 kg/ha en el bosque tropical subperennifolio. Por otro lado, hubo correlación positiva significativa entre la lluvia y la biomasa del bosque caducifolio (r2 = 0.87, F = 38.7, P = 0.001) y subperennifolio (r2 = 0.78, F = 21.3, P = 0.004).

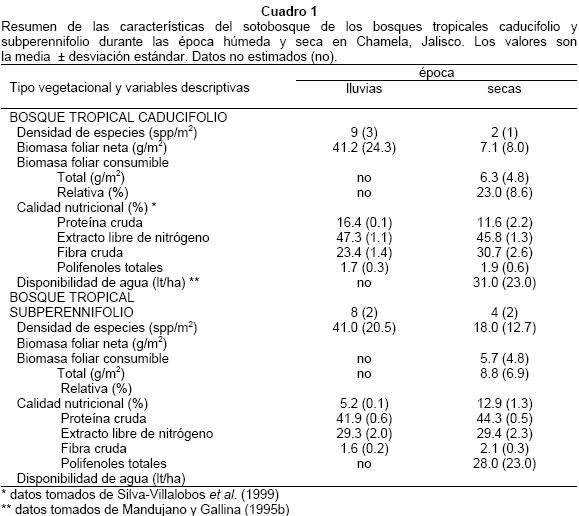
Disponibilidad relativa de alimento en la época seca. En el bosque tropical caducifolio la disponibilidad de biomasa foliar de las especies que consume el venado varió entre 2y12 g/m2, lo que en promedio corresponde al 23% de la biomasa foliar total en el sotobosque (Fig. 6). Para el bosque tropical subperennifolio se estimó una disponibilidad entre 1 y 10 g/m2, lo que corresponde al 9% de la biomasa foliar total. En el Cuadro 1 se presenta un resumen de las características del sotobosque durante las épocas húmeda y seca en los bosques tropicales caducifolio y subperennifolio. Además, se presenta información adicional acerca del valor nutritivo de las plantas y disponibilidad de agua.

2. Actividad y uso del hábitat por el venado
Se contaron un total de 299 venados en 691 km recorridos durante los cuatro años del estudio. Mensualmente, se recorrieron en promedio 19.7 km (rango de 10.7 a 25.3 km) durante 35 meses de muestreo. Aunque el número de venados observados tendió a aumentar conforme más kilómetros se recorrieron, esta relación no fue significativa para las épocas húmeda (r2 = 0.07, F = 0.06, P = 0.81), de transición (r2 = 0.18, F = 1.85, P = 0.21) y de secas (r2 = 0.17, F = 2.05, P = 0.18); ya que para similar número de kilómetros recorrido el número de venados detectados varió considerablemente. Por otro lado, del 50% al 70% de los venados se observaron entre los 0 y 10 metros perpendiculares al centro del transecto. El número de detecciones disminuyó conforme se incrementó la distancia perpendicular al centro del transecto, con solo el 2.5% al 10% de venados observados en la categoría de distancia más distante. La frecuencia de venados en cada clase de distancia perpendicular varió significativamente entre las épocas del año (X2 = 14.0, P = 0.03). En la época de lluvias se detectaron más venados en la clase cercana al centro del transecto, mientras que para la época seca el número de venados observados a distancias más lejanas se incrementó.
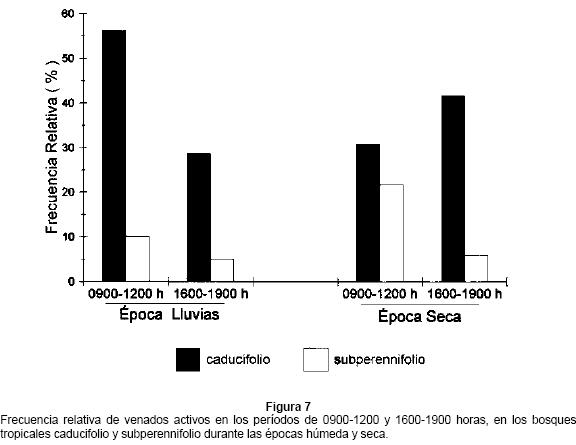
Con base en el número de venados observados, se encontró que el bosque tropical caducifolio fue significativamente más usado que el bosque tropical subperennifolio (X2 = 15, P = 0.01). En particular, el bosque tropical caducifolio fue más usado durante las mañanas (0900-1200 h) en la época húmeda, mientras que en la época seca fue más usado en la tarde (1600-1900 h) (X2 = 120, P = 0.001). Por su parte, el bosque tropical subperennifolio fue más usado en las mañanas durante la época seca y muy poco usado en la época húmeda (Fig. 7).
