










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkActa zoológica mexicana
versão On-line ISSN 2448-8445versão impressa ISSN 0065-1737
Acta Zool. Mex no.88 Xalapa Abr. 2003
Artículo
Fauna de coleoptera Melolonthidae de Villa Las Rosas, Chiapas, México
José Antonio Alcázar-Ruiz1, Alejandro Morón-Ríos2 y Miguel Angel Morón3
1,2 Departamento de Ecología y Sistemática Terrestre, El Colegio de la Frontera Sur (ECOSUR). Carr. Panamericana y Periférico Sur s/n, María Auxiliadora, CP 29290 San Cristóbal de las Casas, Chiapas, MÉXICO.
3 Departamento de Entomología, Instituto de Ecología, A. C. Apartado Postal 63, CP 91000 Xalapa, Veracruz, MÉXICO.
Recibido: 8 de febrero 2002
Aceptado: 29 de julio 2002
Resumen
Se presenta un estudio de las especies nocturnas de la familia Melolonthidae (sensu Endrödi, 1966) en Villa Las Rosas, Chiapas, con los objetivos de conocer su riqueza, abundancia y fenología. El municipio de Villa Las Rosas, se encuentra a 1,240 m de altitud, en un ambiente de transición entre selva baja subcaducifolia y bosque de coníferas. Para la captura de los ejemplares se utilizaron como trampas de luz las lámparas del alumbrado público. Durante 1999-2000 se aplicó un esfuerzo de colecta sistemática de 103 noches, y se capturaron 4,029 ejemplares pertenecientes a 50 especies de los géneros Diplotaxis, Phyllophaga, Polyphylla, Hoplia, Anomala, Pelidnota, Cyclocephala, Euetheola, Ligyrus, Coelosis, Enema, Strategus, Xyloryctes, Spodistes, Archophileurus, Hemiphileurus y Phileurus. La mayor riqueza y abundancia correspondió a las especies de Phyllophaga (16 spp.) y Anomala (11 spp.), algunas de las cuales podrían tener importancia económica, como posibles plagas que consumen las raíces de los cultivos en la región. Se citan por primera vez para México: Archophileurus simplex (Bates) y Phyllophaga anolaminata (Moser), y se consideran como primeros registros para Chiapas: Anomala decolor Bates, A. forreri Bates, Diplotaxis consentanea Bates, D. simillima Moser y Hoplia squamifera Burmeister. Se incluye una clave para separar las 50 especies encontradas. El 51% de las especies de Villa Las Rosas corresponde al patrón Paleoamericano con amplio éxito (sensu Halffter, 1976), pero a nivel genérico el predominio es Neotropical. Esta fauna tiene más similitud (QS Sorensen) con Los Tuxtlas, Veracruz (27.72%) y las montañas del Noreste de Hidalgo (27.40%).
Palabras clave: Melolonthidae, Phyllophaga, Anomala, taxonomía, distribución, selva baja subcaducifolia.
Abstract
A study of the nocturnal Melolonthidae beetle fauna of Villa Las Rosas, Chiapas, México is presented. Data were obtained during 1999-2000 with 103 nights of collecting effort at lights near the borders of subdeciduos tropical forest and pine-oak forest, located at 1,240 m of altitude. Records of 3 subfamilies, 9 tribes and 50 species of the genera Diplotaxis, Phyllophaga, Polyphylla, Hoplia, Anomala, Pelidnota, Cyclocephala, Euetheola, Ligyrus, Coelosis, Enema, Strategus, Xyloryctes, Spodistes, Archophileurus, Hemiphileurus and Phileurus are included. Higher specific diversity is presented in the genera Phyllophaga (16 spp.) and, Anomala (11 spp.). Phyllophaga tenuipilis (Bates), P. ravida (Blanchard), Xyloryctes lobicollis Bates and, Anomala inconstans Burmeister were the most abundant species in the samples. Archophileurus simplex (Bates) and Phyllophaga anolaminata (Moser) are cited for first time in México, and Anomala decolor Bates, A. forreri (Bates), Diplotaxis consentanea Bates, D. simillima Moser and, Hoplia squamifera Burmeister are recorded for first time from Chiapas state. A key to the 50 species is included. Most genera represented in the study area have Neotropical origin, but 51 % of the species belong to Paleoamerican dispersal pattern (sensu Halffter 1976). The Melolonthidae fauna of Villa Las Rosas area is 27.72% similar (QS Sorensen) with the species found in Los Tuxtlas, Veracruz; and 27.40% similar with the species from the mountains at northeast of Hidalgo, Mexico.
Key Words: Melolonthidae, Phyllophaga, Anomala, taxonomy, distribution, subdeciduous tropical forest.
Introducción
La familia Melolonthidae se encuentra distribuida ampliamente en el territorio mexicano, desde el nivel del mar hasta los 3,800 m de altitud, abarcando la mayor parte de los diferentes tipos de vegetación natural y modificados (Morón 1984). Morón et al. (1997) citan para México la presencia de 110 géneros y 1,040 especies. Entre los estados con mayor riqueza para esta familia se encuentra Chiapas con más de 250 especies (Alcázar 2000).
Las especies de esta familia desempeñan funciones importantes dentro de las comunidades bióticas a través de la polinización, la degradación, la facilitación del reciclaje de la materia orgánica, etc. (Morón 1984, Dirzo 1986; Palacios-Ríos et al. 1990). Sin embargo, esta familia también incluye especies cuyas larvas tienen importancia económica porque se alimentan con raíces de plantas vivas y dañan cultivos como el maíz, sorgo, trigo y caña de azúcar, entre otros (Morón 1983, Campos 1983).
