










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkActa zoológica mexicana
On-line version ISSN 2448-8445Print version ISSN 0065-1737
Acta Zool. Mex n.86 Xalapa Aug. 2002
Nota científica
Notes on the nest habits and host range of cleptobiotic Lestrimelitta niitkib (Ayala 1999) (Hymenoptera: Meliponini) from the Yucatan Peninsula, Mexico
José Javier G. Quezada-Euán y J. A. González-Acereto
Facultad de Medicina Veterinaria, Universidad Autónoma de Yucatán. Apdo. Postal 4-116, Mérida, Yucatán CP 97100 MEXICO. Email: qeuan@tunku.uady.mx
Abstract
Lestrimelitta niitkib is the only cleptobiotic stingless bee species reported for the Yucatán Peninsula. Very little is known about its biology and host range although it is recognised that its role may be crucial in the ecology of stingless bee populations. In this note we present preliminary information on the nest characteristics of this species, its host range and behaviour during nest raids.
Lestrimelitta is a Neotropical genus among the highly social stingless bees (Apidae: Meliponini) that obtains food and nesting materials through exclusively raiding nests of other social bees. Their life-style, so called cleptobiosis, is together with the African genus Cleptotrigona, unique to these bees among all eusocial Apidae (Roubik, D.W. 1989. The Ecology and natural history of Tropical bees. Cambridge, Univ. Press, Cambridge). Workers of this genus show a highly modified morphology addapted to robbing. They have lost all but few branched hairs on the body and the corbiculae that are used in Apidae to collect pollen and resins from plants.
In Yucatan, there are 13 documented species of native Meliponines, only one species of Lestrimelitta is so far been reported (Ayala, R. 1999 Folia Entomol. Mex. 186:1-123). Ten years ago this species was classified as L. limao, but recently it has been assigned a separate taxonomic status.This nova specie seems to be indigenous to the Yucatan Peninsula and Chiapas and has been classified as L. niitkib (Ayala op. cit.). Little information has been gathered about the limits of its geographic distribution and thus no information is available about interactions with other species. We have made some observations on the nesting characteristics and the interactions of L. niitkib with other meliponids and Apis mellifera colonies. We report on those observations in this note.
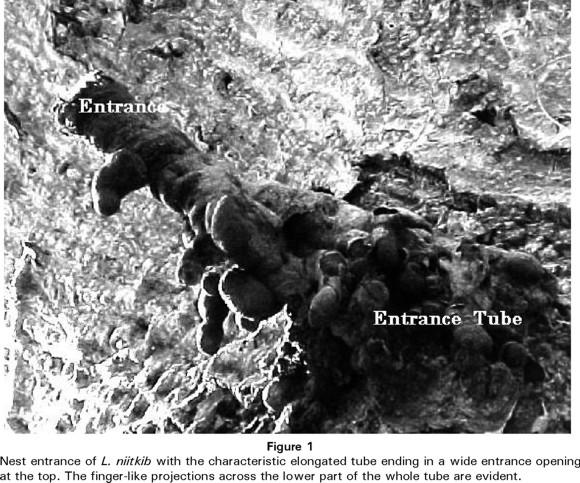
Nest characteristics of L. niitkib. The nest structures of L. niitkib were measured in 6 colonies collected in several localities across the state of Yucatán. Compared with other Meliponids from Yucatán, the nests of L. niitkib are among the largest, the cavities were in average 10.51 (± 3.47). The entrance tube to the nest is in a characteristic and rather conspicuous shape which gives the species its Mayan name "niit-kib" meaning nose of wax (Fig. 1). The tube is made of cerumen (a mixture of bees wax and plant resins) it is in average 10 cm long and 6 cm wide with thick finger-like prolongations in its lower part and the entrance always faces upwards. The height above the ground of the entrance tube in the nests we observed was between 2.7 to 3.2 m. As many as 40 guardian workers were observed at the entrance of the nests. The volume occupied by the brood was in average 3.71 with as many as 37 brood combs (mean=22). Worker brood cells were 0.5 cm by 0.3 cm; the queen cells were larger and measured 0.65 cm by 0.45 cm and located in the periphery of the combs. Interestingly, the combs in some nests were horizontal and helicoidal in others. Storage pots were 2.3 cm by 1.7 cm. There were no separate pollen and honey pots. A mixture of pollen and honey was stored in the pots. This mixture has a very pleasant lemon flavour. The nests we have found, were in cavities of the tree Bursera simaruba (n=4) and cavities on cement and stone walls (n=2). We have the impression that colony density of L. niitkib is rather low compared with other stingless bee species, three nests we found were separated by approximately 2km from each other. This is confirmed by the fact that nests seem difficult to find by beekeepers in rural areas. This species shows a remarkable adaptability to the scarcity of natural nesting sites due to deforestation compared with other Yucatecan species such as Melipona, they use a wide range of cavities. Additionally, three nests we collected easily survived and developed in hive-boxes which also indicates its adaptability.