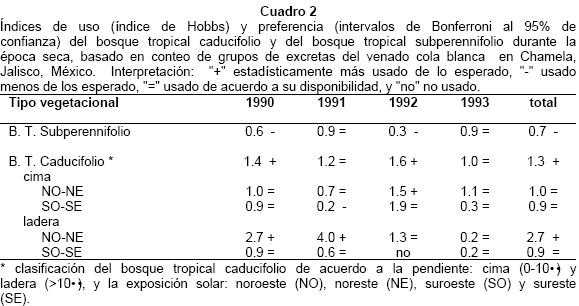
Se contaron un total de 422 grupos fecales en 960 parcelas. Durante la época seca, el bosque tropical caducifolio fue más usado que el bosque tropical subperennifolio (Cuadro 2, X2= 14.7, P = 0.001). En particular, en 1990 y 1992, el bosque tropical caducifolio fue más usado de lo esperado; mientras que en 1991 y 1993 ambos tipos vegetacionales fueron usados de manera similar. Por otro lado, las laderas con exposición norte fueron preferidas (X2 = 26.6, P =0.001).
DISCUSIÓN
Los resultados apoyan la primera hipótesis planteada la cual predecía mayor preferencia del venado cola blanca por el bosque tropical caducifolio durante la época húmeda. Esto podría deberse a que en esta época este tipo vegetacional ofrece mayor riqueza y producción de biomasa de especies de las familias Euphorbiaceae, Convolvulaceae, Leguminosae y Sapindaceae, y de algunos géneros que el venado prefiere en su dieta como Acalypha e Ipomoea (Arceo 2003). Además de que este tipo vegetacional presenta plantas de mayor valor nutricional (alto porcentaje de proteína y extracto libre de nitrógeno, y bajo en fibra y polifenoles) en comparación al bosque tropical subperennifolio (Silva-Villalobos et al. 1999). Particularmente, en la época de lluvias las plantas están activamente creciendo por lo que invierten más energía al tejido de crecimiento (hojas y ramas jóvenes) y reproductivo (flores y frutos), que al tejido estructural como son las partes leñosas las cuales tienen mayor porcentaje de fibra. Como consecuencia, durante la fase de crecimiento las plantas tienen mayor cantidad de compuestos nitrogenados, mayor digestibilidad y menor contenido de fibra y lignina (Short et al. 1974, Blair et al. 1977, Mattson 1980), por lo que son muy palatables para un herbívoro como el venado cola blanca (Hanley 1982). Esta debe ser la razón por la que el venado selecciona en su dieta un mayor número de especies que se establecen principalmente en el bosque tropical caducifolio, ya que de 109 especies registradas en la dieta anual de este herbívoro, el 85% se encuentran principalmente en el bosque tropical caducifolio (Arceo 2003). Además, la preferencia por este tipo vegetacional es importante pues durante la época de lluvias las hembras tienen a sus crías y los machos están desarrollando astas nuevas para competir con otros machos por hembras en estro durante la época de apareamiento, por lo que la demanda de energía y nutrientes es muy alta (French et al. 1956, Thompson et al. 1973, Ullrey et al. 1973). Los resultados del radioseguimiento de una hembra por un periodo de dos años también sugiere mayor preferencia por el bosque tropical caducifolio (Sánchez-Rojas et al. 1997).