En Chiapas la mayoría de los estudios entomológicos que se han realizado con esta familia se refieren a descripciones de nuevas especies, nuevos registros, o estudios faunísticos que, sin dejar de ser importantes aportan escasa información sobre su biología y ecología (Morón et al. 1985, Delgado 1990, Palacios-Ríos et al. 1990, Morón-Ríos & Morón 2001). En este sentido, diversos autores (Riess & Flores-Cáceres 1976, García-Martell 1977, Michel 1978, Morón 1983, 1986) han insistido en la importancia agrícola y ecológica que tienen las especies de Melolonthidae, sin embargo, aun es muy poco lo que se sabe acerca de la riqueza de este grupo en áreas consideradas de alta diversidad, y en éstas sólo se pueden efectuar estimaciones a partir de trabajos generales.
Considerando que los trabajos sobre la riqueza, abundancia e importancia de las especies de la familia Melolonthidae, realizados a la fecha en el estado de Chiapas se refieren a los ambientes tropicales cálido-húmedos o de bosque mesófilo de montaña, se planteó el presente estudio para conocer la diversidad, la abundancia, y la distribución estacional de este grupo de insectos en un ambiente semicálido-húmedo, como el que caracteriza a Villa Las Rosas, Chiapas, y compararlo con otras localidades mexicanas previamente estudiadas.
Área de estudio. La investigación se realizó en la cabecera municipal de Villa Las Rosas, Chiapas. El municipio se localiza en la parte central del Estado (Fig. 1), cuya cabecera municipal está situada dentro de las coordenadas 16° 21' 51" lat. N y 92° 21' 56" de long. O (INEGI, 1984) a 1,240 m de altitud. El municipio forma parte de la altiplanicie de Chiapas, la cual colinda con la Depresión Central. Su orografía está conformada por tres zonas con diferentes características de relieve: la zona accidentada se localiza al este y representa el 60% de la superficie, las zonas semiplanas se ubican en el centro y este del municipio (área de estudio) y la zona plana en el suroeste. El subsuelo está constituido por depósitos del Cretácico superior e inferior y del Oligoceno. Los tipos de suelo predominantes son el luvisol, gleysol y solonchak. Presenta varios cursos hidrológicos, como los ríos Yola y Seco y los arroyos Zanja de Piedra, Paso Hondo, Lamesao y Chila (Gobierno del Estado de Chiapas, 1988).

El municipio presenta un clima semicálido húmedo con lluvias en verano (Aw2(w)i) (García 1988). Los meses de abril a julio son los más cálidos y la época de lluvia es de mayo a octubre. La precipitación pluvial es de 1251.5 mm al año con una temperatura media anual de 19.8°C (Fig. 2) (CNA- Chiapas, 1980-1996). La vegetación característica es de selva baja subcaducifolia con transición a bosque de coníferas (Miranda 1975, Breedlove 1986). Los principales elementos arbóreos son: Cordia dodecandra, Byrsonima crasifolia, Calycophyllum sp., Combretum fruticosum, Leucaena sp., Bombax ellipticum, Juniperus comitana y J. gamboana, Pinus spp., Quercus scytophyla, Q. crispipilis, Crataegus pubescens, Salix humboldtiana, Taxodium sp., Acacia farnesiana, Acacia sp., Prosopis sp., (Gobierno del Estado de Chiapas, 1988). Debido al desarrollo de las actividades de agricultura y ganadería, también existen grandes zonas con pastizal inducido y cultivos de maíz y caña de azúcar. Destacan también los cultivos de frijol, café, plátano, guayaba y hortalizas. La vegetación original se conserva como remanentes en las faldas de los cerros y a lo largo de los cauces de arroyos de temporal, linderos y callejones.

Según Morón et al. (1997) se esperaría encontrar una gran diversidad de Melolonthidae en Villa Las Rosas, por presentar una vegetación característica de bosque tropical subcaducifolio, el cual constituye uno de los ecosistemas con mayor riqueza de Melolonthidae en el país.
Material y métodos
La captura se realizó dos días por semana durante 13 meses, de mayo 1999 a mayo de 2000. Los muestreos se realizaron durante cuatro horas comenzando al anochecer, para ello se utilizaron como trampas de luz las lámparas incandescentes del alumbrado público de 180 watts y un reflector de 300 watts, los cuales emiten dos tipos diferentes de longitud de onda (consideradas como luz blanca y amarilla respectivamente). Para realizar los muestreos se subdividió el poblado de Villa Las Rosas en cuatro zonas de acuerdo a los puntos cardinales, cada zona con tres sitios de muestreo fijo. La mitad de los sitios se ubican hacia el interior del poblado (6) y los restantes hacia las afueras. Para cada ocasión se colectaron ejemplares de una sola zona recorriendo los tres sitios aleatoriamente. En cada sitio se colectó durante una hora aproximadamente y se consideraron también los insectos encontrados durante el recorrido.
Los insectos capturados cada noche se sacrificaron en una cámara letal con acetato de etilo. Una muestra de dos a seis ejemplares por especie, sexo y fecha se montaron en alfileres entomológicos, en el caso de los machos se extrajeron sus órganos genitales para montarlos en seco y facilitar la determinación taxonómica; la otra parte del material se almacenó en sobres de papel glacine y posteriormente en alcohol al 70%. Los ejemplares se encuentran depositados en la Colección Entomológica de ECOSUR, San Cristóbal de Las Casas, Chiapas (ECOSC-E).
Para estimar la eficiencia de la colecta se construyó una curva de acumulación de especies para los Melolonthidae nocturnos. El arreglo de las especies se dio de acuerdo a su captura durante el periodo de estudio. Considerando las sugerencias de León-Cortés et al. (1998), se utilizó el "número de especímenes colectados por noche" como medida del esfuerzo de colecta. Con esta unidad de esfuerzo, la forma de la curva de acumulación llega a ser mas regular, además de que representa mejor el esfuerzo real, en comparación al uso de "noches" como medida de esfuerzo. En este ultimo caso, una noche con un solo espécimen capturado, aumenta las "noches" de esfuerzo en la misma cantidad como lo hace una noche con diez especímenes. El ajuste de los datos a los modelos de Clench y Von Bertalanffy se efectuó por medio del procedimiento de regresión no linear (SPSS v.10).
