










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkActa zoológica mexicana
On-line version ISSN 2448-8445Print version ISSN 0065-1737
Acta Zool. Mex vol.20 n.1 Xalapa Apr. 2004
Population parameters of Spodoptera frugiperda (Smith) (Lep.: Noctuidae) fed on corn and two predominant grasess in Tucuman (Argentina)
Gabriela Murúa & Eduardo Virla
PROIMI-Biotechnology (Biological Control Division) Av. Belgrano & Pje. Caseros (4000) S.M. de Tucumán-ARGENTINA. gmurua@yahoo.com.
Recibido: 12 de abril 2003
Aceptado: 7 de octubre 2003
ABSTRACT
A comparative laboratory study on population parameters of Spodoptera frugiperda (Smith) (Lep.: Noctuidae) fed on corn (Zea mays L.) and two of the most predominant grasses in Tucuman (Argentina) Guineagrass (Panicum maximum Jacq.) and Bermudagrass (Cynodon dactylon (L.) Pers.) was carried out. The diets used determined changes in diverse parameters such as duration of life cycle, number of larval instars, sex ratio, life expectancy, fertility and/or fecundity. Regarding to the life cycle duration, significant differences among host plants were recorded for the different developmental stages. Females fed on a corn diet showed a fertility of 92.1%, while those breeding on a diet based on Guineagrass showed 96.4% and 99.8% on Bermudagrass. Mean daily egg production ranged within 99.4 (fed on Bermudagrass) and 187.8 (fed on Guineagrass). Total mean egg production also varied with larval diet; maximum value was recorded for females fed on Guineagrass leaves (0 1282.7 ± 38.6 eggs/females). Results suggest that individuals bred on this grasses during final winter could play an important role in FAW population dynamic performance affecting corn crops in Argentina's Norther Region.
Key words: Spodoptera frugiperda, life history, host plant, performance, population parameters.
RESUMEN
Se realizó un estudio comparativo en condiciones de laboratorio sobre los parámetros poblacionales de S. frugiperda alimentada con maíz (Zea mays L.) y con las dos pasturas más predominantes en Tucumán (Argentina): pasto guinea (Panicum maximum Jacq.) y gramón (Cynodon dactylon (L.) Pers.). Las dietas usadas determinaron cambios en diversos parámetros tales como duración del ciclo de vida, número de instares larvales, proporción de sexos, expectativa de vida, fertilidad y fecundidad. Teniendo en cuenta la duración de ciclo de vida, diferencias significativas entre las plantas huésped fueron registradas entre los distintos estados de desarrollo. Las hembras mostraron una fertilidad del 92,1%, 96,4% y 99,8% cuando fueron alimentadas con maíz, pasto guinea y gramón, respectivamente. La producción diaria media de huevos varió entre 99,4 para hembras alimentadas con gramón y 187,8 para las alimentadas con pasto guinea. La producción media total de huevos también varió según la dieta suministrada, el máximo valor fue registrado para las hembras que durante el estadío larval fueron alimentadas con hojas de pasto guinea (0 1282.7 ± 38.6 huevos/hembras). Los resultados sugieren que los individuos alimentados con las pasturas antes mencionadas jugarían un papel importante en el comportamiento de la dinámica poblacional de S. frugiperda afectando cultivos de maíz en la región del Norese Argentino. Estas pasturas podrían albergar a la primera generación anual de la plaga y su manejo racional podría disminuir la incidencia del cogollero sobre los cultivos de maíz.
Palabras clave: Spodoptera frugiperda, ciclo de vida, planta huésped, comportamiento, parámetros poblacionales.
INTRODUCTION
Fall armyworm (FAW), Spodoptera frugiperda (Smith) (Lep., Noctuidae), is an important pest on various crops (corn, cotton, sorghum, and diverse pasture grasses) and it is widely distributed in America (Sparks 1979). By affecting corn crops in tropical and subtropical regions of Latin America, it becomes one of the most serious problems because of the importance of the damages that produces during the crop season.
In Northeastern Argentina, FAW is the most important corn pest determining losses that fluctuate between 17 to 72% (Perdiguero et al. 1967).
