










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkActa zoológica mexicana
versão On-line ISSN 2448-8445versão impressa ISSN 0065-1737
Acta Zool. Mex no.86 Xalapa Ago. 2002
Artículo
Dieta y nicho de alimentación del coyote, zorra gris, mapache y jaguarundi en un bosque tropical caducifolio de la costa sur del estado de Jalisco, México
Sergio Guerrero*, Mohammad H. Badii**, Silvia S. Zalapa* y Adriana E. Flores**.
*Centro de Estudios en Zoología, Universidad de Guadalajara. Apdo. Postal 1-1919, CP 44101, Guadalajara, Jalisco, MÉXICO.
**Departamento de Entomología, Facultad de Ciencias Biológicas, UANL. Apdo. Postal 391, CP 66450, San Nicolas de los Garza, Nuevo León, MÉXICO.
Recibido: 2 de julio 2001
Aceptado: 11 de febrero 2002
Resumen
Se analizó la dieta, amplitud y sobreposición de nicho de alimentación para el coyote (Canis latrans), zorra gris (Urocyon cinereaoargenteus), mapache (Procyon lotor) y jaguarundi (Herpailurus yagouaroundi) en una zona de bosque tropical caducifolio de la costa sur de Jalisco, México. Para ello se recolectaron 430 excrementos de las cuatro especies: coyote (118), zorra gris (78), mapache (198) jaguarundi (36), fueron analizados y los contenidos determinados. Los resultados son reportados en frecuencia relativa de ocurrencia (FRO). Para el análisis de amplitud de nicho se aplicó el índice de Levins y el de Renkonen para determinar sobreposición de nicho. Los resultados mostraron que el material vegetal fue el elemento de mayor FRO para coyote (36.76%), zorra gris (38.16%) y mapache (47.62%), mientras que los mamíferos lo fueron para jaguarundi (43.37%). Otros grupos como insectos, aves, crustáceos y peces complementan la dieta de estas especies en diferentes proporciones; cambios en la frecuencia de esos grupos en cada especie son reportados entre la estación húmeda y seca. El valor con mayor amplitud de nicho lo presentó la zorra gris (15.32) y el coyote (14.95) y el menor el mapache (11.16) y el jaguarundi (4.46), aunque estos valores se ven modificados al analizar los datos por estación del año (seca-húmeda). Respecto a la sobreposición del nicho de alimentación, el mayor fue para coyote-zorra gris (57.18%) y el menor fue para mapache y jaguarundi (19.95%). Estacionalmente, el coyote y el jaguarundi registraron el mayor valor en la sobreposición de nicho, tanto en la estación seca (58.26%) como en la húmeda (49.48%).
Palabras clave: Amplitud de nicho, carnívoros, dieta, sobreposición de nicho.
Abstract
Food habits, niche breadth and niche overlap were analyzed for coyote (Canis latrans), gray fox (Urocyon cinereoargenteus), raccoon (Procyon lotor) and jaguarundi (Herpailurus yagouaroundi) in an area of dry forest located southern coast of the State of Jalisco, Mexico. We analyzed a total of 430 scats of the four species: coyote (118), gray fox (78), raccoon (198), and jaguarundi cat (36). The results are reported in relative frequency of occurrence (FRO). For the analysis of niche breadth, it was used the index of Levins, while the index of Renkonen was used in order to determined the niche overlap. The results showed that the vegetable food items was highest FRO for coyote (36.76%), gray fox (38.16%) and raccoon (47.62%). In the other hand, the mammals food items had the FRO highest for jaguarundi (43.37%). Other preys like insects, birds, crustaceans and fish supplemented the diet of these species in different proportions. Changes in the frequency of the food items utilized in the wet and dry seasons are reported per species. The niche was broad for the gray fox (15.32) and the coyote (14.95), and narrow for the raccoon (11.16) and the jaguarundi cat (4.46). However these results changed when the season of the year (dry-wet) was considered. The largest niche overlap was among coyote and gray fox (57.18%) and the smallest among raccoon and jaguarundi (19.95%). The coyote and jaguarundi cat showed the highest value of niche overlap in both dry season (58.26%) and wet season (49.48%).
Key Words: Carnivores, Food habits, niche breadth, niche overlap.
Introducción
La dieta es uno de los aspectos ampliamente estudiados para los carnívoros en Norteamérica. Entre las especies en las que se ha puesto mayor énfasis están el coyote (Ozoga & Harger 1966, Gipson 1974, Litvaitis & Shaw 1980, MacCraken & Uresk 1984, Andelt 1985, Andelt et al. 1987), zorra (Errington 1935, Wood 1959, Fritzell & Haroldon 1982, Garrott et al. 1983, Kodany 1995), mapache (Dearborne 1932, Stuewer 1943, Cagle 1949, Hamilton 1951, Schoonover & Marshall 1951, Tester 1953, Wood 1954, Gander 1966) y algunos felinos (Fitts & Sealander 1978, Ackerman et al. 1984, Delibes & Hiraldo 1987). El enfoque principal de muchos de estos trabajos ha sido conocer el efecto de la depredación de estas especies sobre otras de interés para el hombre (venado, ganado, etc.), o bien su efecto sobre cultivos (maíz, trigo, entre otros). En los últimos años ese enfoque ha cambiado, lo que ha permitido demostrar su relevancia no sólo para profundizar en el conocimiento de la ecología de las especies sino, además, para establecer planes y programas de conservación, manejo y aprovechamiento de las mismas (Korschgen 1980).