El modelo de Clench se describe como: S (t) = a* t / (1+ b* t ) y el de Von Bertalanffy: S (t) = a / b * [(1- exp (- b* t )]
Donde, S(t) es el tamaño esperado de la lista, a es la pendiente al comienzo de la colección, b es un parámetro relacionado a la forma de la acumulación de nuevas especies durante la colección, t es el esfuerzo de colecta, y a / b es la asíntota.
Resultados
Se capturaron un total de 4,029 ejemplares adultos en 103 noches de muestreo. Estos ejemplares pertenecen a 50 especies, de 17 géneros, nueve tribus y tres subfamilias. A continuación se indica la composición específica para cada género, y en el Apéndice I se presenta una clave dicotómica para su identificación.
MELOLONTHINAE, Melolonthini
Phyllophaga Harris. Se registraron 16 especies de los subgéneros Chlaenobia, Phytalus y Phyllophaga (s. str.) (Cuadro 1). La especie más abundante fue P. tenuipilis (Bates) con un registro de 1,028 individuos atraídos a las luces. Phyllophaga ravida (Blanchard) y P. testaceipennis (Blanchard) ocuparon los siguientes niveles de abundancia (871 y 280, respectivamente), mientras que Phyllophaga anolaminata (Moser), P. sp. aff. wittkugeli (Nonfried) y P. sp. aff. integra (Say) solo estuvieron representadas por uno a tres individuos. Phyllophaga anolaminata representa el primer registro para México, ya que sólo se le conocía para Honduras y Guatemala. Ocho especies, como P. (Chlaenobia) tumulosa (Bates) (Fig. 3) solo se colectaron durante el segundo período de colectas (marzo-mayo 2000), lo cuál podría indicar que tienen ciclo de vida bianual, o que su actividad de vuelo se adelantó durante 1999, por lo cuál no se les capturó en mayo de ese año.


Diplotaxis Kirby. Se capturaron 25 ejemplares, representados por seis especies (Cuadro 2). Las más abundantes fueron Diplotaxis commixta Vaurie y D. hirsuta Vaurie, las cuales se encontraron sobre el follaje de zacate silvestre, muy cerca de una trampa de luz. Es importante mencionar que en el cuadrante sur-poniente, donde se colectó bajo un reflector, se capturaron las demás especies: D. simplex Blanchard, D. sp aff. jacala Vaurie, D. consentanea Bates y D. simillima Moser, las últimas dos especies corresponden al primer registro para el estado de Chiapas.

Polyphylla Harris. Solo se capturó un macho de Polyphylla petiti Guérin poco después de las 21:00 hrs cerca de una trampa de luz. Anteriormente, esta especie se había citado para El Triunfo, Albino Corzo, Chiapas; Jalapa, Veracruz; Cerro del Higuerón y Jojutla, Morelos; Guadalajara, Jalisco (Morón et al. 1997, Morón-Ríos & Morón 2001, Navarrete et al. 2001).
MELOLONTHINAE, Hopliini
Hoplia Illiger. Por primera ocasión se cita Hoplia squamifera Burmeister para el estado de Chiapas, con base en una muestra de seis machos y tres hembras. Forma parte de un complejo de especies con amplia distribución desde el estado de Hidalgo en México, hasta Panamá (Blackwelder 1944, Morón et al. 1997).
RUTELINAE, Anomalini
Anomala Samouelle. Se capturaron 561 ejemplares que pertenecen a 8 especies (Cuadro 3). Anomala inconstans Burmeister es la especie más abundante con 402 ejemplares, se le capturó en el mes de mayo y se le observó consumiendo la parte interna de la cáscara de frutos de mango. También se encontraron sus larvas en suelos con cultivos de maíz y caña de azúcar. Se citan por primera ves para el estado de Chiapas Anomala decolor Bates y A. forreri Bates, que previamente solo se habían registrado para los estados de Veracruz y Sinaloa respectivamente (Morón et al. 1997).

RUTELINAE, Rutelini
Pelidnota MacLeay. Se capturaron 11 hembras y 6 machos de Pelidnota guatemalensis Bates, los ejemplares fueron capturados principalmente en el cuadrante nor-poniente. Esta especie tiene una distribución restringida a Chiapas (Comitán, Teopisca, Cañón del Sumidero), Belice y Guatemala. Sus larvas se desarrollan en troncos derribados o tocones en estado avanzado de descomposición (Hardy, 1975; Morón, obs. pers.).
DYNASTINAE, Cyclocephalini
Cyclocephala Latreille. Se capturaron 46 ejemplares representantes de cinco especies (Cuadro 4). Sólo se capturaron hembras de la especie más abundante Cyclocephala lunulata Burmeister, la cual se encuentra ampliamente distribuida en todo el país. C. amazona (Linné), C. complanata Burmeister, C. mafaffa Burmeister y C. stictica Burmesiter, se han registrado en bosques tropicales perennifolios y caducifolios y bosque mesófilo de montaña en nuestro país (Morón et al. 1997).

DYNASTINAE, Oryctini
Coelosis Hope. Dos machos y tres hembras de Coelosis biloba (Linné) fueron atraídos por luces de color amarillo en dos noches calurosas de octubre. Es una especie común aunque no es abundante, asociada con hormigas cortadoras del género Atta, distribuida entre el sureste de México y Argentina (Morón et al. 1997, Navarrete 2001).
Enema Hope. Se capturaron 31 individuos de Enema endymion Chevrolat en los cuadrantes sur- poniente y sur-oriente. Se distribuye desde el sureste de México hasta Bolivia. En algunas localidades de Chiapas puede ser muy abundante, y ha sido registrada en Ocozocuautla, Tuxtla Gutiérrez, Chiapa de Corzo y Berriozábal (Morón et al. 1997). En Villa Las Rosas se encontraron sus larvas dentro del suelo, frecuentemente parasitadas por el hongo ascomiceto Cordyceps melolonthae var. rickii (Lloyd) Mains (Guzmán et al. 2001).
Strategus Hope. Se recolectaron 46 ejemplares de Strategus aloeus (Linné). Se trata de una especie muy común distribuida desde el sur de los Estados Unidos hasta Argentina. Sus larvas se desarrollan en suelos ricos en materia orgánica, y en troncos derribados en estado avanzado de descomposición (Morón et al. 1997).