In Tucumán, Willink et al. (1993) studied yield losses in corn, and determined that early crops (early spring) are less affected. Commercial crops are growth in the region mainly at late spring and at the early summer (November-December), depending on the arrival of the first rains.
FAW larvae are frequently collected in the spontaneous grass communities during spring. The most abundant grasses are Guineagrass (Panicum maximum Jacq.) and Bermudagrass (Cynodon dactylon (L.) Pers.). In spite of this, the lack of field studies on life history of FAW in the region is evident, in particular when corn crops are not present. Due to its perennial pest status, strategies for management of FAW are required. To develop feasible strategies, it is often necessary to standardize one or more biological parameters of the FAW in a controlled environment. The knowledge of these characters is important, and literature shows that life and fecundity tables under controlled conditions provides useful information for characterizing population dynamic. FAW life cycle laboratory studies on these grasses could derive in formulation of strategies in order to manage the first generation responsible of colonization on corn crops.
The reproductive potential, behavior, fecundity, and fertility of the FAW have been studied under a variety of both natural and controlled environmental conditions. These published reports indicate a wide variation in those parameters, which may be influenced by temperature, larval diet and the strain of FAW (Simmons & Lynch 1990, Rogers & Marti 1994 b).
The aim of this contribution is to analyze the potential of Guinea and Bermuda grasses as possible hosts of FAW and to evaluate their influence in different biological parameters when these grass species are utilized as food resource for this pest.
MATERIALS AND METHODS
Collection of samples:
FAW individuals (larvae) used in this study were collected from corn crops in Vipos (26° 28' S, 65° 19' W) (Tucumán province). Rearing and testing of the insects were carried out between January 2000 and May of 2001 in chambers under controlled conditions at 25 ± 2 ºC, 70-75% RH and 14L: 10D of artificial photoperiod.
Rearing colony with corn and two grasses:
After establishing a continuous breeding colony, three cohorts (n: 130) were settled, feeding on either on corn, Guineagrass or Bermudagrass.
Adults of FAW were maintained in PET (polyethylene-terephthalate) cylindrical cages (30-cm high x 10-cm diameter). For aeration, the top was closed with a nylon mesh cloth, and a hole was made on one side. These cages contained pieces of paper that allowed the females to rest and to lay the eggs. Food was provided via a cotton plug saturated with honey and water mixture (1:1 vol/vol).
The cages were checked daily for oviposition and adult mortality; meanwhile egg masses were collected and deposited in glass tubes (12-cm high x 1.5-cm diameter). Once emerged, the neonate larvae were placed individually in glass tubes as mentioned above and fed with corn, Guineagrass and/or Bermudagrass leaves. The food was changed daily. Developing insects were observed through each instar and the data through ecdysis were recorded.
Reproduction data and Fecundity and Life horizontal tables:
To obtain Reproduction and Fecundity table 18 females were checked daily. Summary life tables for cohorts on each of the three host plants were built by combining the observational data on the three replicates (Rabinovich 1978, Sedlacek et al. 1986, Carey 1995, Bellows & Van Driesche 1999). The net reproductive rate "Ro", generation time "T", instantaneous rate of population increase "r", doubling time "D" and finite rate of increase "λ" were estimated from the summarized life tables using the following equations:
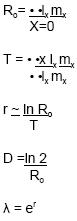
Interpretation of the data:
All data were analyzed by STATISTICS program (1995).
RESULTS AND DISCUSSION
Rearing colony with corn and two grasses (Table I)
Egg stage: Egg incubation period lasted 3.53 ± 1.17, 2.96 ± 0.89 and 3.08 ± 0.61 days, when the individuals are fed on corn, Guineagrass and Bermudagrass, respectively. Egg duration showed significant differences (P < 0.001, Tukey), the smallest duration was observed with Guineagrass.
This values are agreed with those mentioned by Doporto & Enkerlin (1964), Nieto-Hernández & Llanderal-Cázares (1982), Valverde et al. (1995) and Perez et al. (1997).
Larval stage: Significant differences were found in the duration of the larval stage. It is important to underline that the longest larval developmental time was recorded on those larvae fed on Bermudagrass (P < 0.001, Tukey). However, the larval stage duration in our tests was generally higher than the values reported by Doporto & Enkerlin (1964), Valverde et al.(1995), Pashley et al. (1995), Perez et al. (1997) and Pantoja et al. (1997).