El concepto de nicho ha sido incorporado en diversos estudios de dieta (Badii et al. 1992). Hutchinson (1957) lo definió como el rango total de las variables ambientales (p. e. alimento, temperatura, ph, etc.) a las cuales una especie está adaptada y en las cuales vive y se substituye a si misma. Esta concepción del nicho permitió darle un enfoque cuantitativo, cuya base está en dos descriptores: amplitud y sobreposición (Smith 1982). La amplitud es la suma total de la variedad de recursos utilizados por una especie, en cambio la sobreposición, resulta de las demandas simultaneas sobre algunos recursos por parte de dos o más especies (Giller 1984). El alimento es una de las más relevantes dimensiones del nicho (Krebs 1999), ya que su análisis muestra, por un lado, la gama de elementos usados en la dieta de una especie, y por el otro, el grado de interacción entre especies con base en su dieta, particularmente en las que comparten un gremio trófico.
En México, son pocos los estudios que describen la dieta para varios carnívoros y la mayoría de los realizados se han enfocado en el coyote (Vela-Coiffer 1985, Servin & Huxley 1991, García-Hernández 1994 y Aranda et al. 1995), además de concentrarse en ambientes de bosque templado y zonas áridas. Esto es más evidente en Jalisco por el bajo números de investigaciónes (Graff 1988, Esparza-García 1991).
El objeto del presente estudio fue describir la dieta del coyote, zorra gris, mapache y jaguarundi en una zona de bosque tropical caducifolio de la costa del estado de Jalisco, y analizar la amplitud y sobreposición del nicho de alimentación de dichas especies. Esto aportará elementos para entender su coexistencia en este tipo de hábitats. Cabe hacer notar que las especies mencionadas están entre los carnívoros más comunes en la zona de trabajo (Ceballos y Miranda 1986, López-Acosta & Guerrero 1994, Ordorica-Hermosillo 1996).
Materiales y métodos
El área de estudio es conocida como Tenacatita y es una llanura costera con una extensión aproximada de 4,000 ha, localizada en el Municipio de La Huerta, estado de Jalisco. Topográficamente está limitada al Este por el parteaguas de los cerros y lomeríos de la llanura de La Manzanilla y la Bahía Tenacatita; al Oeste con el parteaguas de los cerros y lomeríos de la llanura costera El Tecuán y la albufera La Fortuna; al Norte por los lomeríos de la zona de Miguel Hidalgo y el poblado Aguacaliente y al Sur por el Océano Pacífico. Sus coordenadas geográficas son 19°16' y 19°21' de latitud N y 104°51' y 104°55' de longitud W (López-Acosta & Guerrero 1994).
Los tipos de vegetación presentes en la zona de estudio son bosque tropical caducifolio, manglares, bosque tropical subcaducifolio, pastizales y vegetación halófila. Los cultivos son comunes, particularmente los cocoteros, platanares y papayas, entre los de tipo permanente, y maíz entre los de temporal (Ceballos & Miranda 1986, López-Acosta & Guerrero 1994).
El trabajo de campo se llevó a cabo durante un año, de agosto de 1994 a julio de 1995. Se recolectaron 430 excrementos de coyote (n=118), zorra (n=78), mapache (n=198) y jaguarundi (n=36), utilizando para su identificación los criterios de Aranda (1981) Halfpenny & Biesot (1986) y Servín & Huxley (1991), tales como presencia de huellas de la especie, tamaño, forma y olor de la excreta.
Las muestras fueron secadas a temperatura ambiente y sometidas a un proceso de lavado en una solución a base de agua y detergente hasta remover todo el material soluble. Los materiales resultantes se separaron y colocaron en bolsa de plástico. La determinación de los contenidos se llevó al nivel taxonómico más específico posible, de acuerdo con las condiciones del material encontrado en las excretas. Para mamíferos se usaron características del pelo (escama y médula), mediante el método propuesto por Arita & Aranda (1987). El material óseo (principalmente dientes) se comparó con ejemplares recolectados en la zona. Los insectos, crustáceos, peces, aves y material vegetal, fueron determinados con el apoyo de investigadores del Centro de Estudios en Zoología, del Instituto de Botánica y del Departamento de Ecología, de la Universidad de Guadalajara.
Los resultados son reportados como frecuencia relativa de ocurrencia (FRO = (fi/∑fi)X 100) (Servín & Huxley 1991; Aranda et al. 1995), esto es, la frecuencia de cada elemento (fi), con relación a la frecuencia total de elementos registrados (∑fi). Se consideraron como elementos traza aquellos que presentaron menos de 1% de FRO, por lo que no fueron incluídos en los resultados.
Para conocer la amplitud del nicho de alimentación de cada una de las especies, se utilizó el índice de Levins (Krebs 1999), cuya fórmula es la siguiente: B=(1/∑Pi2), en donde Pi= ni/N, siendo ni= la frecuencia de ocurrencia del elemento i, y N= la frecuencia total de elementos. Para conocer y comparar la sobreposición o empalme del nicho de alimentación entre dos o más especies, se utilizó el índice de Renkonen ((Pjk=[∑(minimum Pij,Pik)] 100), en donde Pjk=porcentaje de sobreposición entre la especie j y la especie k, Pij=proporción del recurso i utilizado por la especie j, Pik=proporción del recurso i utilizado por la especie k, n=número total de categorías de recursos (Magurran 1989, Krebs 1999). Para esto se consideraron todos los elementos registrados en los excrementos, incluyendo aquellos con valores menores al 1% de FRO.
Resultados
Dieta del Coyote
Un total de 118 heces fecales de coyote fueron recolectadas. Donde el material vegetal conformó el 36.76%, los mamíferos el 33.82%, las aves el 12.5% y los insectos con un 11.39%, siendo estos los grupos con mayor FRO en todo el periodo de estudio. Estacionalmente se observó que los mamíferos fueron más frecuentes durante la estación húmeda (36.41%), que en la seca (28.41%). Un patrón estacional similar se observó para el material vegetal (38.04% y 34.09% respectivamente). En cambio la FRO de aves fue inversa a lo antes descrito, ya que en la estación seca fue mayor su consumo (18.18%) respecto a la húmeda (9.78%). Con relación a los insectos, se obtuvieron valores similares entre ambas estaciones (11.36% la seca y 11.41% la húmeda). Para reptiles su FRO fue superior en la estación seca (6.82%) que en la húmeda (2.72%) (Cuadro 1A).