Xyloryctes Hope. Se capturaron 550 ejemplares de Xyloryctes lobicollis Bates. Entre el 14 y el 30 de abril de 2000 esta especie fue muy abundante en todo el poblado, en promedio hubo más de 50 ejemplares por lámpara. Se le ha registrado de varias localidades de Oaxaca y Chiapas, así como en casi toda América Central (Morón et al. 1997). Sus larvas se desarrollan en suelos de origen forestal o en troncos muy descompuestos (Morelli & Morón, 2002).
DYNASTINAE, Pentodontini
Ligyrus Burmeister. Se capturaron 212 ejemplares de Ligyrus (Ligyrodes) sallei Bates y 23 de L. (Ligyrus) nasutus Burmeister. La primera de ellas tiene amplia distribución en nuestro país, y también se le ha citado del sur de los Estados Unidos, y en América Central, sobre todo en tierras altas con clima templado; sus larvas se desarrollan en suelos ricos en materia orgánica. Ligyrus nasutus se ha registrado desde Texas hasta Costa Rica, en regiones bajas con clima cálido, pero se desconoce su biología (Morón et al. 1997).
Euetheola Bates. Este género estuvo representado por 34 individuos de Euetheola bidentata Burmeister, especie que en México habita distintos ambientes tropicales y subtropicales de la vertiente del Pacífico, y también se ha registrado en Venezuela y Brasil (Blackwelder 1944, Morón et al. 1997). Sus larvas se desarrollan en el suelo, y los adultos se han citado en Villaflores, Chiapas como dañinos para las plántulas de maíz, confundidos como Euetheola humilis Burmeister (Pontigo & Badii 1988).
DYNASTINAE, Agaocephalini
Spodistes Hope. Sólo se capturó una hembra de Spodistes monzoni Warner en un potrero al comenzar la noche. Es una especie poco común, localizada en bosques caducifolios y subperennifolios de Guerrero (Mochitlán), Oaxaca (Pochutla), Chiapas (Huixtla) y Guatemala (Morón et al. 1997).
DYNASTINAE, Phileurini
Phileurus Latreille. Se capturó una hembra de Phileurus didymus (Linné) cerca de un potrero al ser atraída a la luz. Las larvas se desarrollan dentro de troncos derribados y tocones en estado avanzado de descomposición, y los adultos pueden depredar a otros coleópteros xilófilos. Es una especie con amplia distribución desde el sur de los Estados Unidos hasta Argentina; en Chiapas se ha registrado para Tuxtla Chico, Tapachula, Palenque, Ocozocuautla, Yaxchilan y Boca del Chajul (Morón et al., 1997).
Hemiphileurus Kolbe. Se capturó un macho de Hemiphileurus dejeani (Bates) (Fig. 4) y una hembra de Hemiphileurus microps (Burmeister). La primera se ha citado para el sureste de México y en Guatemala; en Chiapas se tienen registros de ella en Bochil, Ocosingo y Berriozábal. Hemiphileurus microps se conoce de Hidalgo, Puebla, Veracruz, Oaxaca y Chiapas (Ocosingo). Las larvas se desarrollan dentro de troncos podridos y los adultos permanecen la mayor parte del tiempo en ese ambiente (Morón et al. 1997).

Archophileurus Kolbe. Por primera ocasión se cita a Archophileurus simplex (Bates) para México, con base en una hembra colectada en el cuadrante nor-poniente. Los miembros de este género pertenecen a un linaje sudamericano, supuestamente muy antiguo, con buena capacidad para colonizar medios hostiles, xéricos, muy diferentes de los bosques tropicales en donde pudieron haberse originado (Morón 1990). Hasta ahora esta especie era conocida únicamente en Nicaragua y Costa Rica. Su hallazgo en la localidad bajo estudio origina una amplia discontinuidad en su distribución e implica la posibilidad de que también exista en el resto de Centroamérica.
Discusión
Fenología y ecología
Respecto a su abundancia, cuatro especies (Phyllophaga tenuipilis, P. ravida, Xyloryctes lobicollis y Anomala inconstans) representan el 71% de los ejemplares con 2,851 individuos. De las especies restantes, 15 están representadas por un solo ejemplar. La mayor riqueza y abundancia está concentrada en un solo género ( Phyllophaga ), que reúne a la tercera parte de las especies (30.61%) y el 61.17% de los individuos de nuestras muestras. La riqueza del género Phyllophaga también ha sido observada en otros bosques tropicales caducifolios y subcaducifolios del país (Deloya et al. 1993).
Al agrupar las especies de acuerdo con sus hábitos alimentarios (Morón 1984, Morón & Deloya 1991), el 81.63% de los individuos capturados corresponde a especies filo-rizófagas o saprófagas facultativas, cuyas larvas se desarrollan dentro del suelo, y sólo el 18.36% de estos ejemplares representan especies saprófagas estrictas o saproxilófagas que frecuentan distintos microhábitats. La presencia de un gran número de especies con adultos filófagos y larvas rizófagas de los géneros Phyllophaga, Anomala, Diplotaxis y Cyclocephala, puede interpretarse como el producto de una colonización más o menos reciente, ya que son especies euriecas muy adaptables, que se han estado insertando en un proceso de transición, durante el cuál una comunidad típica de un bosque subcaducifolio está siendo reemplazada por una comunidad propia de un matorral xerófilo.
Al reunir los registros de actividad mensual de todas las especies recolectadas, se observa que la mayor riqueza (r) y abundancia (a) anual de especies se concentra en los meses de abril (r =28 ; a = 31.1%) y mayo (r = 33; a = 45%) en relación directa con la temporada de lluvias, observándose también un descenso en los meses más secos y fríos (Fig. 5). Estas fluctuaciones están relacionadas con los ciclos vitales de las especies, lo cual propicia la concentración de adultos en la época más favorable para la reproducción y el desarrollo inicial de sus larvas, cuyos recursos alimentarios tienen una distribución espacio-temporal más o menos homogénea y constante (raíces, humus y madera podrida). Este patrón se ha observado en otros lugares como en La Michilía, Durango, Chamela, Jalisco y El Tentzo, Puebla (Morón & Deloya 1991, Morón 1996, Morón et al. 1990, 2000). Sin embargo, en otras localidades tropicales húmedas como Los Tuxtlas, Veracruz y Boca del Chajul, Chiapas, la riqueza y abundancia de adultos son notables aún durante los meses menos favorables (Morón 1979, Morón et al. 1985).

