










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkActa zoológica mexicana
versão On-line ISSN 2448-8445versão impressa ISSN 0065-1737
Acta Zool. Mex no.80 Xalapa Ago. 2000
Artículo
Determinación de la dieta del mapache (Procyon lotor hernandezii Wagler, 1831) en la costa sur de Jalisco, México
Sergio Guerrero, Ma. Rosario Sandoval y Silvia S. Zalapa
Centro de Estudios en Zoología, División de Ciencias Biológicas, Centro Universitario de Ciencias Biológicas y Agropecuarias, U. de G. Apdo. Postal 1-1919, CP 44101 Guadalajara, Jalisco, México.
Recibido: 24 de febrero 1999
Aceptado: 26 de enero 2000
Resumen
En este trabajo se dan a conocer algunos aspectos de la dieta del mapache (Procyon lotor hernandezii) en la zona de Tenacatita, en el Estado de Jalisco. Se realizaron colectas de excrementos mensualmente, de marzo de 1994 a diciembre de 1995, a partir de los cuales se determinó que el material vegetal representó el 41.0%, los insectos el 18.4% y los crustáceos el 17.8% de frecuencia relativa de ocurrencia (FRO). Entre los vegetales, guázima (Guazuma ulmifolia), guamuchil (Pithecellobium dulce), gramíneas, papaya (Carica papaya) y coco (Cocus nucifera) representaron el 70%. Cuatro plantas que se cultivan en la zona fueron registradas en las heces fecales de esta especie con FRO bajas: papaya (3.93%), coco (1.54%), maíz (1.03%) y mango (0.68%). Entre los insectos sobresalieron chapulines (Acrididae), escarabajos (Scarabaeidae), grillos (Grillidae) y hormigas (Formicidae). Se observaron variaciones estacionales en la FRO de algunas categorías de alimento como en vegetales e insectos. La diversidad de elementos contenidos en la dieta alcanzó un valor de H'=1.36.
Palabras clave: Mapache, dieta, cultivos, diversidad.
Abstract
In this work, we documented the food habits of the Raccoon (Procyon lotor hernandezii) in the zone of Tenacatita, in the State of Jalisco. We collected scats each month, from March 1994 to December 1995. We determined that plant material was first in importance at 41.0%, the next was insects with 18.4%, and crustaceans with 17.8% of relative frequency of occurrence (RFO). The plants with higher frequencies were guazima (Guazuma ulmifolia), guamuchil (Pithecellobium dulce), grasses (Gramineae), papaya (Carica papaya) and coconut (Cocus nucifera). Four cultivated plants were found in the scats: papaya (3.93%), coconut (1.54%), corn (1.03%) and mango (0.68%), but their RFO was not high. Of the insects consumed, grasshoppers (Acrididae), beetles (Scarabaeidae), crickets (Grillidae), and ants (Formicidae) predominated. Seasonal variations in the RFO of plant material and insects were observed. The diversity of food items overall was H'= 1.36.
Key words: Raccoon, food habits, field crops, diversity.
Introducción
El mapache (Procyon lotor), es una especie con una amplia distribución y capacidad de adaptación a distintos ambientes (Leopold 1977). En la costa sur de Jalisco es común (Ceballos y Miranda 1986, López-Acosta y Guerrero 1994, Ordorica-Hermosillo 1996); sin embargo, es poca la información que se tiene sobre su ecología en esa zona en particular y, en general, en el Estado de Jalisco.
El mapache vive prácticamente en cualquier tipo de ambiente, siempre y cuando tenga agua cercana, de preferencia durante todo el año. En épocas de abundancia de frutos suele alejarse del agua, pero durante la temporada seca regresa a los lugares húmedos de su hábitat donde siempre obtiene comida (Leopold 1977).