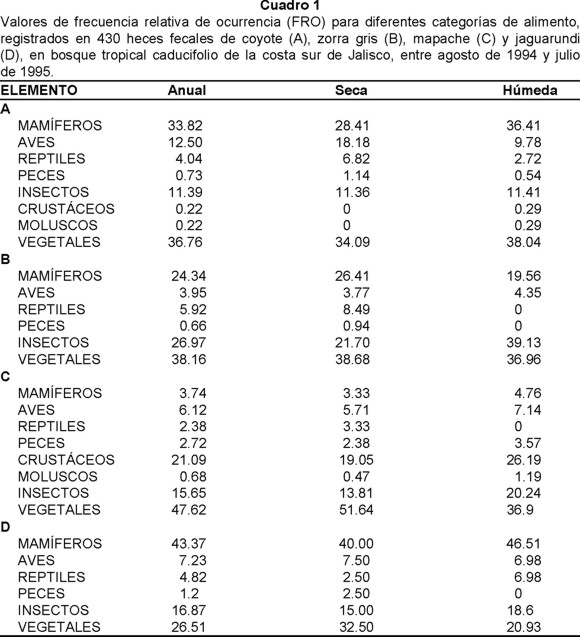
Entre los vegetales, se registraron como las más frecuentes a las gramíneas (Gramineae) (17.83%), compuestas (Compositae) (3.7%), coco (Cocus nucifera) (3.04%), ciruela (Spondias sp.) (2.61%), cactáceas (Cactaceae) (2.39%), solanáceas (Solanum sp.) (2.17%) y sandía (Citullus vulgaris) (1.96%). Aunque las gramíneas fueron dominantes en ambas estaciones, se observó una mayor frecuencia de consumo en la época húmeda (18.37%) que en la seca (16.24%). Otros frutos, como el coco y la ciruela, mostraron su mayor consumo durante la estación seca. En cambio las compuestas y la sandía (Citullus vulgaris) fueron más consumidas en la estación húmeda. La papaya (Carica papaya), cactáceas, guamúchil (Phitecellobium dulce) y otras leguminosas fueron consumidas únicamente en la estación húmeda (Cuadro 2).

Los mamíferos con mayor consumo fueron la rata algodonera (Sigmodon mascotensis) (9.78%), la rata arrocera (Oryzomys couesi) (3.91%), el ganado (Bos taurus) (4.56%) y el ratón espinoso (Liomys pictus) (1.96%). En ambas estaciones (seca y húmeda) tanto la rata algodonera como la carroña de ganado y la rata arrocera, fueron las más consumidas. Entre las aves, los mímidos (Mimidae) (1.09%) tuvieron los consumos más frecuentes tanto anual como estacionalmente. Cabe hacer notar que la mayor proporción de huesos y plumas de aves encontrados en las heces fecales no pudieron ser determinados (Cuadro 2).
La familia de insectos de mayor FRO fueron los chapulines (Acrididae) (3.04%); seguidos de escarabajos (Scarabaeidae) (1.52%), mayates (Melolonthidae) (1.52%), hormigas (Formicidae) (1.96%) y gorgojos o picudos (Curculionidae) (1.09%). Estas mismas presas predominaron en ambas estaciones del año, a excepción de gorgojos que no fueron registrados durante la época húmeda (Cuadro 2).
Reptiles, peces, crustáceos y moluscos formaron parte también de la dieta del coyote. Del primer grupo la iguana negra (Ctenosaura pectinata) fue la especie más consumida (1.96%), siendo su frecuencia de aparición mayor en la estación húmeda que en la seca (Cuadro 2).
Dieta de la Zorra gris
Un total de 78 heces fecales de zorra fueron recolectadas, donde se encontraron las siguientes categorías de alimentos: los vegetales (38.16%), los insectos (26.97%) y los mamíferos (24.34%) que fueron los más frecuentes. En la estación húmeda los insectos fueron los más consumidos (39.13%), seguido de vegetales (36.96%) y mamíferos (19.56%). Sin embargo, en la estación seca estos últimos incrementaron sus FRO (26.41%), en tanto se mantuvo el consumo del material vegetal (38.68%), mientras que los insectos disminuyeron (21.70%) (Cuadro 1B).
Las gramíneas (13.78%), el fruto de la guázima (Guazuma ulmifolia) (2.76%) y el fruto del guamúchil (2.76%) fueron los materiales vegetales más frecuentemente consumidos a lo largo del año. Las gramíneas fueron consumidas abundantemente en ambas estaciones, alcanzando el 14.44% en la estación húmeda y 13.41% en la seca. En esta última, el guamúchil (4.27%) y la guázima (4.27%) también registraron una frecuencia de consumo considerable (Cuadro 3).
