










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkSalud Pública de México
versión impresa ISSN 0036-3634
Salud pública Méx vol.52 no.1 Cuernavaca ene./feb. 2010
ARTÍCULO DE REVISIÓN
Aspectos biológicos, clínicos y epidemiológicos de la tuberculosis latente
Biological, clinical and epidemiological aspects of latent tuberculosis
Jorge A Barrios-Payán, MSI,II; Mauricio Castañón-Arreola, PhDII; Mario A Flores-Valdez, PhDIII; Rogelio Hernández-Pando, PhDI
IDepartamento de Patología, sección de Patología Experimental, Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán. México DF, México
IIPosgrado en Ciencias Genómicas, Universidad Autónoma de la Ciudad de México. México DF, México
IIIUnidad de Biotecnología, Centro de Investigación y Asistencia en Tecnología y Diseño del Estado de Jalisco AC. Guadalajara, Jalisco, México
RESUMEN
Mycobacterium tuberculosis afecta a la humanidad desde hace más de 20 000 años. Su morbimortalidad es elevada, por lo que repercute económicamente en los países en desarrollo. La infección latente, caracterizada por la presencia de bacilos vivos en tejidos del huésped, con ausencia de signos y síntomas clínicos, es una característica de esta enfermedad, ya que la micobacteria puede adaptar su metabolismo para mantenerse viva con baja o nula replicación, dificultando su eliminación de los tejidos por los fármacos antituberculosos y permaneciendo inadvertida al reconocimiento y eliminación por el sistema inmunológico. Varias son las interrogantes de esta forma de tuberculosis (TB): la falta de conocimiento del metabolismo del bacilo en estado durmiente, su relación con la inmunidad del hospedero y la identificación de antígenos como marcadores diagnósticos de infección subclínica durante la latencia. Este artículo resume los aspectos biológicos, clínicos y epidemiológicos más importantes de esta forma de tuberculosis.
Palabras clave: Mycobacterium tuberculosis, latencia, tuberculosis, diagnóstico
ABSTRACT
Mycobacterium tuberculosis, the causal agent of tuberculosis, has affected humankind for approximately 20 000 years. Tuberculosis is a devastating disease, particularly in developing countries. One of its most notable characteristics is latent infection, in which live bacilli persist in the host tissues without clinical manifestations. Thus, the tuberculous bacilli adapt their metabolism to remain viable with low or no replication, avoiding their elimination by the immune system or conventional chemotherapy. Among the several problems that are particularly important to the understanding of this form of tuberculosis, and are not well-known, are the key metabolic steps that allow mycobacteria to remain in a dormant state and its interaction with host immunity. This article reviews some of the most significant biological, clinical and epidemiological aspects of this form of tuberculosis.
Key words: Mycobacterium tuberculosis, latency, tuberculosis, diagnostic
La tuberculosis (TB) es la enfermedad infecciosa que más ha afectado a la humanidad a lo largo de su historia, siendo aun hoy en día una importante causa de enfermedad y muerte, sobre todo en los estratos más pobres de la sociedad.1
La TB se manifiesta en dos formas principales: enfermedad activa e infección latente (TBL). En esta última no hay sintomatología clínica ni es transmisible, debido a que la respuesta inmune es capaz de contener el crecimiento del patógeno pero no de eliminarlo, de manera que la bacteria persiste en el organismo manteniendo una baja o nula actividad replicativa.2,3 Recientemente se ha demostrado la presencia de ADN de la micobacteria en tejido pulmonar macroscópicamente e histológicamente normal de individuos que murieron por causas ajenas a la TB.4 Este estudio reveló que el ADN micobacteriano que sugiere infección latente se localizaba frecuentemente en células epiteliales de pulmón (neumocitos tipo II, células endoteliales) y fibroblastos, lo cual evidencia la infección por M. tuberculosis en células no fagocíticas y sugiere otra estrategia de evasión inmunológica del bacilo tuberculoso cuya implicación en la epidemiología de la enfermedad no ha sido considerada y estudiada a fondo.5 Aunque la TB se encuentra lejos de ser la primera causa de muerte, como lo fue en Europa durante el siglo XIX, hoy en día se estima que anualmente existen 9 millones de nuevos casos de enfermedad activa a nivel mundial6 y un tercio de la humanidad mantiene un estado de infección latente,3 de los que se calcula que 10% desarrollará eventualmente TB. Con un reservorio de tal magnitud, el control de la TB parece ser una tarea difícil de cumplir (cuadro I).6