Por el contrario, la segunda hipótesis planteada predecía que el venado cola blanca prefiere el bosque tropical subperennifolio como área de resguardo y forrajeo durante la época seca, sin embargo los resultados no la sustentan. Es interesante notar que no obstante que este tipo vegetacional presenta mayor riqueza de especies y biomasa foliar de las plantas en comparación al bosque tropical caducifolio en esta época, no se encontró evidencia de ramoneo intensivo por parte del venado. Esto sugiere que el bajo valor nutricional de las plantas del bosque tropical subperennifolio tiene un papel importante para limitar su uso por el venado. Durante esta época decrece el contenido de proteína y extracto libre de nitrógeno, y aumenta el contenido de fibra (Silva-Villalobos et al. 1999). Se sabe que a medida que maduran las plantas aumenta la fibra contenida en sus paredes celulares, por lo que disminuye la digestibilidad y palatabilidad de las mismas (Short et al. 1974). Por lo tanto, es posible que la estrategia de forrajeo del venado durante la época seca incluya evitar plantas con alto contenido de fibra (tal vez lignina) y compuestos químicos tóxicos del bosque tropical subperennifolio; además, de incrementar el consumo de especies poco disponibles en el bosque tropical caducifolio (v.gr., Lundberg & Astrom 1990). En particular, es posible que debido a que muchas especies del bosque tropical subperennifolio mantienen sus hojas durante la época seca, su contenido de lignina en las paredes celulares podría ser mayor respecto a las plantas del bosque caducifolio. Se ha documentado que la lignina es un compuesto que no puede ser aprovechado por los rumiantes (Vangilder et al. 1982) y que es contenido en alto porcentaje en las plantas perennes (Janzen & Waterman 1984). Por lo tanto, el bosque tropical subperennifolio tendría menor disponibilidad de alimento para el venado durante la época seca, en comparación al bosque tropical caducifolio. Esto podría explicar la alta diversidad de familias y especies en la dieta de este herbívoro en la época seca (Arceo 2003). Por otro lado, se espera que se incremente la agregación de mamíferos herbívoros en hábitats con variación sustancial en la calidad y producción de forraje (Fryxell 1991), la única asociación que se observa con más frecuencia en el área de estudio durante la época seca, es el de hembras adultas con sus crías y es más común observar a individuos solitarios (Mandujano & Gallina 1996). Ocasionalmente se observó agregación de venados de ambos sexos asociado al consumo de frutos de árboles como Ficus spp. y Brosimum alicastrum en el bosque tropical subperennifolio; sin embargo, el mayor uso lo recibió el bosque tropical caducifolio en esta época. Nuevamente, el radio seguimiento de una hembra durante dos años, sugiere que prefiere el bosque tropical caducifolio en la época seca (Sánchez-Rojas et al. 1997).
En particular, dentro del bosque tropical caducifolio el venado prefirió las colinas con exposición norte durante la época seca, lo cual podría interpretarse como otra estrategia para obtener alimento, conservar el agua o protegerse de depredadores. Estas laderas son más frescas (Galicia 1992), presentan una mayor acumulación de rocío entre noviembre y marzo (González 1992) y los árboles de Spondias purpurea son comunes en estos sitios (Mandujano et al. 1994). En las cimas de las colinas es muy común encontrar echaderos de los venados y observar rastros de hembras adultas con crías. Por lo tanto, las cimas y laderas norte posiblemente provean al venado con cobertura termal de protección, con mayor alimento y agua durante la época seca. Por otra parte, la alta variación en la riqueza y biomasa foliar por metro cuadrado aunado a la baja calidad de las plantas en el bosque tropical caducifolio durante la época seca, podría tener por lo menos dos consecuencias respecto a la estrategia de forrajeo de este ungulado. La primera es que los venados disminuyan la ingestión de alimento y bajen su metabolismo a través de una reducción del tiempo de actividad y las distancias desplazadas, como ha sido documentado en otros estudios (Hanley 1997). La segunda consecuencia es que el venado incremente el consumo de frutos y flores de alto valor nutricional (Ford et al. 1993, Johnson et al. 1995). Los resultados apoyan ambas posibilidades ya que los desplazamientos de los venados con radiocollares, aunque no es representativo de la población por el escaso número de individuos seguidos, sugieren que los venados se mueven menos durante la época seca (Sánchez-Rojas et al. 1997). Además, estos movimientos están asociados con un incremento en la diversidad de familias y especies en la dieta, y consumen hasta el 30% de frutos y flores (Arceo 2003).