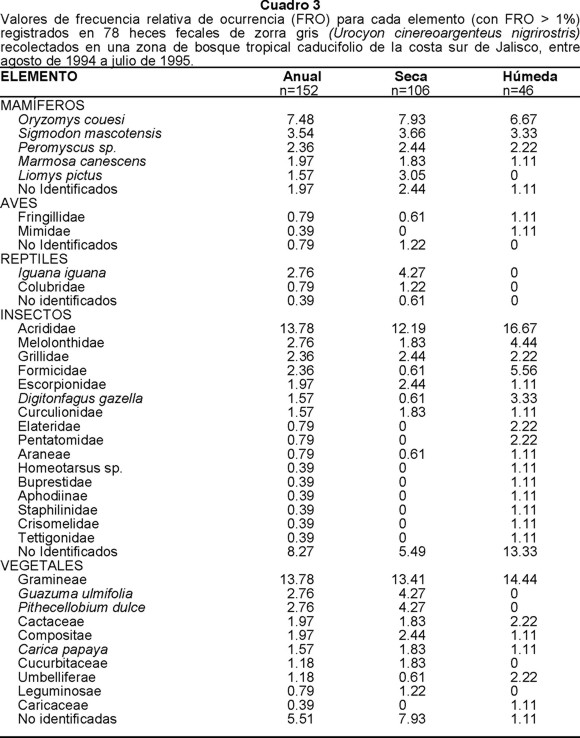
Para la zorra gris, los insectos fueron uno de los grupos más importantes en su dieta. Los chapulines fueron los de mayor FRO (13.78%). Se registraron también mayates (Scarabaeidae), afodinos (Aphodinidae) y grillos. En la estación húmeda la variedad de insectos fue alta, predominando los chapulines (16.67%), cuya frecuencia fue mayor con relación a la estación seca. En la misma estación seca, los gorgojos, escorpiones (Escorpionidae) y grillos tuvieron mayor frecuencia que en la húmeda; no así los mayates cuyas frecuencias se mantuvieron similares en ambas estaciones (Cuadro 3).
De los mamíferos, los de mayor FRO fueron la rata arrocera (7.48%), la rata algodonera (3.54%) y ratones del género Peromyscus sp. (2.36%). Estacionalmente no se presentó cambio en el patrón antes señalado; aunque para la estación seca se registró el consumo del ratón tlacuache (Marmosa canescens), no así en la húmeda (Cuadro 3).
La dieta de esta especie incluyó además reptiles, aves y peces. Los reptiles se presentaron solamente en las heces fecales recolectadas en la estación seca, principalmente iguana verde (Iguana iguana) (2.76%). Las proporciones en aves fueron más altas en la estación húmeda en donde los gorriones (Fringillidae) aparecieron como el principal componente (0.79%) (Cuadro 3).
Dieta del Mapache
Para el mapache, 198 heces fecales fueron analizadas. El material vegetal (47.62%), los crustáceos (21.09%) y los insectos (15.65%) constituyeron los componentes con mayor FRO. Las aves (6.12%), los mamíferos (3.74%), los peces (2.72%) y los reptiles (2.38%) presentaron las FRO más bajas. En la estación seca el material vegetal registró el 51.64%, y en la húmeda disminuyó hasta un 36.90%. Por otra parte los crustáceos, en la estación seca, constituyeron el 19.05% y en la húmeda, el 26.19%. Patrón similar se encontró para aves, mamíferos e insectos. Los reptiles únicamente aparecieron durante la estación seca (Cuadro 1C).
Entre los vegetales, los frutos de guázima, guamúchil, papaya, coco y las gramíneas sumaron las mayores frecuencias de consumo con más del 35%. Durante la estación húmeda predominaron el mabolo (Morisonia americana), el coco y el maíz (Zea mays). En cambio durante la estación seca el guamúchil, la guázima y las gramíneas fueron los elementos más consumidos (Cuadro 4).
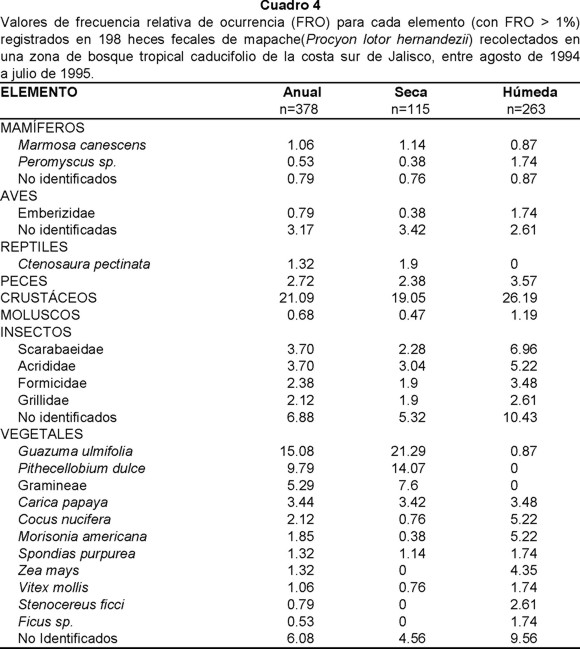
Los insectos como chapulines, escarabajos, grillos y hormigas, sumaron más de 11% del total de FRO y fueron dominantes. A diferencia de los vegetales, estos elementos mostraron una mayor variedad en la estación seca que en la húmeda. Entre las aves, los de la familia Emberizidae fueron los más consumidos a lo largo del año. Sin embargo estacionalmente su frecuencia de consumo fue mayor en la estación húmeda que en la seca (Cuadro 4).
Por otro lado, el ratón tlacuache (Marmosa canescens) y Peromyscus sp. fueron los mamíferos de mayor FRO con el 1.59%. En la estación húmeda Peromyscus sp. alcanzó 1.74%, en cambio en la estación seca el ratón tlacuache tuvo el 1.14% (Cuadro 4).
Dieta del Jaguarundi
Para el jaguarundi se colectaron 36 heces fecales durante todo el periodo de estudio. Los mamíferos (43.37%) constituyeron el grupo más consumido, seguido por vegetales (26.51%), insectos (16.87%) y aves (7.23%). En todos los excrementos recolectados se registraron mamíferos, lo que indica su importancia en la dieta de esta especie. En ambas estaciones mamíferos, vegetales e insectos se muestran como las categorías más frecuentemente consumidas; aunque en la húmeda los mamíferos fueron más consumidos con respecto a la seca (46.51% y 40% respectivamente). Un patrón similar se presentó para insectos, pero no así para vegetales que tuvieron un patrón inverso (Cuadro 1D).