Es posible distinguir tres patrones fenológicos que involucran tanto a las especies filo-rizófagas, como a las saprófagas. El primer patrón agrupa a las especies poco abundantes, las cuales aparecen con las primeras lluvias de marzo y abril, hasta desaparecer entre junio y julio. Este patrón lo presentaron 28 especies: Anomala decolor, A. denticollis, A. discoidalis, A. donovani, A. sp. aff. trapezifera, Cyclocephala amazona, C. complanata, C. mafaffa, C. stictica, Diplotaxis commixta, D. consentanea, D. simillima, D. simplex, D. sp. aff . jacala, Enema endymion, Hoplia squamifera, Phileurus didymus, Phyllophaga anolaminata, P. cometes, P. lenis, P. parvisetis, P. soctona, P. sp. aff . integra, P. sp. aff. temora, P. sp. aff. wittkugeli, P. tumulosa, Polyphylla petiti y Spodistes monzoni. En el segundo patrón, las especies también aparecen con las primeras lluvias de abril cuando alcanzan su máxima representatividad, pero su abundancia decrece entre junio y julio hasta desaparecer en agosto. Este patrón se observó en Anomala forreri, A. inconstans, A. sticticoptera, Diplotaxis hirsuta, Pelidnota guatemalensis, Phyllophaga menetriesi, P. multipora, P. obsoleta, P. tenuipilis, P. testaceipennis, P. ravida y Xyloryctes lobicollis.
En el tercer patrón, las especies se distinguen porque aún cuando algunas inician su actividad con las primeras lluvias, o en junio, alcanzan su máxima abundancia en julio, decrecen en agosto y disminuyen o desaparecen en octubre, como ocurre con Ligyrus nasutus, Strategus aloeus y Euetheola bidentata.
Es posible observar también que algunas especies parecen estar menos vinculadas con la temporada de lluvias, como Diplotaxis hirsuta, Archophileurus simplex, Coelosis biloba, Hemiphileurus dejeani, y H. microps que aparecen antes o después de las lluvias, y por otra parte Cyclocephala lunulata y Ligyrus sallei que tienen actividad a lo largo de todo el año.
Por lo que respecta a la riqueza y abundancia total por cuadrante del poblado, el mayor número de especies rizófagas se registró en la zona sur, mientras que el número de saprófagas fue similar en todos los cuadrantes (Cuadro 5). La riqueza de especies rizófagas de la zona sur se debe quizás a los cultivos de caña de azúcar que existen en este lugar, y a su cercanía con la montaña en la zona oriente. Por otro lado, el 57% de la abundancia se registró en la zona poniente, que es una planicie donde abundan los cultivos (maíz, frijol, etc.), además de que en el cuadrante sur-poniente la fuente de luz fué un reflector (300 watts) que pudo haber ejercido mayor atracción e incrementado las capturas.
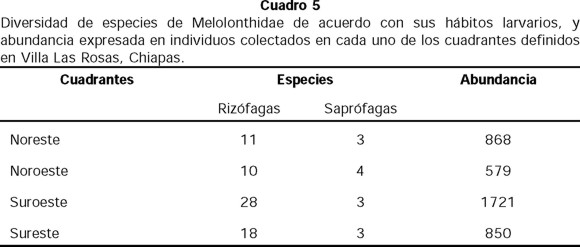
En este estudio no se separaron las especies colectadas por tipo de luz, ni por muestra (interior y exterior del poblado). Sin embargo, se observó que las especies se colectaron tanto bajo focos de luz blanca como amarilla, y que su abundancia fue mayor en el primer tipo de luz. También se observó que la abundancia tiende a ser mayor en los focos que se encuentran más alejados del poblado. En este sentido sería importante realizar muestreos en otras localidades cercanas al municipio dentro del siguiente gradiente altitudinal: montañas al oriente del poblado a 1800 msnm y en las zonas bajas al poniente a 700 msnm.
Respecto a la proporción de sexos encontrados, en general se capturó una mayor proporción de hembras (63%) que de machos. Sin embargo las especies que tuvieron una abundancia mayor a 10 individuos, Xyloryctes lobicollis, Phyllophaga lenis, P. cometes, P. tumulosa, P. soctona, Ligyrus nasutus y Anomala sticticoptera, presentaron una mayor proporción de machos. La proporción 1:1 únicamente fue observada para Phyllophaga obsoleta y Enema endymion.
Curva de acumulación de especies por esfuerzo de colecta
Las predicciones basadas en los modelos de Clench y Von Bertalanffy coinciden con el número de especies observadas para el esfuerzo de colecta efectuado (Fig. 6). El modelo de Clench estima 60 especies al aumentar en 100 noches el esfuerzo de colecta, mientras que el modelo de Von Bertalanffy predice 54 especies para el mismo incremento en el esfuerzo de captura. A partir de estos datos se considera que el modelo de Clench predice mejor la acumulación de especies nuevas para esta localidad. Como se menciona mas adelante, el tamaño de la fauna de otros sitios con condiciones ambientales similares es cercano a 60 especies. Por lo anterior es posible que la fauna de Melolonthidae nocturnos de Villa las Rosas esté prácticamente completa y probablemente falten por registrar entre seis y diez especies, probablemente de los géneros Phyllophaga, Diplotaxis y Anomala.