Durante la época seca no hay fuentes de agua libre en Chamela, por lo que el venado aprovecha la humedad contenida en las plantas y los frutos (Mandujano & Gallina 1995b). En este sentido, los frutos de S. purpurea son un recurso clave para el venado (Mandujano et al. 1994). Por lo tanto, la distribución y abundancia de los árboles con frutos de S. purpurea podría ser un factor para explicar porque el venado usa preferentemente el bosque caducifolio durante la época seca. Esto se debe a que casi todos los árboles de esta especie se establecen en este tipo vegetacional. En particular, el modo de consumo de los frutos de éste árbol por la chachalaca (Ortalis poliocephala) pudiera tener consecuencias importantes en los movimientos de los venados en este sitio. Esto se debe a que esta ave consume los frutos de S. purpurea y en este proceso tira muchos frutos de las ramas, haciéndolos disponibles para que el venado los consuma en el suelo. Resultados de un experimento de exclusión mostró que en ausencia de esta ave la tasa de caída de los frutos es cinco veces menor (Mandujano & Martínez-Romero 1997). Sin embargo, los frutos no se encuentran de manera masiva en el suelo. Esto es importante para el venado pues estos frutos son consumidos además por otros mamíferos entre los que destaca el pecarí de collar (Pecari tajacu) (Mandujano et al. 1994, Martínez-Romero & Mandujano 1995, Mandujano & Martínez-Romero, en prensa). Por lo tanto, los frutos de S. purpurea en el suelo son un recurso muy competido por diferentes especies, por lo tanto los venados deben trasladarse rápidamente de un árbol a otro para consumirlos.
El riesgo a la depredación debe ser otro factor que contribuye a que los venados prefieran durante todo el año el bosque tropical caducifolio. Se ha estimado que la presión de depredación por el ocelote y coyote es del 12% (López-González et al. 1997), mientras que la depredación por pumas y jaguares se ha estimado en 25% (Nuñez et al. 2000) de la población de venado en este sitio. Además, estos depredadores y cazadores provenientes de las poblaciones humanas aledañas, usan los arroyos (donde se establece el bosque tropical subperennifolio) como rutas en sus recorridos de búsqueda (Núñez & Miller 1997). Por lo tanto, para el venado forrajear en el bosque tropical subperennifolio representa un sitio de alto riesgo. En este sentido, se considera que la variación en el tamaño de las manada como una estrategia antidepredatoria (Hirth 1977, Nelson & Mech 1981). En el área de estudio, el venado no forma manadas grandes sino que es más común observar a venados solitarios y a hembras con sus crías (Mandujano & Gallina 1996). Esto es muy importante ya que los cervatillos durante la época de lluvias son muy susceptibles a la depredación (Cook et al. 1971), en particular por el ocelote en el área de estudio (López-González et al. 1997). Esto sugiere que en Chamela, similar a otros sitios (Nelson & Mech 1981, Sih 1993), la probabilidad de depredación influye en el uso y preferencia del hábitat por el venado.