El 53% de los mamíferos registrados lo conformaron la rata arrocera (O. couesi) (15.18%), rata algodonera (S. mascotensis) (9.82%), Peromyscus sp. (8.04%) y ratón espinoso (L. pictus) (5.36%). En la estación seca, Peromyscus sp. (12.72%), rata algodonera (9.09%), rata arrocera (9.09%) y ratón espinoso (7.27%) fueron las dominantes. En cambio, para la húmeda, la rata arrocera (21.05%) y la rata algodonera (10.53%) registraron las mayores FRO (Cuadro 5).
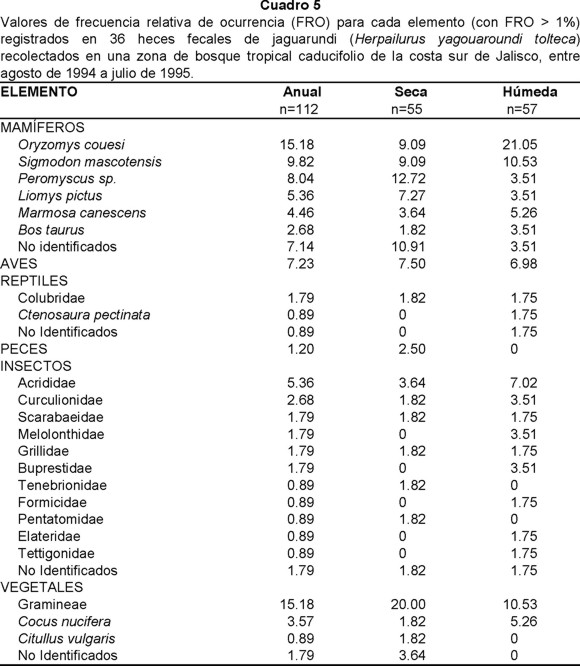
De los vegetales registrados en la dieta del jaguarundi, las gramíneas constituyeron 15.18%, seguida por el fruto de coco (3.57%). Estacionalmente, las gramíneas siguen siendo el elemento dominante con 20% en la estación seca y 10.53% en la húmeda (Cuadro 5).
Los chapulines (5.36%) y gorgojos (2.68%) fueron los insectos más frecuentes. La variedad de insectos fue mayor en la estación húmeda que en la seca, observándose que en esta última, seis de los siete elementos registraron 1.75%. Los chapulines fueron los más consumidos con valores de 7.02% en la estación húmeda y 3.64% en la seca. En la húmeda sobresalieron, además de los antes citados, los mayates (3.51%), los gorgojos (3.51%) y los buprestidos (Buprestidae) (3.51%) (Cuadro 5).
El resto de la dieta estuvo compuesta por aves y reptiles. De estos últimos, las culebras (Colubridae) fueron las más consumidas, sobre todo en la estación seca en la cual únicamente se registraron componentes de esta familia. En cambio la iguana negra sólo se presentó en la estación húmeda.
Nicho de alimentación
De acuerdo a los resultados obtenidos de amplitud del nicho de alimentación anual, tanto la zorra gris (15.32) como el coyote (14.95) presentaron el más amplio, seguidos por el mapache (11.16) y el jaguarundi (4.46) con los más estrechos. Estacionalmente este patrón se modificó, registrandose para la estación seca a la zorra gris con el nicho más amplio (16.04) y el más estrecho fue para el mapache (8.83); en cambio durante la estación húmeda el coyote fue la especie con mayor amplitud en su nicho (14.42) y la de menor amplitud fue para el mapache (10.46) (Cuadro 6).
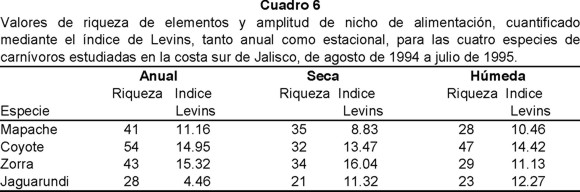
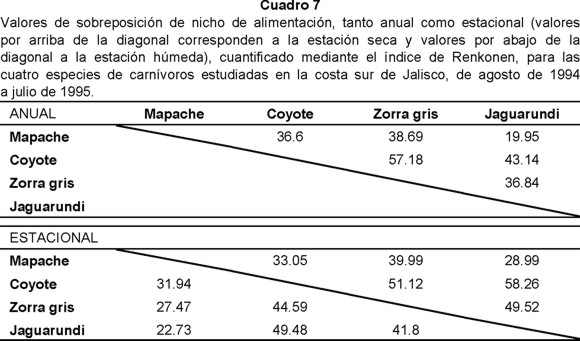
Con relación a la sobreposición o empalme del nicho, el análisis anual arrojó como resultado que el coyote y la zorra (57.18%) registraron la mayor sobreposición de nicho, en cambio el menor fue entre mapache y jaguarundi (19.95%). Durante la estación seca las especies con el mayor traslape fueron coyote y jaguarundi (58.26%) y las de menor mapache y jaguarundi (28.99%). Para la estación húmeda esta tendencia se mantuvo siendo en el caso del primer par de especies 49.48% y del segundo 22.73% (Cuadro 7).
Discusión
El coyote se caracteriza por su adaptabilidad a diferentes condiciones de hábitat, lo que se ve reflejado en su dieta, esto acorde con la disponibilidad de recursos en los sitios en que habita. En los estudios de Ozoga & Harger (1966), Gipson (1974), MacCraken & Uresk (1984) y Andelt (1985), se describe que, en bosques templados, los mamíferos son su principal fuente de alimento, específicamente venado cola blanca, conejos y roedores. Sin embargo, en el norte y centro de la República Mexicana, los estudios de Servín & Huxley (1991), García-Hernández (1994) y Aranda et al. (1995) indican que son los roedores y lagomorfos los que registran las mayores frecuencias de consumo. Ewer (1973) señala que cuando los insectos y las plantas son abundantes, suelen ser consumidas por el coyote. A diferencia de los anteriores, el presente estudio reveló que en el bosque tropical, la categoría referida como material vegetal (frutos) fue la más frecuentemente consumida por el coyote, seguido de los roedores, reflejando la adaptabilidad de esta especie a los recursos disponibles en el sitio en donde habita. Los bosques tropicales tienen una considerable variedad y riqueza de recursos alimentarios, entre ellos los frutos, lo cual permite que, especies como el coyote, puedan hacer uso de ellos durante las temporadas de fructificación y abundancia.