Entre los aspectos mejor conocidos de esta especie se encuentra su dieta (Ewer 1973). Es considerado omnívoro, ya que incluye en su alimentación mamíferos, ranas, reptiles, crustáceos, peces, insectos, frutos y semillas (Leopold 1977, Ceballos y Galindo 1984, Guerrero et al. 1995); de esta variedad en la dieta depende gran parte su adaptabilidad (Leopold 1977). Diversos estudios (Stuewer 1943, Cagle 1949, Schoonover y Marshall 1951, Tester 1953 y Gander 1966) destacan la variación que existe por su dieta en relación con la disponibilidad de alimento en las distintas estaciones del año. Así mismo, se ha resaltado su predilección por los granos cultivados, entre ellos el maíz, lo que la hace potencialmente dañina para los campos de cultivo (Gander 1966, Ceballos y Galindo 1984), sobre todo donde los cambios en el uso del suelo han favorecido el incremento de sus poblaciones.
En estudios realizados en la zona de Tenacatita en el Estado de Jalisco (López-Acosta y Guerrero 1994, Guerrero et al. 1995 y Ordorica-Hermosillo 1996) se observó que el mapache se encuentra entre los carnívoros más comunes, siendo señalado por los pobladores como causante de daños a los cultivos de maíz y papaya. El objeto del presente trabajo es describir algunos aspectos de su dieta y con ello contribuir al conocimiento de su ecología.
Descripción del área de estudio
El área de Tenacatita es una llanura costera con una extensión de aproximadamente 4,000 ha, localizada en el Municipio de La Huerta, Jalisco. Topográficamente está limitada al E por el parteaguas de los cerros y lomeríos de la llanura de La Manzanilla y la Bahía Tenacatita; al W con el parteaguas de los cerros y lomeríos de la llanura costera El Tecuán y la albufera La Fortuna; al N por los lomeríos de la zona de Miguel Hidalgo y el poblado Aguacaliente y al S por el Océano Pacífico. Sus coordenadas geográficas son 19/16' y 19/21' de latitud N y 104/51' y 104/55' de longitud W (López-Acosta y Guerrero 1994).
La vegetación de la región está compuesta por selva baja caducifolia, manglares, selva mediana subcaducifolia, pastizales y vegetación halófita. Los cultivos son comunes en el área, particularmente los cocoteros, platanares y papaya, entre los de tipo permanente, y maíz entre los de temporal (Ceballos y Miranda 1986, López-Acosta y Guerrero 1994).
Material y métodos.
Se realizaron recolectas de excrementos, cada uno considerado como una muestra, en los caminos y brechas presentes en la zona de estudio, de marzo de 1994 a diciembre de 1995, con lo que se cubrieron dos estaciones: la seca (marzo-mayo de 1994 y enero-mayo de 1995) y la húmeda (junio-diciembre de 1994 y 1995).
Para la identificación de los excrementos se aplicaron los criterios señalados por Aranda (1981), así como también se consideró la presencia de huellas de la especie en el sitio de recolecta. Con el objeto de evitar sesgo, aquellas sobre las que se tuvo duda en su identificación fueron descartadas del análisis
Las muestras fueron secadas a temperatura ambiente; ya en el laboratorio, se terminó su deshidratación en una estufa de secado. Se depositaron en bolsas de manta (debidamente rotuladas) para su lavado en una solución a base de agua y detergente, hasta que fue removido todo el material soluble. Una vez limpias, se procedió a su secado por medio de pistola de aire caliente. Los materiales resultantes contenidos en los excrementos se separaron y colocaron en bolsas de plástico para su posterior determinación.
La determinación se realizó al nivel más específico posible. Para determinar los mamíferos se usaron características del pelo (escamas y médula) (MacCraken y Uresk 1984) aplicando el método propuesto por Arita y Aranda (1987). El material óseo (principalmente dientes) se comparó con ejemplares recolectados en la zona, los cuales se encuentran depositado en la Colección de Vertebrados del Centro de Estudios en Zoología de la Universidad de Guadalajara (CZUG). Los insectos, crustáceos, peces, aves y material vegetal, fueron determinados con el apoyo de investigadores del CZUG. Los peces se determinaron mediante las escamas presentes en las muestras y las aves mediante las plumas, principalmente; en ambos casos la comparación se hizo con material recolectado en la zona de estudio.