Conclusiones La heterogeneidad del sotobosque de los bosques tropicales caducifolio y subperennifolio de Chamela, está determinada por la variación estacional y anual en la composición florística, la densidad de especies, la biomasa foliar, el valor nutricional, el porcentaje de especies con hojas deciduas y perennes y, posiblemente, estrategias antiherbivoría. Esta heterogeneidad espacial y temporal del sotobosque tiene consecuencias en términos del patrones de uso y preferencia de hábitat por el venado cola blanca. No obstante la alta densidad de especies y biomasa foliar de las plantas durante todo el año en el bosque tropical subperennifolio, el venado no lo utiliza preferentemente como área de forrajeo. Por el contrario, el venado prefiere el bosque tropical caducifolio durante todo el año. Una posible causa de la menor utilización del bosque tropical subperennifolio sea la mayor depredación, aspecto que debe investigarse con más detalle. Otro factor que debe influir en el uso y preferencia del hábitat es la baja disponibilidad de fuentes de agua en la época seca, lo que determina que el venado busque fuentes alternativas como por ejemplo el agua contenida en los frutos del ciruelo, Spondias purpurea. Por otro lado, la competencia con otros mamíferos como el pecarí de collar no debe ser un factor importante en el uso del hábitat dadas las diferencias en las preferencias alimenticias de ambos ungulados. Sería altamente recomendable el empleo de la técnica de radiotelemetría con un mayor número de animales marcados, con el fin de tener información más precisa acerca de los patrones de actividad, desplazamiento y uso del hábitat.
AGRADECIMIENTOS
Le agradecemos a L. E. Martínez-Romero, G. Sánchez-Rojas, G. Silva-Villalobos, R. E. Sánchez-Mantilla, D. Valenzuela, G. Zavala y G. González su ayuda en algunos de los muestreos. Las siguientes personas colaboraron en la identificación de ejemplares en el herbario: F. Chiang (todos los grupos), M. Sousa (Leguminosae), O. Tellez (Leguminosae) y J. L. Villaseñor (Compositae). Apreciamos los comentarios de R. Putman a un manuscrito inicial de la sección sobre la descripción de la vegetación y a J. Laundré sobre la sección del uso del hábitat. El presente estudio recibió apoyo de CONACYT (P220CC0R-892154, P020CCOR-903703 y 0327N9107) y SEP (DGICSA-902467). Además, CONACYT proporcionó becas para estudios de posgrado a S. Mandujano y G. Arceo. La Estación de Biología Chamela de la UNAM brindó todas las facilidades y días de apoyo para la realización del estudio.
LITERATURA CITADA
Arceo, G. 2003. Hábitos alimentarios del venado cola blanca (Odocoileus virginianus) en el bosque tropical caducifolio de Chamela, Jalisco. Tesis de Maestría, Facultad de Ciencias, UNAM. México, D.F. [ Links ]
Armstrong, E., D. Euler & G. Racey. 1983. White-tailed deer habitat and cottage development in central Ontario. J. Wildl. Manage. 47:605-612. [ Links ]
Bello, J. 2001. Comportamiento del venado cola blanca texano en sitios con distintos manejo del agua en el noreste de México. Tesis Doctoral, Instituto de Ecología A. C., Ver. [ Links ]
Blair, R. M. & L. E. Brunnet. 1980. Seasonal browse selection by deer in a southern pine-hard-wood habitat. J. Wildl. Manage. 44:79-88. [ Links ]
Bullock, S. H. 1986. Climate of Chamela, Jalisco, and trends in south coastal region of Mexico. Arch. Metereo. Geoph. and Bioclim. 36:297-316. [ Links ]
Bullock, S. H. & J. A. Solís-Magallanes. 1990. Phenology of canopy trees of a tropical deciduous forest in Mexico. Biotropica 22:22-35. [ Links ]