El aumento de consumo de frutos y mamíferos en la dieta del coyote durante la estación seca y el de aves e insectos en la húmeda, es un patrón que ha sido encontrado en otros trabajos. Servín & Huxley (1991) en el bosque templado registraron que, los mamíferos fueron el principal alimento durante el invierno y primavera (época seca), mientras que los frutos lo fueron en el verano (época húmeda). Por su parte Andelt (1985), en una zona árida, reportó que los mamíferos constituyeron el 87% de la dieta durante el invierno y 28% en el verano. Frutos y tunas formaron el 65% de la dieta durante el verano y únicamente el 1% de la dieta en invierno, en cambio los insectos, principalmente chapulines, formaron parte de la dieta a finales del verano. Estos cambios estacionales en su dieta parece ser una forma de compensar los requerimientos nutricionales para sobrevivir y muestra su capacidad de adaptación a los recursos disponibles. Es por esto que se ha catalogado al coyote como generalista (Ozoga & Harger 1966, Litvaitis & Shaw 1980), y forrajeador oportunista (Servín & Huxley 1991).
En la zona de Tenacatita se observó que las mayores frecuencias de ocurrencia anuales en las heces fecales de zorra gris recolectadas, fueron de materia vegetal, seguido de insectos y mamíferos. A diferencia de nuestros resultados, Errington (1935) y Wood (1959) señalan que esta especie tiene preferencia por los pequeños mamíferos, como lepóridos y roedores. En cambio Kodani (1995), encontró que elementos vegetales, mamíferos, insectos y aves fueron dominantes en la dieta de esta especie en su zona de estudio en California. Estas variaciones registradas entre los estudios realizados en bosques templados y el presente, realizado en el bosque tropical caducifolio, son un indicador de la adaptabilidad de la zorra gris que, al igual que el coyote, es considerada altamente adaptable a diferentes ambientes y sus cambios.
Dicha adaptabilidad de la zorra gris, es aún más evidente con los cambios estacionales en su dieta. En la estación húmeda, cuando los frutos e insectos son abundantes, éstos fueron más frecuentemente consumidos. En tanto en la estación seca, cuando frutos e insectos disminuyen, los mamíferos predominaron en su dieta, lo cual es una manera de responder al cambio que ocurre en los recursos de mayor disponibilidad estacional en su hábitat.
El mapache, tiende a vivir cerca de lagunas, presas o ríos, de los cuales obtiene sus alimentos preferidos como, crustáceos, anfibios y bivalvos, pasando a segundo término los frutos (Dearborn 1932, Stuewer 1943, Cagle 1949, Hamilton 1951, Schoonover & Marshall 1951, Tester 1953, Wood 1954, Gander 1966), lo cual concuerda con los resultados obtenidos en este estudio, en donde los frutos, crustáceos e insectos fueron consumidos.
En los trabajos de Stuewer (1943), Cagle (1949), Hamilton (1951), Schoonover & Marshall (1951), Tester (1953) y Gander (1966) se encuentran cambios estacionales en la dieta del mapache, lo cual incluye vegetales, crustáceos y ocasionalmente insectos y mamíferos. Esta descripción es semejante a lo obtenido en este estudio para el bosque tropical caducifolio.
Para el jaguarundi son escasos los estudios relacionados con su dieta pero, al igual que otros felinos (p. e. ocelote y tigrillo) que habitan la zona, los mamíferos son la base de su alimentación, principalmente los roedores. En el presente estudio, el jaguarundi fue la especie con el menor valor en cuanto a diversidad de elementos encontrados en sus heces fecales, destacando los pequeños mamíferos, materia vegetal e insectos. Esta especie ha sido señalada por campesinos que habitan el área de estudio como depredadora de aves de corral, en este trabajo la FRO de aves fue de apenas 5.36%, no apareciendo las aves de corral, lo cual descarta el papel de depredador nocivo para especies domésticas usadas por los pobladores locales. La frecuencia de depredación se incrementó durante la estación húmeda tanto sobre mamíferos como sobre insectos, mientras que sobre material vegetal disminuyó. Esto sugiere que los materiales vegetales forman un complemento básico en su dieta pero su preferencia es consumir proteína animal.
En diferentes estudios y regiones se ha destacado la capacidad de adaptación de la zorra gris y el coyote a distintas condiciones del ambiente, llegando a ser catalogados como oportunistas y generalistas (Leopold 1977, Bekoff 1977, Fritzell & Haroldson 1982, Servín & Huxley 1991). El registro del nicho de alimentación más amplio para ambas especies, durante este estudio, corrobora ésta hipótesis. Su capacidad adaptativa se vio igualmente reflejada estacionalmente, al ser ambas especies las que mostraron los valores mayores de nicho, ello sugiere que su espectro alimentario es el más amplio de los carnívoros estudiados, lo que les otorga ventajas importantes para su sobrevivencia, también permite explicar el hecho de que sean de las especies de carnívoros más abundantes, no sólo en la zona de estudio (López-Acosta & Guerrero 1994, Ordorica-Hermosillo 1996), sino también en otros ambientes.
El mapache y el jaguarundi, requieren de condiciones más estrictas en su hábitat. La primera de ellas (mapache) preferentemente habita en sitios donde la presencia de agua es permanente (Leopold 1977), y a cuyos ambientes esta ligada su especialización alimentaria. Por su parte el jaguarundi, es una especie cuyos requerimientos ecológicos son más estrictos respecto a la zorra gris y el coyote, lo que se refleja en un nicho de alimentación más estrecho, sugiriendo esto un alto grado de especialización trófica, comparada con las otras tres especies. Aún cuando el jaguarundi es una especie con un alto grado de especialización en su dieta, durante la estación húmeda fue la segunda con mayor amplitud de nicho. Si bien el consumo de los pequeños mamíferos, que son la base de su dieta, se mantuvo constante entre ambas estaciones, la presencia de insectos fue mayor en la húmeda, lo que ocasionó un incremento en el espectro de elementos consumidos. Ello habla de que a pesar de su preferencia por ciertas presas, en un momento dado puede incorporar otras que le permiten complementar sus requerimientos energéticos.