Los resultados son presentados como frecuencia relativa de ocurrencia (FRO) y porcentaje de ocurrencia (PO), siguiendo los criterios utilizados por Servín y Huxley (1991) y Aranda et al. (1995). La FRO fue calculada como FRO=(fi/∑fi)(100), en donde fi es la frecuencia absoluta de un componente " i ". El PO, fue calculado como PO=(fi/N)(100), en donde fi es la ocurrencia de cada componente " i ", y N el número total de muestras. La diversidad de elementos contenidos en la dieta del mapache fue cuantificada mediante el índice de Shannon:
H'=–∑Pi logPi
en donde Pi es la proporción de la frecuencia registrada para el elemento "i". Este cálculo se hizo tanto a nivel global como estacional (considerando de manera conjunta los datos de 1994 y 1995 de las estaciones respectivas). Para determinar si los valores de diversidad eran significativamente diferentes entre estaciones, se aplicó una prueba t (Zar 1996). Así mismo se cuantificó la diversidad máxima mediante la fórmula:
Hmax = LogS
en donde S es la riqueza de elementos, y la equitatividad se calculó como:
E = H'/Hmax (Magurran 1989).
Resultados
Se analizaron 260 excrementos. Vegetales (41.0%), insectos (18.4%) y crustáceos (17.8%) fueron los componentes más frecuentes en las heces fecales de esta especie. Aves (6.6%), mamíferos (2.9%), peces (3.9%) y reptiles (1.8%) se consideraron como un complemento de la dieta por sus bajos porcentajes (Cuadro 1).
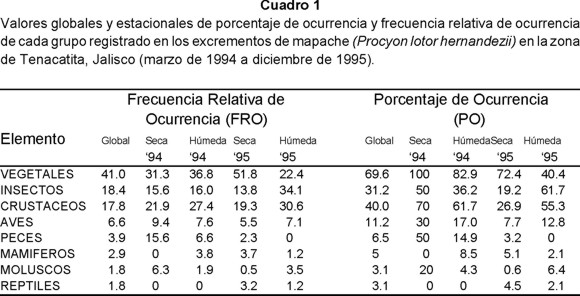
Los vegetales fueron más frecuentes en la estación húmeda de 1994 (36.8%) y en la estación seca de 1995 (51.8%). Las FRO más altas de insectos y crustáceos se registraron en la estación húmeda de ambos años en comparación con la estación seca; en el caso de los primeros con 16.0% (húmeda 94) y 34.1% (húmeda 95) y en el de los segundos con 27.4% (húmeda 94) y 30.6% (húmeda 95). Entre las estaciones se registraron FRO diferentes de reptiles, mamíferos, aves, moluscos y peces, siendo notable el valor registrado para este último grupo durante la estación seca de 1994 en la cual alcanzó un 15.6%. Los reptiles únicamente aparecieron en las muestras recolectadas en 1995, siendo mayor su frecuencia en la estación seca (3.2%) que en la húmeda (1.2%)(Cuadro 1).
A continuación se describen los elementos identificados en los excrementos y sus valores de FRO y PO.
Vegetales.- Constituyeron el grupo con mayor FRO en las heces fecales del mapache. Guázima (Guazuma ulmifolia), guamuchil (Pithecellobium dulce), gramíneas, papaya (Carica papaya) y coco (Cocus nucifera) registraron en conjunto las mayores frecuencias con más del 70% del material vegetal, lo que evidencia su importancia para esta especie (Cuadros 2 y 3).