FAW had a mean of seven, eight and nine larval instars when were fed with corn, Guineagrass and Bermudagrass respectively. It is important to mention that some larvae fed with corn went through up to eight and nine instars (3 and 1 respectively), and one individual fed with Guineagrass reached the tenth instar.
Pupal stage: Regarding to the duration of this stage, the means were 10.35 ± 1.41 when fed on corn, 10.8 ± 1.42 on Guineagrass and 11.38 ± 1.45 days on Bermudagrass. ANOVA analysis reveals that there are significant effects of the host plant in the duration of this stage, being observed the lesser value with larvae fed on corn (P < 0.001, Tukey). As it is represented in Table I, the pupal developmental time obtained in this study is in agreement with values reported by other authors (Doporto & Enkerlin 1964, Nieto-Hernández & Llanderal-Cázares 1982, Valverde et al. 1995, Pashley et al. 1995, Perez et al. 1997, Pantoja et al. 1997).
Reproduction data: (Table I)
Sex Ratio and Longevity: Sex ratio of S. frugiperda fed on corn resulted in a female-biased sex ratio (1.16:1), the opposite being true on Guineagrass (1:1.5) and non-biased in those fed on Bermudagrass (1:1) (Table II). In all treatments, females lived more than males. Analyzing adult longevity according to food, non significant differences (ANOVA, P < 0.001) were noticed. García & Clavijo (1989) using corn leaves, corn grains and artificial diet as foods made similar observations.
Adult longevity was 16 ± 2.8 days for the insects fed on Z. mays, 17.35 ± 5.39 days on P. maximum, and 16.23 ± 4.69 days on C. dactylon. As it is summarized in Table I, the observed longevities matched with those reported by Valverde et al. (1995) and Pantoja et al. (1997) working with diverse diets.
Considering the sex of the insects, significant differences were recorded only for individuals fed on P. maximum and C. dactylon (P < 0.05, t-test) where females (with an average of 21.1 and 18.27 days, respectively) lived more than males (14 and 14.2 days respectively).
It was also observed that longevity in unmated individuals was notably diminished in comparison with mated ones. When corn, Guineagrass and Bermudagrass are used as food resource, longevity of unmated was 7.02 ± 4.67, 9.91 ± 5.72 and 7.13 ± 3.9 days respectively. ANOVA analysis reveals that there are significant differences, recording the longest duration with Bermudagrass (P < 0.001, Tukey). This information contrasts with the one obtained by Rogers & Marti (1994a), where extending females' virginity increased their longevity.
Duration of life cycle: In laboratory and under the established ambient conditions, the life cycle (egg to adult) of S. frugiperda lasts a mean of 36.88 ± 8.95 days on maize, 36.14 ± 9.55 days on Guineagrass, and 36.29 ± 7.27 days on Bermudagrass. ANOVA analysis recorded a not significant effect of the host plant on the duration of the developmental cycle. The obtained duration are longer than the mentioned by Valverde et al. (1995), where for individuals fed on artificial diet, attained a life cycle of 28 days.
Fertility: When analyzing fertility data obtained among diets, significant differences were found. The lesser fertility was obtained when fed in Z. mais (P<0,001, Tukey). When fed on corn, females showed a fertility (egg viability) of 92.08%, those fed on Guineagrass 96.41% and until 99.84% on Bermudagrass. This observation contrasts with the one provided by García & Clavijo (1989) and Rogers & Marti (1994a).
Other biological parameters: (Table III) The longest preoviposition period was observed in those females whose larvae were fed with Bermudagrass; in spite of this the differences were not significant. The oviposition period was larger on individuals fed with Guineagrass (P < 0.001, Tukey).
Mean daily egg production by the females varied with the host plant provided from 99.4 in those breeding on Bermudagrass to 187.81 in those breeding on Guineagrass. ANOVA analysis showed significant effect of the diet on egg production, registering the smallest production with Bermudagrass (P < 0.001, Tukey). Simmons & Lynch (1990), working with FAW breeding on artificial diet recorded a mean of 284.1 eggs/female/day.