No obstante que el mapache y el jaguarundi fueron las especies cuyo valor del nicho de alimentación es el más estrecho de las cuatro estudiadas, también fueron las especies que registraron el menor valor de sobreposición, tanto anual como estacional. Esto muestra que cada una tiene requerimientos de alimentación muy particulares. Mientras que el mapache tiene una mayor preferencia por frutos y crustáceos, el jaguarundi tiene predilección por los pequeños mamíferos.
En cambio, entre la zorra y el coyote existe una notable afinidad en su dieta, al ser también las de mayor sobreposición, con base en los datos anuales. Material vegetal, insectos y mamíferos constituyeron los grupos base de la dieta para ambas especies. Sin embargo, estacionalmente se observó que entre el coyote y el jaguarundi se registró el mayor grado de sopreposición, tanto para la estación seca como para la húmeda. Esto, en cierta medida, esta dado porque ambas especies tienen una fuerte afinidad por el consumo de pequeños mamíferos. Si bien es cierto que en el caso del coyote el material vegetal resultó ser el de mayor FRO, esto se presenta por la alta cantidad de gramíneas. En algunos trabajos se sugiere que son consumidas de manera ocasional y no como una fuente de alimento (Aranda et al. 1995), si se excluyeran éstas del análisis de dieta para el coyote, el resultado sería que los mamíferos representan el grupo más relevante por su FRO, lo que daría más claridad al alto grado de sobreposición de nicho mostrado entre coyote y jaguarundi.
Este análisis mostró que tanto la zorra gris como el coyote son especies generalistas, con base en los valores de amplitud de nicho mostrados. En cambio, el jaguarundi es una especies especialista, cuya dieta esta basada en gran medida en pequeños mamíferos. Por otro lado, aún cuando la relación trófica entre estas especies es notable, particularmente entre coyote, zorra gris y mapache, los sitios en donde obtienen sus recursos puede ser el factor que evite la competencia y por lo tanto permita su coexistencia. El caso más claro se presenta entre mapache y jaguarundi, ya que cada una prefiere diferentes ambientes (Zalapa et al. 1998) sobre los cuales está ligada la obtención de su alimento.
Agradecimientos
Agradecemos a Ma. Rosario Sandoval y a Antonio Ordorica su apoyo durante el trabajo de campo y en el trabajo de laboratorio. Así mismo a Agustín Camacho Rodríguez, José Luis Navarrete-Heredia, Martín Huerta-Martínez, Rosio Amparán Salido y a Jorge Tellez López por su apoyo para la determinación del material. A Arturo Jiménez-Guzmán por sus sugerencias al presente manuscrito. También deseamos hacer patentes nuestros agradecimientos por los comentarios y sugerencias a Carolina Valdespino y Jorge Servín que permitieron enriquecer y mejorar en gran medida este manuscrito.
Literatura citada
Ackerman, B. B., F. G. Lindzey, & T. P. Hemker. 1984. Cougar food hHabits in Southern Utah. J. Wildl. Manage. 48(1):147-155. [ Links ]
Andelt, W. f. 1985. Behavioral ecology of coyotes in South Texas. Wildl. Monogr. No. 94, Pp. 45. [ Links ]
Andelt, W. F., J. G. Kie, F. F. Knowlton & K. Cardwell. 1987. Variation in coyote diets associated with season and successional change in vegetation. J. Wildl. Manage. 51(2):273-277. [ Links ]
Aranda, J. M. 1981. Rastros de los mamíferos silvestres de México. INIREB, México. 178 pp. [ Links ]
Aranda, M., N. López-Rivera & L. López-de Buen. 1995. Hábitos alimentarios del coyote (Canis latrans) en la Sierra del Ajusco, México. Acta Zool. Mex. (n.s.), 65:89-99. [ Links ]
Arita, H. T. & M. Aranda. 1987. Técnicas para el estudio y clasificación de los pelos. Cuadernos de Divulgación INIREB No. 32, INIREB, Xalapa, Veracruz. Pp. 21. [ Links ]
Badii, M. H., M. Villa, D. Lazcano & H. Quiróz. 1992. Análisis conceptual del nicho alimenticio y diversidad intraespecífica ejemplificada mediante dos especies de lagartijas. Pub. Biol.- F.C.B./U.A.N.L. 6(1):65-69. [ Links ]
Bekoff, M. 1977. Canis latrans Say. Mamm. Spec. 79:1-9. [ Links ]
Cagle, F. R. 1949. Notes on the raccoon, Procyon lotor megalodous, Lowery. J. Mammal. 30:45-47. [ Links ]
Ceballos, M. & A. Miranda. 1986. Los mamíferos de Chamela, Jalisco. Ed. Limusa-Instituto de Biología, UNAM, México. 436 pp. [ Links ]
Dearborne, N. 1932. Foods on some predatory fur-bearing animal in Michigan. Bull. Sch. For. Conserv. Univ. Michigan. I:1-52. [ Links ]
Delibes, M. & F. Hiraldo. 1987. Food habits of the bobcat in two habitats of the Southern Chihuahua Desert. The Southwestern Naturalist. 32(4):457-461. [ Links ]
Errington, P. l. 1935. Food habits of mid-west foxes. J. Mammal. 18:203-205. [ Links ]