El concepto de infección latente
El término de latencia se ha utilizado desde 1927, inicialmente propuesto por Opie y Aronson, quienes reportaron la presencia de micobacterias en poco más del 10% de las lesiones tuberculosas antiguas (nódulos o granulomas fibróticos), de los cuales pudieron recuperar microorganismos vivos después de inyectar homogenizados de estos tejidos y producir la enfermedad en cobayos, una especie muy susceptible al bacilo tuberculoso. Este mismo estudio demostró al evaluar tejido pulmonar macroscópicamente normal (obtenido de sujetos que habían muerto por causas ajenas a la TB) que casi 50% de las muestras albergaban bacilos vivos en estado latente.7
' En 1938, Amberson8 definió el estado de latencia como "la presencia de cualquier lesión tuberculosa que no produce síntomas". Posteriormente Parrish y colaboradores la definieron como "la entidad clínica que ocurre después de que un individuo ha estado expuesto a M. tuberculosis, la infección se establece y se genera una respuesta inmune que controla al patógeno, el cual permanece en estado quiescente".9
El Comité del Consejo Médico para la Eliminación de la Tuberculosis de Massachussets, EUA, define a la latencia como "Un estado asintomático en personas que han sido infectadas con M. tuberculosis y no se tiene evidencia clínica o radiológica de la enfermedad activa". El organismo reside dentro del hospedero en un estado clínicamente inactivo o latente, contenido por el sistema inmune.10 A diferencia de los pacientes con TB activa, los individuos con TBL no representan un riesgo inmediato a la salud pública al no ser transmisores del agente infeccioso.9 La importancia de la TBL radica en el riesgo de reactivación de la infección, favorecida por condiciones como la coinfección con VIH y los trastornos metabólicos como diabetes, obesidad y malnutrición, que suprimen al sistema inmunológico propiciando la reactivación de la infección.
El término durmiente también ha sido utilizado para describir la TBL y hace referencia al estado metabólico del bacilo tuberculoso. Contrariamente a lo observado en las lesiones de TB activa, en las que generalmente se observan bacterias ácido alcohol resistentes (BAAR) que es posible cultivar, el estado metabólico de los bacilos observados en lesiones antiguas de sujetos curados o con historia de exposición a M. tuberculosis permanece incierto a casi un siglo de estudios y debates.5
A principios del siglo pasado, las investigaciones en necropsias demostraron la presencia de micobacterias viables en las lesiones tuberculosas de sujetos asintomáticos.7 Una característica de estos bacilos es que no son observables por microscopía convencional, lo que sugiere que las micobacterias en latencia se encuentran en un estado metabólico en el cual pierden su carácter de ácido alcohol resistencia, o se encuentran en cantidades menores a las detectables por microscopía convencional.11,12 Estos datos sugieren que las micobacterias son eliminadas de las lesiones primarias en pulmón, pero no de las lesiones secundarias que se presentan en los ganglios linfáticos, además de que pueden establecerse en lesiones secundarias nuevas o incluso en tejido aparentemente normal.5
Recientemente se demostró que individuos asintomáticos con reacción positiva a la tuberculina (TST+) que no tenían historia de vacunación con BCG, reconocieron antígenos de M. tuberculosis específicos de la fase no replicativa del bacilo13. En contraste, individuos sanos, no vacunados y TST- reconocen de manera diferencial los antígenos mencionados, indicando que M. tuberculosis posee determinantes antigénicos específicos de latencia2,14 que podrían utilizarse en el diagnóstico de la TBL.
Genómica de la infección latente
Sin lugar a dudas la expresión de genes durante la latencia es uno de los aspectos más intrigantes en la biología de M. tuberculosis. El esclarecimiento de los mecanismos moleculares que permiten la supervivencia de la micobacteria durante el estado latente, y de las señales que le indican cuándo reactivarse, son fundamentales para el desarrollo de vacunas, fármacos y estrategias que permitan el control y posterior erradicación de la TB.