Byers, C. R., R. K. Steinhosrt & P. R. Krusman. 1984. Clarification of a technique for analysis of utilization-availability data. J. Wildl. Manage. 48:1050-1052. [ Links ]
Cook, R. S., M. White, D. O. Trainer & W. C. Glazener. 1971. Mortality of young white-tailed deer fawns in south Texas. J. Wildl. Manage. 35:47-56. [ Links ]
Ford, W. M., A. S. Johnson & P. E. Hale. 1993. Yellow-poplar flowers in the spring diet of white-tailed deer in the Southern Appalachians. J. Tenn. Acad. Scien. 68:56. [ Links ]
----------. 1994. Nutritional quality of deer browse in southern Appalachian clearcuts and mature forests. Forest. Ecol. Manage. 67:149-157. [ Links ]
French, C. E., L. C. McEwen, N. D. Magruder, R. H. Ingram & R. W. Swift. 1956. Nutrient requirements for growth and antler development in the white-tailed deer. J. Wildl. Manage. 20:221-232. [ Links ]
Fryxell, J. M. 1991. Foraging quality and agregation by large herbivores. Am. Nat. 138:478-498. [ Links ]
Galicia, L. 1992. Influencia de la variabilidad de la forma de la pendiente en las propiedades físicas del suelo y su capacidad de retención de agua, en una cuenca tropical estacional. Tesis Licenciatura, Universidad Nacional Autónoma de México. México, D. F. [ Links ]
Gallina, S. 1994. Uso del hábitat por el venado cola blanca en la reserva de la biosfera La Michilia, México. Pp. 299-314. In: Ch. Vaughan and M. A. Rodríguez (Eds.) Ecología y Manejo del Venado Cola Blanca en México y Costa Rica. EUNA, Heredia, Costa Rica. [ Links ]
Gallina, S., S. Mandujano, J. Bello & C. Delfín. 1998. Home-range size of white-tailed deer in northeastern Mexico. Pp. 47-50. In: J. C. de Vos, Jr. (Ed.), Deer & Elk Workshop, Fish & Wildlife Department, Rio Rico, Arizona. [ Links ]
Gallina, S., A. Pérez-Arteaga & S. Mandujano. 1998. Patrones de actividad del venado cola blanca (Odocoileus virginianus texanus) en un matorral xerófilo de México. Bol. Soc. Biol. Concepción Chile 69:221-228. [ Links ]
García-Oliva, F., E. Ezcurra & L. Galicia. 1991. Pattern of rainfall distribution in the central Pacific coast of Mexico. Geog. Ann. 73:179-186. [ Links ]
González, M. G. 1992. Importancia ecológica del rocío en la selva baja caducifolia de Chamela, Jalisco. Tesis Licenciatura, Universidad Nacional Autónoma de México, México, D. F. [ Links ]
Hanley, T. A. 1982. The nutritional basis for food selection by ungulates. J. Range. Manage. 28:298-300. [ Links ]
----------. 1997. A nutritional view of understanding and complexity in the problem of diet selection by deer (Cervidae). Oikos 79:209-218. [ Links ]
Harestad, A. S. 1985. Habitat use by black-tailed deer on northern Vancouver island. J. Wildl. Manage. 49:946-950. [ Links ]
Henke, S. E., S. Demarais & J. A. Pfister. 1988. Digestive capacity and diets of white-tailed deer and exotic ruminants. J. Wildl. Manage. 52:595-598. [ Links ]
Hirth, D. H. 1977. Social behavior of white-tailed deer in relation to habitat. Wildl. Monog. 53:1-55. [ Links ]
Irby, L. R. & J. Calvopiña. 1994. Uso de hábitat por el venado cola blanca reintroducido en la península de Nicoya, Costa Rica. Pp. 333-347. In: Ch. Vaughan and M. A. Rodríguez (Eds.) Ecología y Manejo del Venado Cola Blanca en México y Costa Rica. EUNA, Heredia, Costa Rica. [ Links ]
Janzen, D. H. & P. G. Waterman. 1984. A seasonal census of phenolics, fibre and alkaloids in foliage of forests trees in Costa Rica: Some factors influencing their distribution and relation to host selection by Sphingidae and Saturniidae. Biol. J. Linn. Soc. 21: 439-454. [ Links ]