Mean total egg production also varied with to the larval food source, and significant differences were observed. The lowest value was recorded for females coming from larvae fed on Bermudagrass leaves (x: 621 ± 250 eggs/female) (P< 0.001, Tukey). Other authors (Simmons & Lynch 1990, Rogers & Marti 1994a, Sen-Seong et al. 1985, Pashley et al. 1995) reported higher values.
It is important to point out that after the pre-oviposition period, females of all the treatments did not lay eggs every day, and survived approximately two days after their last oviposition.
Analyzing the number of egg masses laid per female, significant differences were found. The highest number of egg masses laid by female was recorded in the individuals coming from larvae fed with Guineagrass ( 10.1 ± 2.74 egg masses/female) (P< 0.001, Tukey). It is important to highlight that the values obtained in this study were similar to the ones reported in other contributions.
10.1 ± 2.74 egg masses/female) (P< 0.001, Tukey). It is important to highlight that the values obtained in this study were similar to the ones reported in other contributions.
The differences in the number of eggs/egg mass in those individuals fed on the provided host plants was significant, being evidenced by highest number with Guineagrass (P < 0.001, Tukey). The mean number of eggs per egg mass was 108.74 ± 98.57, 147.65 ± 132.21 and 90.52 ± 59.14 eggs, when females were fed on Z. mays, P. maximum and C. dactylon respectively. These values differ from those obtained by Valverde et al. (1995) using artificial diet.
Fecundity Table: (Figs. 1, 2 and 3)


The calculated net reproductive rate "Ro" was 1316.67 (corn), 1214.85 (Guineagrass) and 678.78 (Bermudagrass); because this value was greater than one, under the laboratory conditions used, and in absence of natural enemies and/or others mortality factors, the population grew. Ro values here obtained were higher than the ones reported by Sen-Seong et al. (1985) using four corn genotypes as food resource.
When corn was used as host the generation time "T" was 50.80 days, with Guineagrass T= 46.38 days and with Bermudagrass T= 48.27 days. The instantaneous rate of population increase "r" (=intrinsic rate of increase) was 0.92, 0.96 and 0.78 females/females/day for Z. mays, P. maximum and C. dactylon respectively.
The doubling time "D" was 0.75, 0.72 and 0.88 days with corn, Guineagrass and Bermudagrass. The finite rate of increase "λ" was 2.51 (corn), 2.61 (Guineagrass) and 2.18 (Bermudagrass) days.
The number of female eggs per female per days (mx) on Guineagrass was higher than the one obtained from individuals reared on corn and Bermudagrass.
Life horizontal tables and survival analysis: (Table II) and (Figs. 4 and 5)


First larval instar showed a life expectancy of 16.82 days when fed on P. maximum, 13.71 days on C. dactylon and 18.75 days on Z. mays. The "ex" curves present inflections showing certain "increases" or "peaks" that point out which are the critical ages of the species regarding the risk to die at a determinate age.
In corn, the period of higher mortality occurred during the first three instars and during the passage from mature larvae to the pupal stage; 42.40% of the individuals reach the adult stage. With Guineagrass, the "ex" curve shows three periods of heavy mortality; the highest one occurred from second to third instar; the second period occurs from the fifth to sixth instar, and the third one occurs before adult emergency; only 42.45% of the individuals reached the adult stage. When Bermudagrass was used as the critical period of mortality occurred from the sixth to seventh instar.
Conclusions:
In laboratory studies, S. frugiperda successfully developed feeding on C. dactylon and P. maximum. The most important change was in mean total egg production. The highest number of egg laid by female was recorded in the individuals from larvae fed with Guineagrass.
Taking into account the importance of these grasses in northern Argentina, and that they could play an essential role in FAW population performance, it would be necessary to begin field studies in order to manage the first generation responsible to corn crops colonization and define FAW dispersal patterns and the factors governing them.
ACKNOWLEDGEMENTS
We appreciate the valuable comments of Dr. Carlos Coviella (Univ. California at Riverside) and Msc. Patricia Diez for commenting on this manuscript, the valuable statistical assistance of Dr. Gerardo Liljesthrom (CEPAVE –Univ La Plata). We are thankful to Mrs. G. Fanjul (PROIMI) and Dr. Carlos Coviella too for help on the English. This work was supported by CONICET PIP 0702/98, Argentina.