El 84% de las especies capturadas se obtuvieron en 18 noches de colecta, las cuales coinciden con el inicio y la mitad del periodo de lluvias. Doce de las especies capturadas en estos períodos, están representadas por un solo ejemplar. A este tipo de comportamiento de las especies Rabinowitz et al. (1986) lo clasifican como rareza demográfica. Se trata de especies que presentan densidades bajas en el área de estudio aunque éstas tengan amplia distribución, tal como sucede con Cyclocephala amazona, Diplotaxis consentanea, D. jacala, D. simillima, Polyphylla petiti, Phyllophaga temora, P. tumulosa y Hoplia squamifera.
Según Halffter & Ezcurra (1992) y Deloya et al. (1993, 1995) la abundancia de las capturas refleja relativamente bien a las poblaciones reales conforme el tamaño de muestra aumenta; por ello las faunas mejor estudiadas presentan un exceso de especies raras. En este sentido es posible pensar que con el número de especies raras capturadas (12) y con la intensidad de muestreo realizado en Villa Las Rosas (103 noches) se hayan recolectado la mayoría de las especies nocturnas de esta localidad, como lo señala la estimación del modelo de Clench antes mencionada.
Es importante mencionar que es mínimo el número de especies nocturnas que rara vez o nunca son atraídas a la luz; Morón et al. (1997) citan para México nueve especies en esa situación. Por otro lado, en algunas especies únicamente un sexo es atraído por la luz, sin embargo, esto no afectaría la riqueza total encontrada. Dentro de las limitaciones encontradas con el uso de las trampas de luz se puede considerar que éstas no proporcionan datos precisos sobre los hábitos de alimentación y apareamiento, aunque permiten conocer las fechas y horarios de vuelo de las especies. Las trampas de luz, como otros tipos de muestreo, tienen radios de acción desconocidos.
Respecto a las horas de vuelo de las especies nocturnas de esta familia, la mayoría inicia su actividad en el crepúsculo, concentrando su mayor actividad en las dos primeras horas de la noche. En general son escasos las referencias de especies que estén volando después de las 00:00 horas.
Por último, al analizar el número total de especies de la familia Melolonthidae en varias localidades del país, se observa que las especies nocturnas aportan en promedio el 85% de la riqueza, mientras que la proporción de especies diurnas es del 15%, tal como se observa en el Cuadro 6. Aunque es necesario hacer más eficientes los métodos de colecta de especies diurnas. A partir de los datos anteriores es posible estimar que en Villa Las Rosas faltarían por conocer nueve especies diurnas, probablemente de los géneros Macrodactylus, Anomala, Strigoderma, Epectinaspis, Cotinis, Euphoria y Trigonopeltastes.

Representación faunística y zoogeográfica
De acuerdo con los patrones de dispersión propuestos para la Zona de Transición Mexicana (Halffter 1976, 1978, 1987), las 17 especies de Pelidnota, Cyclocephala, Euetheola, Ligyrus, Strategus, Phileurus, Archophileurus, Hemiphileurus y Spodistes quedan incluidas dentro del patrón Neotropical típico, con cuatro especies propias de la Zona de Transición Mexicana ( Hemiphileurus dejeani, H. microps, Spodistes monzoni y Pelidnota guatemalensis ). El patrón Paleoamericano lo presentan 26 especies de los géneros Phyllophaga, Polyphylla, Anomala y Hoplia, las tres primeros con amplio éxito. Los géneros Diplotaxis y Xyloryctes, que agrupan a 7 especies, pertenecen al patrón de dispersión en el Altiplano Mexicano.
A nivel genérico se observa un claro predominio de los elementos neotropicales, pero a nivel específico, el predominio corresponde a elementos paleoamericanos (Cuadro 7) debido a la gran diversidad de especies de Phyllophaga y Anomala. Al parecer, la mayoría de los géneros paleárticos y algunas especies sudamericanas se han introducido aprovechando la deforestación en toda la zona, y su transformación a pastizales, monocultivos y acahuales, como sucede con las especies de Phyllophaga, Anomala y algunas de Cyclocephala (Morón 1979). La mezcla de los tres patrones faunísticos en Villa Las Rosas sería atribuible a la proximidad de las montañas y a la cercanía con la Depresión Central de Chiapas, ubicación que resulta estratégica, porque combina áreas templadas y subtropicales que aportan numerosos elementos neotropicales a la fauna típica de los 1500 msnm. En este sentido, la localidad estudiada y la Depresión Central tienen gran importancia sobre todo como un corredor de baja altitud que ha permitido el paso de fauna neotropical y paleoamericana hacia ambos lados del continente, lo que apoya las ideas expuestas por Halffter (1976, 1978, 1987) sobre la composición de la entomofauna de la Zona de Transición Mexicana y los desplazamientos que han originado su actual estructura.

Indice de Similitud
Al comparar la fauna de Coleoptera Melolonthidae nocturnos obtenida en este trabajo con las de otras zonas estudiadas en el país, mediante el índice de similitud QS de Sorensen (1948), Los Tuxtlas, Veracruz (Morón 1979) y las montañas del norte de Hidalgo (Morón 1994) son las que ofrecen una mayor similitud a nivel específico aunque el porcentaje es bajo (Cuadro 8). La combinación de las faunas (Halffter 1976, 1978, 1987, Halffter & Ezcurra 1992) resulta interesante, por un lado se presentan elementos de origen sudamericano con reciente penetración (Los Tuxtlas, Veracruz) y por otra parte especies con origen Paleoamericano y especies con filiación del Altiplano Mexicano (montañas de Hidalgo).
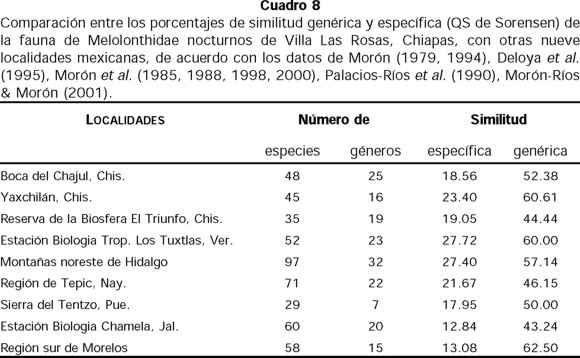
En contraste, la mayor similitud a nivel de género se presenta con la fauna del sur de Morelos, esto se explica al comparar sus características climáticas, altitudinales y de vegetación, que son las más similares al área de estudio. Sin embargo, a nivel de especie la similitud fue la menor, debido al gran predominio de elementos paleoamericanos.