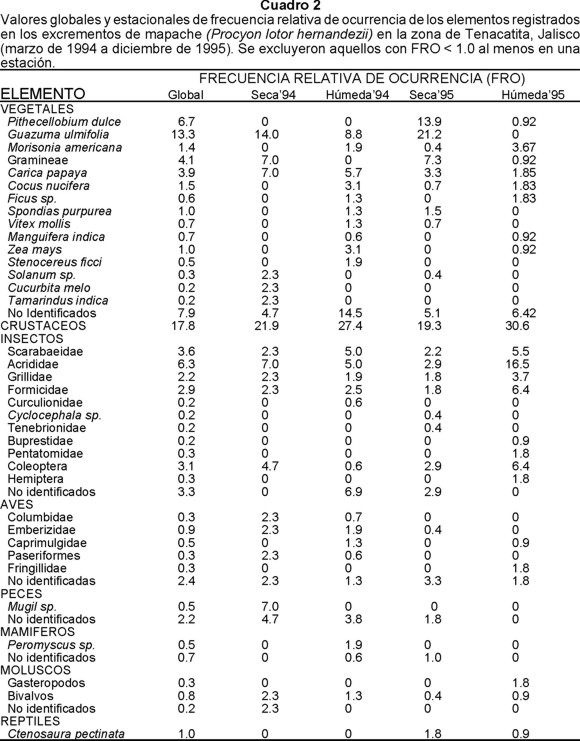
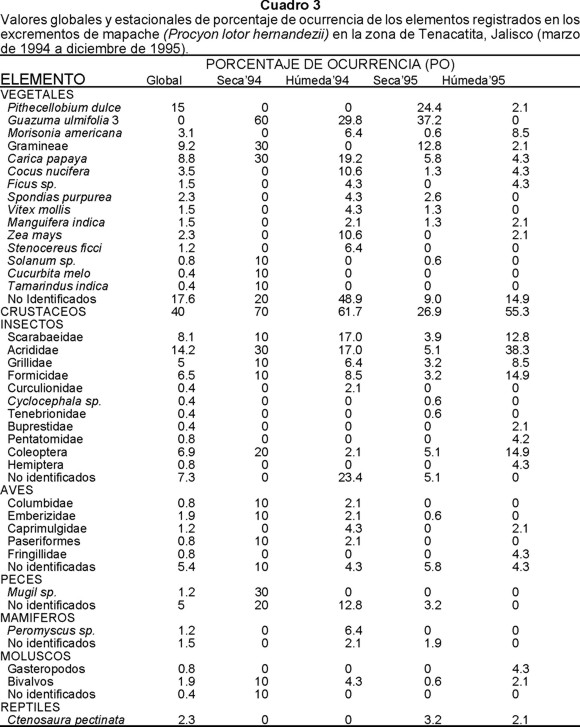
En las heces fecales recolectadas durante las estaciones húmedas se registró una mayor riqueza de elementos, en la que predominaron guázima, papaya, mabolo (Morizonia americana), coco y maíz (Zea mays). En cambio en las estaciones secas fueron más frecuentes guázima, guamuchil, gramíneas, papaya y ciruela (Spondias purpurea); tanto guázima como guamuchil fueron un componente importante durante la estación seca de ambos años, alcanzando el primero el 35% del material vegetal registrado en 1994 y en 1995 ambos alcanzaron 64.4% (Cuadros 2 y 3).
De las plantas cultivadas que se identificaron en las muestras, para la papaya se registró, de manera global, el valor más alto con 3.9%, seguida del coco con 1.5% y mango con 0.7%. En el caso del maíz, se obtuvo un valor de 1.0% a lo largo de todo el periodo de estudio; esta planta fue registrada sólo en las estaciones húmedas con porcentajes bajos tanto de 1994 como de 1995 (Cuadro 2). En términos del PO global, 2.3 de cada 100 muestras contuvieron maíz, pero en la estación húmeda de 1994 se encontró que 10.6 de cada 100 muestras lo contenían (Cuadro 3).
Insectos.- Fue el segundo grupo en importancia por su FRO. Sobresalieron chapulines (Acrididae), escarabajos (Scarabaeidae), grillos (Grillidae) y hormigas (Formicidae) que alcanzaron un 57% del total de insectos registrados.