REFERENCES CITED
Bellows T.S. & R.G. Van Driesche. 1999. Life table construction and analysis for evaluating biological control agents. In: Bellows TS, Fischer TW (eds.), Handbook of Biological Control. Principles and Applications of Biological Control. Academics Press, 199-223. [ Links ]
Carey R.J. 1995. Insect demography. In: Encyclopedia of Environmental Biology (Vol. 2). Academic Press, 289-303. [ Links ]
Doporto L. & D. Enkerlein. 1964. La biología del cogollero Laphigma frugiperda (S y A) bajo condiciones de laboratorio. Folia Entomol. Mex., 7-8: 45-46. [ Links ]
Garcia J.L. & S. Clavijo. 1989. Efecto de la alimentación sobre la longevidad, fertilidad y fecundidad de Spodoptera frugiperda (Smith). Bol. Ent. Venez. N. S., 5 (6): 47-53. [ Links ]
Nieto-Hernadez R. & C. Llanderal-Cazares. 1982. Biología e identificación de estadíos larvales de Spodoptera frugiperda (Smith) (Lep. Noctuidae). Folia Entomol. Mex. 54: 11. [ Links ]
Pantoja A., C.M. Smith & J.F. Robinson. 1997. Development of the fall armyworm, Spodoptera frugiperda, strains from Louisiana and Puerto Rico. Envirom. Entomol. 16: 116-119. [ Links ]
Pashley D.P., T.N. Hardy & A.M. Hammond. 1995. Host effects on developmental and reproductive traits in the fall armyworm strains (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 88(6): 748-755. [ Links ]
Perdiguero J.S., J.M. Barral & M.V. de Stacul. 1967. Aspectos biológicos de plagas de maíz de la región chaqueña. Evaluación de daño. INTA, Est. Exp. Agrop., Presidencia Roque Saenz Peña, Boletín. 46: 1-30. [ Links ]
Perez B., F. Piedra & B. Blanco. 1997. Manejo integrado de Spodoptera frugiperda. Bol. Técnico N° 2, INISAV, Cuba. 8-10. [ Links ]
Rabinovich J.E. 1978. Ecología de poblaciones animales. Programa Regional Des. Cient. y Tec. de la OEA, Washington. Monografía N° 21: 114 pp. [ Links ]
Rogers C.E. & O.G. Marti Jr. 1994,a. Effects of age at first mating on the reproductive potential of the fall armyworm (Lepidoptera: Noctuidae). Population Ecology, 23(2): 322-325. [ Links ]
----------. 1994,b. Reproductive potential of once-mated of the fall armyworm (Lepidoptera: Noctuidae). Fla. Entomol. 77(4): 402-410. [ Links ]
Sedlacek J., K. Yeargan & P. Freytag. 1986. Laboratory life table studies on the blackfaced leafhopper (Homoptera: Cicadellidae) on Johnsongrass and Corn. Envirom. Entomol. 15: 1119-1123. [ Links ]
Sen-Seong N.G., F.M. Davis & E.P. Willians. 1985. Survival, growth, and reproduction of the fall armyworm (Lepidoptera: Noctuidae) as affected by resistant corn genotypes. J. Econ. Entomol. 78: 967-971. [ Links ]
Simmons A.M. & R.E. Lynch. 1990. Egg production and adult longevity of Spodoptera frugiperda, Helicoverpa zea (Lepidoptera: Noctuidae), and Elasmopalpus lignosellus (Lepidoptera: Pyralidae) on selected adult diets. Fla. Entomol. 73(4): 665-671. [ Links ]
Sparks A. 1979. A review of the biology of the fall armyworm. Fla. Entomol. 62(2): 82-87. [ Links ]
Valverde L., Z.A. de Toledo & S. Popich. 1995. Ciclo biológico de Spodoptera frugiperda (Smith) (Lep. Noctuidae). Acta Zool. Lilloana. 43(1): 131-143. [ Links ]
Willink E., M. Costilla & V. Osores. 1993. Daños, pérdidas y nivel de daño económico de Spodoptera frugiperda (Lep., Noctuidae) en maíz. Rev. Industrial Agric. Tucumán (EEAOC), 70(1-2): 49-52. [ Links ]