Al analizar las faunas de Melolonthidae nocturnos en el país (Cuadro 8), se observa la gran riqueza que presenta Villa Las Rosas para un solo año de colectas sistemáticas, en comparación con varios años de capturas más esporádicas o heterogéneas, en otras localidades como Los Tuxtlas, Veracruz, Chamela, Jalisco y el Sur de Morelos. Esta riqueza concuerda con lo que se ha registrado para los bosques caducifolios y subcaducifolios parecidos al área de estudio (Morón et al. 1997).
Agradecimientos
El presente trabajo es la contribución No. 3 de la línea de investigación: Dinámica de Poblaciones y Comunidades de Insectos del Departamento de Ecología y Sistemática Terrestre de ECOSUR. Se reconoce el apoyo y colaboración de Liliana Avendaño Vera, José A. Espinoza y Obdulia Alcázar. Manuel Girón Intzin nos apoyó con la preparación y montaje del material, Jorge León Cortés gentilmente nos asesoró en el ajuste de los modelos de acumulación de especies por esfuerzo de colecta e hizo valiosas sugerencias a las versiones preliminares del manuscrito, al igual que Lorena Ruiz Montoya.
Literatura citada
Alcázar-Ruiz, J. A. 2000. La fauna de Coleoptera Melolonthidae de Villa Las Rosas, Chiapas. Tesis profesional (no publicada). Escuela de Biología, Universidad de Ciencias y Artes de Chiapas. 58 pp. [ Links ]
Blackwelder, R. E. 1944. Checklist of the coleopterous insects of México, Central America, the West Indies and South America, Pt. 2. Bull. U. S. Nat. Mus., 185: 189-341. [ Links ]
Breedlove, D. 1986. Listados florísticos de México. IV Flora de Chiapas. UNAM. Instituto de Biología. México. 246 pp. [ Links ]
Campos, B. R. 1983. Las gallinas ciegas como plagas del suelo. pp. C15-C38. In: Memoria II Mesa Redonda sobre plagas del suelo. Chapingo, México. [ Links ]
CNA-Chiapas, 1980-1996. Datos de precipitación pluvial mensual y temperatura media correspondiente al período 1980 a 1996, estación Villa Las Rosas, Municipio Villa Las Rosas, Chiapas. Gerencia Regional Frontera Sur, Subgerencia Regional Técnica, Jefatura de Proyecto del Servicio Meteorológico, Tuxtla Gutiérrez, Chiapas. [ Links ]
Delgado, C. L. 1990. Dos nuevas especies mexicanas de Diplotaxis del grupo "puberea" (Coleoptera: Melolonthidae; Melolonthinae). Folia Entomol. Mex. 78: 61-70. [ Links ]
Deloya, C., A. Burgos, J. Blackaller & J. M. Lobo. 1993. Los coleópteros Lamelicornios de Cuernavaca, Morelos, México (Passalidae, Trogidae, Scarabaeidae y Melolonthidae). Boletín Soc. Ver. Zool. 3(1): 15-55. [ Links ]
Deloya, C., M. A. Morón & J. M. Lobo. 1995. Coleoptera Lamellicornia (Macleay, 1819) del sur del estado de Morelos, México. Acta Zool. Mex. (n.s.) 65:1-42. [ Links ]
Dirzo, R. 1986. Insectos y plantas. SEP. México. 78 pp. [ Links ]
Endrödi, S. 1966. Monographie der Dynastinae (Coleoptera: Lamellicornia) I.Teil. Entomologische Abhandlungen Museum Tierkunde, Dresden 33: 1-457. [ Links ]
García, E. 1988. Modificaciones al Sistema de Clasificación Climática de Köppen. 5ª Edición. Talleres Offset Larios S.A. Editado por la autora, México. 120 pp. [ Links ]
García-Martell, C. 1977. Lista de insectos y ácaros perjudiciales a los cultivos de México. Fitófilo 73. México. 175 pp. [ Links ]
Gobierno del Estado de Chiapas. 1988. Las Rosas. Memorias municipales. Tuxtla Gutiérrez, Chiapas. Gobierno Constitucional del Estado de Chiapas, Tuxtla Gutiérrez. 34 pp. [ Links ]
Guzmán, G., M. A. Morón, F. Ramírez-Guillén & J. H. D. Wolf. 2001. Entomogenous Cordyceps and related genera from Mexico with discussions on their hosts and new records. Mycotaxon 78: 115-125. [ Links ]
Halffter, G. 1976. Distribución de los insectos en la Zona de Transición Mexicana. Relaciones con la entomofauna de Norteamérica. Folia Entomol. Mex. 35: 1-64. [ Links ]
----------. 1978. Un nuevo patrón de dispersión en la Zona de Transición Mexicana: el Mesoamericano de Montaña. Folia Entomol. Mex. 39-40: 219-222. [ Links ]
----------. 1987. Biogeography of the montane entomofauna of México and Central America. Ann. Rev. Entomol. 32: 95-114. [ Links ]
Halffter, G. & E. Ezcurra. 1992. La diversidad Biológica de Iberoamérica. Acta Zool. Mex. Programa Iberoamericano de Ciencia y Tecnología para el desarrollo. Inst. de Ecol. A. C., Ver. México. 389 pp. [ Links ]
Hardy, A. R. 1975. A revision of the genus Pelidnota of America North of Panama (Col. Scarab. Rutelinae). Univ. Calif. Publ. Entom. 78: 1-43. [ Links ]
INEGI. 1984. Carta topográfica 1:50 000. Venustiano Carranza, Chiapas. Clave: E15D72. [ Links ]
León-Cortés, J. J., Soberón-Mainero & J. Llorente-Bousquets. 1998. Assessing completeness of Mexican sphinx moth inventories through species accumulation functions. Diversity and Distributions 4: 37-44. [ Links ]