Al igual que el material vegetal, los insectos mostraron una mayor riqueza en la estación húmeda que en la seca, aunque en ambas predominaron chapulines, escarabajos, grillos y hormigas. Además de estos elementos, en la estación seca se registraron, escarabajos tenebrios (Tenebrionidae) y Cyclocephala sp. (Melolonthidae), y en la estación húmeda, gorgojos o picudos (Curculionidae) (Cuadros 2 y 3).
Durante las estaciones húmedas de 1994 y 1995 las mayores FRO correspondieron a escarabajos (Scarabaeidae), chapulines (Acrididae) y hormigas (Formicidae), lo que indica que estos elementos fueron relevantes en la dieta del mapache durante esa estación (Cuadros 2 y 3).
Crustáceos.- Fue el tercer grupo en importancia alcanzando el 17.8% de FRO. Estacionalmente los crustáceos tuvieron mayores FRO en la estación húmeda de ambos años (27.4% en 1994 y 30.6% en 1995) que en la estación seca (21.9% en 1994 y 19.3% en 1995).
En orden de importancia, otros componentes de la dieta fueron: aves, mamíferos, reptiles, peces y moluscos.
Entre las aves la familia Emberizidae fue la más frecuente, registrándose en tres de las cuatro estaciones estudiadas en ambos años. Sin embargo, su frecuencia estacional no fue homogénea a lo largo del periodo de estudio, ya que su FRO fue mayor durante 1994 en ambas estaciones; en cambio en 1995 solamente se registró en la estación seca con un valor menor al 1%. Los grupos de aves aparecieron ocasionalmente a lo largo del periodo de estudio y no se observó un patrón definido entre estaciones para alguna de las familias identificadas en las muestras recolectadas.
De los mamíferos, Peromyscus sp. fue el elemento que registró la mayor FRO de manera global. En cambio estacionalmente fue durante la estación húmeda de 1994 cuando se observó el valor más alto.
Reptiles, peces y moluscos tuvieron bajas FRO. Los reptiles solamente se registraron en la estación seca.
La diversidad total de elementos contenidos en la dieta del mapache fue H'=1.36. Se observó un valor mayor durante la estación húmeda (H'=1.33) que en la estación seca (H'=1.25), pero no fueron significativamente diferentes (t = 0.79, P>0.05). La riqueza total de elementos fue de 59 y no fue significativamente diferente entre estaciones: 45 en la húmeda y 43 en la seca. La equitatividad total fue E= 0.77, con valores similares en la estación húmeda (E= 0.80) y en la seca (E= 0.77).
Discusión y conclusiones
A pesar de que el mapache tiene una amplia distribución dentro de la República Mexicana, ha sido relativamente poco estudiado. En Jalisco sus mayores densidades se han observado en la región de la costa, en donde se reporta como uno de los carnívoros más abundantes (Ordorica-Hermosillo 1996); si bien es común cerca de lagunas costeras y otros cuerpos de agua (Leopold 1977), también es frecuentemente observado en pastizales y zonas de cultivo. Análisis de preferencia de hábitat en el área de Tenacatita indican que utiliza la selva baja caducifolia, pastizales y cultivos en proporción a su disponibilidad (Zalapa et al. 1998). De acuerdo con habitantes de la zona de estudio, el mapache se ha convertido en un problema, ya que se le atribuyen daños a cultivos temporales como los de maíz (Z. mayz), y semiperennes como los de papaya (C. papaya).
Los resultados obtenidos en el presente estudio muestran que los elementos más frecuentes en su dieta fueron vegetales, insectos y crustáceos, lo cual coincide con lo reportado por Dearborne (1932), Stuewer (1943), Tester (1953), Wood (1954) y Gander (1966). Este último autor señala que la carroña es, tal vez, el único alimento que no consume, lo que habla de la variedad de elementos que acepta en su dieta. Nuestras observaciones confirman que es una especie omnívora.
Los cambios estacionales en la dieta del mapache no son algo fuera de lo común, particularmente en tipos de vegetación con un contraste climático bien definido, como lo es la selva baja caducifolia (Rzedowsky 1978). En este trabajo determinamos que durante la estación seca, cerca del 50% de los componentes de su dieta fueron de tipo vegetal, mientras en la estación húmeda el 70% fueron de tipo animal. Estas variaciones estacionales en el consumo de material vegetal y animal también han sido determinadas en estudios como los de Stuewer (1943), Hamilton (1951) y Schoonover y Marshall (1951).