Johnson, A. S., P. E. Hale, W. M. Ford, J. M. Wentworth, J. R. French, O. F. Anderson & G. B. Pullen. 1995. White-tailed deer foragin in relation to successional stage, overstory type and management of southern Appalachian forests. Ame. Midl. Natur. 133:18-35. [ Links ]
Larson, T. J., O. J. Rongstad & F. W. Terbilcox. 1978. Movement and habitat use of white-tailed deer in southcentral Wisconsin. J. Wildl. Manage. 42:113-117. [ Links ]
Leslie, D. M., E. E. Starkey & M. Vavra. 1984. Elk and deer diets in old-growth forests in western Washington. J. Wildl. Manage. 48:762-775. [ Links ]
Loft, E. R. & J. W. Menke. 1984. Deer use and habitat characteristics of transmission-line corridors in a douglas-fir forest. J. Wildl. Manage. 48:1311-1316. [ Links ]
López-González, C., M. G. Hidalgo-Mihart & A. González-Romero. 1997. White-tailed deer pr edation by mammalian carnivores in tropical dry forests of western Mexico. Proc. Western States & Provinces Deer and Elk Workshop. Rio Rico, AZ. [ Links ]
Lott, E.J., S. H. Bullock & J. A. Solís-Magallanes, J. A. 1987. Floristic diversity and structure of upland and arroyo forests in coastal Jalisco. Biotropica 19:228-235. [ Links ]
Lundberg, P. & M. Astro'm. 1990. Low nutritive quality as a defense againts optimally foraging herbivores. Am. Nat. 135:547-562. [ Links ]
Mandujano, S. 1999. Ecología del venado cola blanca en un bsoque tropical caducifolio de Jalisco. Tesis Doctoral, Universidad Nacional Autónoma de México. México, D.F. [ Links ]
----------. 200-. Ciruelo, Spondias purpurea L. (Anacardeaceae). In: F. Noguera, J. Vega y R. Ayala (Eds.). Historia Natural de Chamela. Universidad Nacional Autónoma de México. México, D.F. [ Links ]
Mandujano, S. & S. Gallina. 1993. Estimaciones de la densidad del venado cola blanca basadas en conteos en transectos en un bosque tropical de Jalisco. Acta Zool. Mex. (n.s.) 56:1-37. [ Links ]
----------. 1995a. Comparison of deer censusing methods in tropical dry forest. Wildl. Soc. Bull. 23:180-186. [ Links ]
----------. 1995b. Disponibilidad de agua para el venado cola blanca en un bosque tropical caducifolio de México. Vida Silvestre Neotropical 4:107-118. [ Links ]
----------. 1996. Size and composition of white-tailed deer groups in a tropical dry forest in Mexico. Ethol. Ecol. & Evol. 8:255-263. [ Links ]
----------. 200-. Dinámica poblacional del venado cola blanca en un bosque tropical caducifolio de Jalisco. In: V. Sánchez-Cordero y R. Medellín (eds.), Homenaje al Doctor Bernardo Villa. Universidad Nacional Autónoma de México. México, D.F. [ Links ]
Mandujano, S. & L. E. Martínez-Romero. 1997. Fruit fall caused by chachalacas (Ortalis poliocephala) on red mombin trees (Spondias purpurea): impact on terrestrial fruit consumers, especially the white-tailed deer (Odocoileus virginianus). Stud. Neotrop. Fauna & Environm. 32:1-3. [ Links ]
----------. 200-. Pecarí de collar, Pecari tajacu. In: F. Noguera, J. Vega y R. Ayala (Eds.). Historia Natural de Chamela. Universidad Nacional Autónoma de México. México, D.F. [ Links ]
Mandujano, S., S. Gallina & S. H. Bullock. 1994. Frugivory and dispersal of Spondias purpurea (Anacardiaceae) in a tropical deciduous forest in Mexico. Rev. Biol. Trop. 42:105-112. [ Links ]
Marchinton, R. L. & D. H. Hirth. 1984. Behavior. Pp. 129-168, In: L. K. Halls (Ed.). White-tailed deer: ecology and management. Stackpole Bokks, Harrisburg, PA. [ Links ]