Esparza-garcia, J. A. 1991. Variaciones estacionales en la dieta de mamíferos carnívoros en la estación científica Las Joyas. Tesis de Licenciatura, Facultad de Ciencias Biológicas, U. de G. Inédita. [ Links ]
Ewer, R. F. 1973. The carnivores. Cornell University Press, New York. 494 pp. [ Links ]
Fitts, S. H. & J. A. Sealander. 1978. Diets of bobcats in Arkansas with special reference To Age And Sex Differences. J. Wildl. Manage. 42(3):533-539. [ Links ]
Fritzell, E. K. & K. J. Haroldson. 1982. Urocyon cinereoargenteus. Mamm. Spec. 189:1-8. [ Links ]
Gander, F. F. 1966. Raccoon. Animal Kingd. 69:84-89. [ Links ]
Garcia-Hernández, R. E. 1994. Análisis preliminar de la dieta invernal del coyote (Canis latrans Say 1823). Estudio comparativo en dos áreas del rancho El Macho, Guerrero, Coahuila, México. Tesis profesional, Fac. Ciencias Biol., UANL. Inédita. [ Links ]
Garrott, R. A., L. E. Eberhardt, & W. C. Hanson. 1983. Summer food habits of juvenile arctic foxes in Northern Alaska. J. Wildl. Manage. 47(2):540-544. [ Links ]
Giller, P. S. 1984. Community structure and the niche. Chapman and Hall, New York. 176 pp. [ Links ]
Gipson, P. S. 1974. Food habits of coyotes in Arkansas. J. Wildl. Manage. 38(4):848-853. [ Links ]
Graff, S. H. 1988. Fauna silvestre en el bosque La Primavera; hábitos de alimentación del coyote (Canis latrans) y zorra gris (Urocyon cinereoargenteus). Tesis profesional, Facultad de Agronomía, U. de G. Inédita. [ Links ]
Hamilton, W. J. 1951. Warm-weather food of the raccoon in New York State. J. Mammal. 32:341-344. [ Links ]
Halfpenny, J. & E. Biesot. 1986. A field guide to mammal tracking in North América. Second Edition. Johnson Publishing Company. USA. 161 pp. [ Links ]
Hutchinson, G. E. 1957. Concluding remarks. Cold Spring Harbor Symposia in Quantitative Biology. 22:415-427. [ Links ]
Kodany, C. H. 1995. Home range and diet of California gray fox (Urocyon cinereoargenteus californicus) in the Cleveland National Forest, Orange County, California. Thesis of Master of Arts in Biology, California State University, Fullerton. 49 pp. [ Links ]
Korschgen, L. J. 1980. Procedimiento para el análisis de los hábitos alimentarios. En: T. R. Rodríguez (ed.). Manual de gestión de vida silvestre, Cuarta Edición, USA. 119-134. [ Links ]
Krebs, C. J. 1999. Ecological methodology. Addison Wesley Longman, Inc., CA., USA. 620 pp. [ Links ]
Leopold, A. S. 1977. Fauna silvestre de México. Instituto Mexicano de Recursos Naturales Renovables, Ed. Pax-México 2da Ed. México. 608 pp. [ Links ]
Levins, R. 1968. Evolution in changing environments. University Press, Princenton, New Yersey, USA. [ Links ]
Litvaitis, J. A. & J. H. Shaw. 1980. Coyote movements, habitat use, and food habits in southwestern Oklahoma. J. Wildl. Manage. 44(4):62-68. [ Links ]
López-Acosta, M. A. & S. Guerrero. 1994. Notas sobre mamíferos de la costa sur de Jalisco, México. BIOTAM, 6(1):29-38. [ Links ]
Maccraken, J. G. & D. W. Uresk 1984. Coyote foods in the Black Hills, South Dakota. J. Wildl. Manage. 48(4):1420-1423. [ Links ]
Magurran, A. 1989. Diversidad ecológica y su medición. Ed. Vedra, España. 197 pp. [ Links ]
Ordorica-Hermosillo, A. R. 1996. Estimación de la densidad relativa de carnívoros (Mammalia:Carnivora) en Tenacatita, Jalisco. Tesis Profesional, División de Ciencias Biológicas, CUCBA, U. de G. Inédita. [ Links ]
Ozoga, J. J. & E. M. Harger. 1966. Winter activities and feeding habits of northern Michigan coyote. J. Wildl. Manage. 30(40):809-818. [ Links ]
Schoonover, L. J. & W. H. Marshall, 1951. Food habits of raccoon (Procyon lotor hirtus) in north-central Minnesota. J. Mammal. 32:422-428. [ Links ]
Servín, J. & C. Huxley. 1991. La dieta del coyote en un bosque de encino-pino de la Sierra Madre Occidental de Durango, México. Acta Zool. Mex. (n. s.) 44:1-26. [ Links ]
Smith, E. P. 1982. Niche beadth, resource availability, and inference. Ecol. 63(6):1675-1681. [ Links ]
Stuewer, F. W. 1943. Raccoons: their habits and management in Michigan. Ecol. Monogr. 13:203-257. [ Links ]
Tester, J. R. 1953. Fall foods of the raccoon on the South Plate Valley of northeastern Colorado. J. Mammal. 34:500-502. [ Links ]
Todd, A. W., L. B. Keith & C. A. Fischer. 1981. Populations ecology of coyotes during a fluctuation of snowshoe hares. J. Wildl. Manage. 45(3):629-640. [ Links ]
Vela-Coiffer, E. l. 1985. Determinación de la composición de la dieta del coyote (Canis latrans Say), por medio del análisis de heces fecales en tres localidades del estado de Chihuahua. Tesis de Licenciatura, Facultad de Ciencias Biológicas, UANL. Inédita. [ Links ]
Wood, J. E. 1954. Food habits of furbearers of the uplands post oak region in Texas. J. Mammal. 35:406-414. [ Links ]
––––––––––. 1959. Relative estimates of fox population levels. J. Wildl. Manage. 23(1):53-63. [ Links ]