Por otro lado, los cambios en el uso del suelo ocasionalmente suelen ser favorables para algunas especies, dada la disponibilidad asociada de una mayor cantidad de ciertos recursos alimentarios en determinadas épocas del año. Tal es el caso de las zonas de cultivo, ya sean temporales o permanentes. En el área de Tenacatita los principales cultivos de tipo permanente son el coco, el mango y la papaya, y entre los de temporal está el maíz. Se ha señalado que el mapache causa los mayores daños sobre estos cultivos. Sin embargo, los resultados obtenidos en este estudio muestran que la FRO de esos componentes en su dieta es baja. Para otras regiones se ha demostrado que cuando la disponibilidad de frutos silvestres disminuye, el maíz puede constituir una fuente importante de alimento para el mapache (Schoonover y Marshall 1951).
En la zona de estudio, el periodo de fructificación de los cultivos coincide con el de mayor variedad de recursos alimentarios disponibles en el hábitat natural del mapache, lo que disminuye la probabilidad de que este tenga un efecto adverso sobre las áreas de cultivo. Las bajas FRO de papaya y mango registradas en los excrementos, indican que la depredación sobre estos frutos quizá no es tan considerable como se supone. Sin embargo, un daño importante sobre la misma planta puede presentarse al ser derribada en el intento de obtener el fruto, como en el caso del maíz, aunque ésto también se observó en árboles de papaya. Cabe hacer notar que el daño en los cultivos probablemente es causado por un bajo porcentaje de la población de mapaches en el área.
El valor de H' indica que esta especie utiliza una amplia variedad de alimentos en su dieta, lo que también se reflejó en los valores estacionales. Dado que este tipo de índices es influido tanto por el número de elementos como por la equitatividad de los valores de importancia de los mismos (Magurran 1989), es recomendable contrastar la contribución de estos factores en la generación del valor del índice. En este caso tanto la riqueza como la equitatividad fueron muy semejantes estacionalmente, lo cual indica que el mapache tiende a equilibrar no sólo el número de elementos sino también la proporción de éstos en su dieta, lo que le permite compensar posibles cambios en la disponibilidad de ciertos recursos entre las estaciones. Dentro de esa variedad se registraron elementos que sobresalieron por su frecuencia en cada una de las estaciones: guázima, guamuchil, papaya, gramíneas y coleópteros durante la estación seca y escarabajos, chapulines, hormigas, coco, maíz, mabolo y crustáceos en la estación húmeda. Estos elementos pueden considerarse básicos en la dieta del mapache en el áea de estudio.
Agradecimientos
Agradecemos a Antonio Ordorica Hermosillo y a Miguel Angel López Acosta, por su apoyo y colaboración en el trabajo de campo. A Martín Huerta Martínez, José Luis Navarrete Heredia, Agustín Camacho Rodríguez, Rosio Amparán Salido, Jorge Téllez López y a Raymundo Ramírez, por su apoyo para la determinación del material. Así mismo a Fernando A. Cervantes, José Luis Navarrete Heredia, Gustavo Moya Raygoza y a dos revisores anónimos, por los comentarios y sugerencias hechos al presente manuscrito. Deseamos también agradecer a la Familia López de El Rebalsito, Jalisco, su hospitalidad durante el periodo de trabajo de campo.