Martínez-Romero, L. E. & S. Mandujano. 1995. Hábitos alimentarios del pecarí de collar (Pecari tajacu) en un bosque tropical de Jalisco. Acta. Zool. Mex. (n. s.) 70:1-30. [ Links ]
Martínez-Yrizar, A., J. Sarukhán, A. Pérez-Jiménez, E. Rincón, J. M. Mass, J. A. Solís-Magallanes & L. Cervantes. 1992. Above-ground phytomass of a tropical deciduous forest on the coast of Jalisco, Mexico. J. Trop. Ecol. 8:87-96. [ Links ]
Mattson, W. J., Jr. 1980. Herbivory in relation to plant nitrogen content. Ann. Rev. Ecol. Sys. 11:119-161. [ Links ]
Melchiors, M. A., T. H. Silker & J. E. Reeb. 1985. Deer use of young pine plantations in southeastern Oklahoma. J. Wildl. Manage. 49:958-962. [ Links ]
Nelson, M. E. & L. D. Mech. 1981. Deer social organization and wolf predation in northeastern Minnesota. Wildl. Monogr. 77:1-53. [ Links ]
Nuñez, R. & B. Miller. 1997. Home range, activity and habitat use by jaguars and pumas in neotropical dry forest of Mexico. Proc. Seventh International Theriological Congress. Acapulco, México. [ Links ]
Núñez, R., B. Miller & F. Lindzey. 2000. Food habits of jaguars and pumas in Jalisco, México. J. Zool., Lond. 254:373-379. [ Links ]
Ordway, L. L. & P. R. Krausman. 1986. Habitat use by desert mule deer. J. Wildl. Manage. 50:677-683. [ Links ]
Owen-Smith, N. 1990. Demography of a large herbivore, the greater kudu Tragelaphus strepsiceros, in relation to rainfall. J. Anim. Ecol. 59:893-913. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Limusa, México, D. F. [ Links ]
Sánchez-Rojas, G., S. Gallina & S. Mandujano. 1997. Área de actividad y uso del hábitat de dos venados cola blanca (Odocoileus virginianus) en un bosque tropical caducifolio de la costa de Jalisco, México. Acta Zool. Mex. (n.s.) 72:39-54. [ Links ]
Short, H. L. 1963. Rumen fermentations and energy relationships in white-tailed deer. J. Wildl. Manage. 27:184-195. [ Links ]
Short, H. L., R. M. Blair & C. A. Segelquis. 1974. Fiber composition and forage digestibility by small ruminants. J. Wildl. Manage. 38:197-209. [ Links ]
Sih, A. 1993. Effects of ecological interactions on forager diets: competition, predation risk, parasitism and prey behaviour. Pp. 182-211. In: R. N. Hughes (Ed.). Diet Selection: an Interdisiciplinary Approach to Foraging Behaviour. Blackwell Scientific Publications, Oxford. [ Links ]
Silva-Villalobos, G., S. Mandujano, G. Arceo, S. Gallina & L. A. Pérez-Jiménez. 1999. Nutritional characteristcis of plants consumed by the white-tailed deer in a tropical forest of Mexico. Vida Silvestre Neotropical 8:38-42. [ Links ]
Sokal, R. R. & F. J. Rohlf. 1995. Biometry. W. H. Freeman & Co., San Francisco. CA. [ Links ]
Soto-Werchitz, A. 2000. Comportamiento de hembras de venado cola blanca (Odocoileus virginianus texanus) durante la época de crianza en un matorral xerófilo del noreste de México. Tesis de Maestría, Instituto de Ecología, A. C., Xalapa, Ver. [ Links ]
Thompson, C. B., J. B. Holter, H. H. Hayes, H. Silver & W. E. Urban, Jr. 1973. Nutrition of white-tailed deer. 1. Energy requirements of fawns. J. Wildl. Manage. 37:301-311. [ Links ]
Ullrey, D. E., W. G. Youatt, H. E. Johnson, L. D. Fay & B. L. Bradley. 1967. Protein requirement of white-tailed deer fawns. J. Wildl. Manage. 31:679-685. [ Links ]
Vangilder, L. D., O. Torgerson & W. R. Porath. 1982. Factors influencing diet selection by white-tailed deer. J. Wildl. Manage. 46:711-718. [ Links ]
Weckerly, F. W. 1994. Selective feeding by black-tailed deer: forage quality or abundance .J. Mamm. 75:905-913. [ Links ]