Host species and nest raiding. We have witnessed 4 complete nest raids to colonies of Meliponids plus observations that indicated a nest raid was already on its way or ending (n=12). L. niitkib seems to have a marked preference for nests of Trigona (Frieseomelitta) nigra and Nannotrigona perilampoides. In contrast with L. limao in Brazil, which does not seem to have T. (F.) nigra as a host, the latter species is the preferred one for L. niitkib (Sakagami et al.1993. Sociobiology 21: 237-277). Out of 16 raids we have witnessed 9 were to nests of T. (F.) nigra, 4 to N. perilampoides, 1 to M.. beecheii, 1 to A. mellifera and 1 to L. niitkib. The raids to the latter three species destroyed and killed those colonies. Intraspecific aggression is strong, and colonies of L. niitkib kept in the same bee yard usually do not last long, only one of them survives. This may serve as a mechanism of control of population density within the species as a result of competition for food. However, raids to T.(F.) nigra and N. perilampoides only in two ocassions killed the colonies, in one of these, the process led to a foundation of a new L. niitkib colony. The rate of raids to nest of stingless bees and A. mellifera that we have witnessed during 3 years is reported in table 1.

We have also included some characteristics of the honey and population of the host species. Apparently, L. niitkib prefers attacking the species with more concentrated honeys and that are less defensive. However, the raids to A. mellifera which is a defensive species may be explained by the fact that they are not native to the neotropics and do not have adaptive strategies to defend against L. niitkib. We have no information on the influence of the quantity and quality of pollen reserves in the host to trigger L. niitkib raids, but one must expect that mechanisms have evolved for the robber bees to judge the quality of the food reserves in their hosts. Raids started with a mass of L. niitkib workers arriving to the host colony, this usually caused N. perilampoides and T. (F.) nigra guard workers to withdraw inside their colony and arriving foragers to stop going in. The attacker bees kept their mandibles open and fanning was intense. After 3-5 minutes at the entrance of the nest, workers of the robber bees started entering the host colony. In colonies of N. perilampoides there occurred a modification of the entrance tube, it was enlarged in length and width to resemble that of L. niitkib but smaller. It has been suggested that the tube might act as a land mark for parasite bees to guide them to the host colony (Sakagami, S.F. et al. 1993. Sociobiology 21: 237-277). Guards of L. niitkib positioned themselves at the entrance tube and guarded it in what seemed like a barrier against the entrance of host foragers coming back from the field since these were not allowed to enter. A strong lemon smell was evident at the entrance of the host colony. After 20-30 minutes of the mass arrival, workers of L. niitkib were witnessed coming out in mass and flying off the host colony carrying food and resins, workers of the host colony started entering their colony again with minimal interaction with the robbers. One raid was recorded to last 5 hours. Colonies of T. nigra do not build an entrance tube, when raiding this species, no construction of a tube by L. niitkib occurs, although 1-3 workers of the latter were seen at the small entrance of the host colony in guarding attitude. Workers of N. perilampoides and T. (F.) nigra do not seem to pose much opposition against raids, few workers were killed especially in the latter. In contrast, workers of M. beecheii, A. mellifera and L. niitkib strongly defend their colonies and many workers were killed. Interestingly, colonies of Scaptotrigona pectoralis, S. mexicana, Partamona bilineata, T. fuscipennis and Plebeia sp. that were in the same locality were never seen being raided. In particular, S. pectoralis and S. mexicana display a strong defensive behaviour towards L. niitkib. Only the slight smell of citral (the pheromone of Lestrimelitta) in the vicinity of their colonies triggers a mass of workers to come out of their nest and search for the origin of the odour. This is in strong contrast with the fact that in Panama S. pectoralis is the preferred host for L. limao (Sakagami et al. 1993. Sociobiology 21: 237-277).
It has been observed that when colonies of L. niitkib were kept within a few meters of Scaptotrigona colonies, the latter constantly attacked them and in one situation actually killed one colony. This behaviour was not triggered by food robbing since the reserves of L. niitkib were left intact. Thus, when a few colonies of Scaptotrigona are kept in a meliponid bee yard, attacks of L. niitkib to succeptible species have been reduced to zero (González-Acereto & Medina-Medina 1998.In: VI Congreso Ibero-Americano de Apicultura, Mérida.México).
The swarming process. We observed that one of the raids to a nest of N. perilampoides ended in the establishment of a new nest of L.nitkib. The first part of the process started with a typical mass raid followed by a fight between workers of the two colonies and the nest entrance of N. perilampoides being modified and ending with L. niitkib guarding the entrance. In this case, all the food pots of the host colony were left intact, but the brood combs were destroyed. Adult and newly emerged N. perilampoides workers were dismembered and discarded outside their colony. Later, larvae and pupae were also removed together with their cocoons. They were left as a pile of debris below the nest entrance. This process took about 5 days and lasted until the brood chamber was left empty and all the host bees were eliminated. After these 5 days, a L. niitkiib virgin queen moved into the nest. Three days after, the construction of new combs and oviposition had started already. Two days later, the construction of new food pots started and the nest entrance tube was completed with all the dimensions and charactersitic shape of L. niitkib. This process of swarming of L. niitkib contrasts with other stingless bee species because apparently there is no strong connection via food and building materials between the mother and daughter colonies (Michener, C. D. 1974. The social behavior of the bees.Harvard, Univ, Press, Cambridge). The food and building materials are obtained mainly from the host nest. This mechanism may have evolved as a way to avoid competition for food resources between mother and daughter colonies.
Acknowledgements
We are thankful to CONACyT 27891-N and PRODUCE which supported our studies with stingless bees. We also thank Ricardo Ayala and an anonymous reviewer for their suggestions.