Literatura citada
Aranda, M. 1981. Rastros de los mamíferos silvestres de México. INIREB, México. 178 pp. [ Links ]
Aranda, M., N. López-Rivera & L. López-de Buen. 1995. Hábitos alimentarios del coyote (Canis latrans) en la Sierra del Ajusco, México. Acta Zool. Mex. (n.s.) 65:89-99. [ Links ]
Arita, H.T. & M. Aranda. 1987. Técnicas para el estudio y clasificación de los pelos. Cuadernos de Divulgación INIREB No. 32, INIREB, Xalapa, Veracruz. 21 pp. [ Links ]
Cagle, F.R. 1949. Notes on the Raccoon, Procyon lotor megalodous, Lowery. J. Mammal. 30:45-47. [ Links ]
Ceballos, M. & C. Galindo. 1984. Mamíferos silvestres de la Cuenca de México. Ed. Limusa, México. 229 pp. [ Links ]
Ceballos, M. & A. Miranda. 1986. Los mamíferos de Chamela, Jalisco. Ed. Limusa-Instituto de Biología, UNAM, México. 436 pp. [ Links ]
Dearborne, N. 1932. Foods on some predatory fur-bearing animal in Michigan. Bull. Sch. For. Conserv. Univ. Michigan I:1-52. [ Links ]
Ewer, R. F. 1973. The Carnivores. Cornell University Press, USA. 494 pp. [ Links ]
Gander, F. F. 1966. Raccoon. Animal Kingd. 69:84-89. [ Links ]
Guerrero, S., M.R. Sandoval-Sandoval, S.S. Zalapa & A. Ordorica-Hermosillo. 1995. Hábitos de alimentación, preferencia de hábitat y densidad relativa del mapache (Procyon lotor hernandezii) en una zona de la costa sur de Jalisco (Resumen). Memorias del XIII Congreso Nacional de Zoología. Morelia, Mich. [ Links ]
Hamilton, W.J. 1951. Warm-weather food of the Raccoon in New York State. J. Mammal. 32:341-344. [ Links ]
Leopold, A. S. 1977. Fauna silvestre de México. Instituto Mexicano de Recursos Naturales Renovables. Pax-México, México. 608 pp. [ Links ]
López-Acosta, M.A. & S. Guerrero. 1994. Notas sobre mamíferos de la costa sur de Jalisco, México. BIOTAM 6(1):29-38. [ Links ]
MacCraken, J.G. & D.W. Uresk. 1984. Coyote foods in the Black Hills, South Dakota. J. Wildl. Manage. 48(4):1420-1423. [ Links ]
Magurran, A.E. 1989. Diversidad ecológica y su medición. Ed. Vedra, España. 197 pp. [ Links ]
Ordorica-Hermosillo, A.R. 1996. Estimación de la densidad relativa de carnívoros (Mammalia: Carnivora) en Tenacatita, Jalisco. Tesis Profesional, División de Ciencias Biológicas y Ambientales, Universidad de Guadalajara. [ Links ]
Schoonover, L.J., & W.H. Marshall. 1951. Food habits of Raccoon (Procyon lotor hirtus) in North-Central Minnesota. J. Mammal. 32:422-428. [ Links ]
Servín, J. & C. Huxley. 1991. La dieta del coyote en un bosque de encino-pino de la Sierra Madre Occidental de Durango, México. Acta Zool. Mex (n.s.) 44:1-26. [ Links ]
Stuewer, F.W. 1943. Raccoons: Their habits and management in Michigan. Ecol. Monogr. 13:203-257. [ Links ]
Rzedowsky, J. 1978. Vegetación de México. 3a. reimpresión, Ed. Limusa, México. 432 pp. [ Links ]
Tester, J.R. 1953. Fall foods of the Raccoon on the South Plate Valley of Northeastern Colorado. J. Mammal. 34:500-502. [ Links ]
Wood, J.E. 1954. Food habits of furbearers of the Uplands Post Oak Region in Texas. J. Mammal. 35:406-414. [ Links ]
Zalapa, S., S. Guerrero, M.H. Badii & R. Sandoval. 1998. Preferencia de hábitat, amplitud y traslape de nicho de sitio en cinco especies de carnívoros (Mammalia:Carnivora) en la costa sur de Jalisco, México. BIOTAM, 9(2 y 3):33-46. [ Links ]
Zar, J.H. 1996. Biostatistical Analysis. Prentice Hall, USA. 662 pp. [ Links ]