Michel, J. B. 1978. Importancia económica de las plagas del suelo en el estado de Jalisco. pp. 53-59. In: Memoria mesa redonda plagas del suelo. Sociedad Mex. Ent., A.C. México. [ Links ]
Miranda, F. 1975. La vegetación de Chiapas. 1a parte. 2a Edición. Ediciones del Gobierno del Estado. Tuxtla Gutiérrez, Chiapas. 325 pp. [ Links ]
Morelli, E. & M. A. Morón. 2002. Description of Xyloryctes Hope larvae with key to species based on the third instar larvae (Coleoptera: Scarabaeidae: Dynastinae). Coleopt. Bull. 56 (3): (en prensa) [ Links ]
Morón, M. A. 1979. Fauna de Coleópteros lamelicornios de la Estación de Biología Tropical "Los Tuxtlas", Veracruz, UNAM, México. An. Inst. Biol. UNAM (ser. Zool.) 50 (1):375-454. [ Links ]
----------. 1983. Introducción a la biosistemática y ecología de los Coléopteros Melolonthidae, edafícolas de México. pp. C1-C14. In: Memoria de la II Mesa Redonda sobre plagas del suelo Chapingo México. Sociedad. Mex. Ent., A.C. México. [ Links ]
----------. 1984. Escarabajos. 200 millones de años de evolución. Instituto de Ecología. Pub. 14. México. 132 pp. [ Links ]
----------. 1986. El género Phyllophaga en México. Morfología, distribución y sistemática supraespecífica. (Insecta:Coleoptera). Publ. No. 20. Inst. de Ecol., A.C. México. D.F. 341 pp. [ Links ]
----------. 1990. Descripción de una especie nueva de Archophileurus Kolbe, 1910 (Coleoptera: Melolonthidae, Dynastinae). An. Inst. Biol. UNAM (Ser. Zool.), 61 (1): 139-146. [ Links ]
----------.. 1994. Fauna de Coleoptera Lamellicornia en las montañas del Noreste de Hidalgo, México. Acta Zool. Mex. (n.s.) 63:7-59. [ Links ]
----------. 1996. Melolonthidae (Coleoptera). pp. 287-307. In: Llorente-Bousquets, J., A. García-Aldrete y E. González-Soriano (Eds.). Biodiversidad, taxonomía y biogeografía de artrópodos de México: Hacia una síntesis de su conocimiento. UNAM. México. [ Links ]
Morón, M. A., A. Aragón, A. M. Tapia-Rojas & R. Rojas-García. 2000. Coleoptera Lamellicornia de la Sierra del Tentzo, Puebla, México. Acta Zool. Mex. (n.s.) 79: 77-102. [ Links ]
Morón, M. A. & C. Deloya. 1991. Los Coleoptera Lamellicornia de la Reserva de la Biosfera La Michilía, Durango, México. Folia Entomol. Mex. 81:209-283. [ Links ]
Morón, M. A., C. Deloya & L. Delgado-Castillo. 1990 (1988). Fauna de coleópteros Melolonthidae, Scarabaeidae y Trogidae de la región de Chamela, Jalisco, México. Folia Entomol. Mex. 77: 313-378. [ Links ]
Morón, M. A., C. Deloya, A. Ramírez-Campos & S. Hernández-Rodríguez. 1998. Fauna de Coleoptera Lamellicornia de la región de Tepic, Nayarit, México. Acta Zool. Mex. (n.s.) 75:73-116. [ Links ]
Morón, M. A., Ratcliffe B. C. & C. Deloya. 1997. Atlas de los escarabajos de México. Coleoptera: Lamellicornia. Vol.I Familia Melolonthidae. CONABIO y Sociedad Mexicana de Entomología, A.C. México. 280 pp. [ Links ]
Morón, M. A., F. J. Villalobos & C. Deloya. 1985. Fauna de coleópteros Lamellicornios de Boca del Chajul, Chiapas, México. Folia Entomol. Mex. 66: 57-118. [ Links ]
Morón-Ríos, A & M. A. Morón. 2001. La fauna de Coleoptera Melolonthidae de la Reserva de la Biosfera "El Triunfo" Chiapas, México. Acta Zool. Mex. (n.s.) 84: 1-25. [ Links ]
Navarrete-Heredia, J. L. 2001. Beetles associated with Atta and Acromymex ants (Hymenoptera: Formicidae: Attini). Trans. Amer. Ent. Soc., 127 (3): 381- 429. [ Links ]
Navarrete-Heredia, J. L., L. Delgado & H. Fierros. 2001. Coleoptera Scarabaeoidea de Jalisco, México. Dugesiana 8 (1): 37-93. [ Links ]
Palacios-Ríos, M., V. Rico-Gray y E. Fuentes. 1990. Inventario preliminar de los Coleoptera Lamellicornia de la zona de Yaxchilán, Chiapas, México. Folia Entomol. Mex. 78: 49-60. [ Links ]
Pontigo, M. A. & M. H. Badii. 1988. Dispersión y fluctuación poblacional de Euetheola humilis (Coleoptera: Scarabaeidae) en maiz de temporal en Villaflores, Chiapas. pp.115-133. In: Memoria Tercera Mesa Redonda de Plagas del Suelo. Sociedad Mexicana de Entomología, A.C. e ICI de México. [ Links ]
Rabinowitz, D., S. Cairns & T. Dillon. 1986. Seven kinds of rarity. pp. 182-204. In: Silé, M. E. (ed.). Conservation biology. Sinauer, Sunderland, Mass. [ Links ]
Riess, C. & S. Flores-Cáceres. 1976. Catálogo de plagas y enfermedades de la caña de azúcar en México. Libro 11, Serie Divulgación Técnica, IMPA. México. 177 pp. [ Links ]
Sorensen, T. 1948. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content. Vidensk. Selsk. Biol. Skr. 5(4): 1-34. [ Links ]
SPSS, 1999. Release 10.05 Standard Version SPSS, Inc. USA. [ Links ]