En los genomas de M. tuberculosis y M. bovis se han identificado 13 factores sigma (sigA, sigB, sigC, sigD, sigE, sigF, sigG, sigH, sigI, sigJ, sigK, sigL y sigM),15 de los cuales sólo tres se han señalado con capacidad de respuesta ante cambios ambientales que probablemente ocurren durante la adaptación o mantenimiento de la infección latente, estos son: sigB que se activa en respuesta a varios estímulos ambientales, entre los que está la disminución en la tensión de oxígeno;16 sigE cuya transcripción ocurre durante el crecimiento exponencial y el inicio de la fase estacionaria, pero también se incrementa en condiciones de choque térmico y baja oxigenación17 y sigF (Rv3286c) (también conocido como mysB ó katF) que sólo se encuentra en micobacterias patógenas de crecimiento lento y su trascripción se incrementa después de la fagocitosis, la disminución en la concentración de nutrientes y en respuesta al tratamiento con antibióticos.13
Recientemente se caracterizó al gene Rv3133, ahora conocido como dosR (del inglés dormancy survival regulator), cuyo producto activa la transcripción de genes en respuesta a la reducción de la tensión de oxígeno y juega un papel crucial en la supervivencia de M. tuberculosis en un ambiente hipóxico.18,19 La micobacteria utiliza un sistema de dos componentes para poder sondear el medio ambiente en el que se encuentra y responder para asegurar su supervivencia; este sistema consiste de los sensores de histidina cinasas DosS y DosT y el regulador de respuesta DosR, el cual regula positivamente alrededor de 50 genes20 y negativamente a aproximadamente 68,21,22 aunque no se ha demostrado cuál es la participación precisa de las proteínas codificadas por estos genes durante la latencia.22
Recientemente se propuso el uso de la técnica de cultivo Chemostat23 en la que las condiciones de cultivo se mantienen constantes y son cuidadosamente controladas, permitiendo realizar estudios de genómica funcional para identificar la regulación genética asociada con el crecimiento del bacilo dependiente de la disponibilidad de carbono en el medio de cultivo.
Modelos animales de infección latente
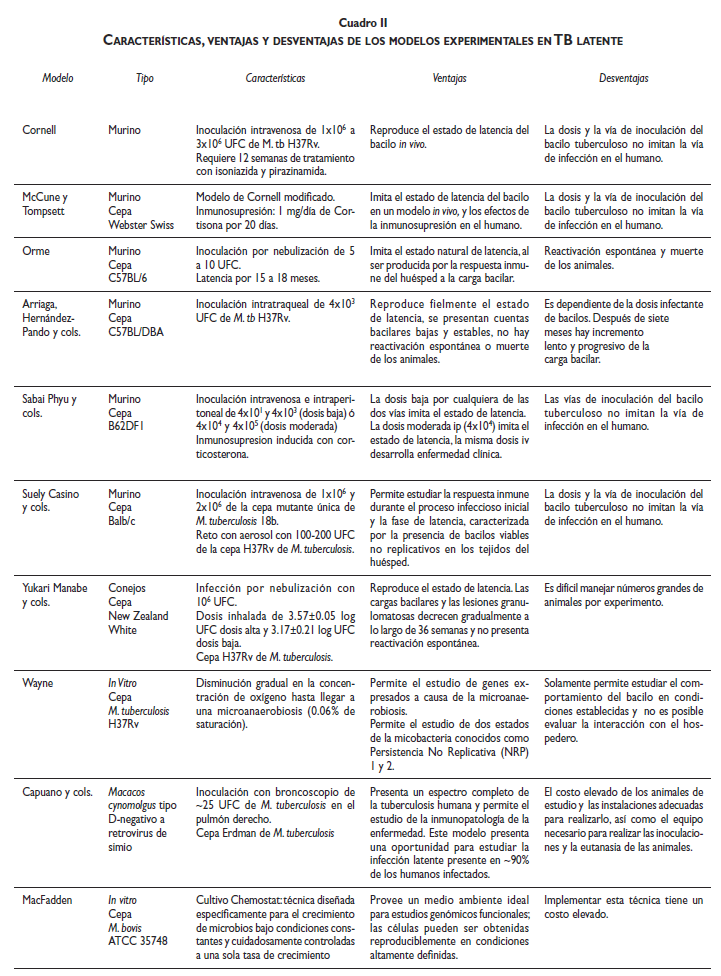
La TBL se presenta en seres humanos, algunos primates, bovinos y posiblemente otros animales de fauna silvestre, aunque en estos últimos ha sido poco estudiado. Ante este panorama las disyuntivas éticas y las limitantes económicas han impedido investigar los mecanismos que operan en el huésped durante el inicio, mantenimiento y salida del estado quiescente, salvo algunos intentos in vitro donde se simulan condiciones que podría encontrar M. tuberculosis durante esta fase. Por ende, para tratar de dilucidar el estado metabólico del bacilo durante la TBL, sus ciclos replicativos, su reactivación y la manera en como evade la respuesta inmune del huésped, se han desarrollado diversos modelos animales que intentan reproducir las condiciones de la TBL (cuadro II).24 En general, en los modelos experimentales de latencia se trata de establecer y mantener un número bajo y estable de micobacterias en los órganos de los animales de experimentación, sin signos de enfermedad hasta el momento que se induzca reactivación, generalmente como consecuencia de debilitar su respuesta inmunológica inducida sobre todo con fármacos (glucocorticoides, inhibidores del TNFα o de la enzima oxido nítrico sintetasa).
El modelo de Cornell25,26 fue el primer acercamiento experimental encaminado a demostrar la existencia de bacilos en latencia. En este modelo, se inoculan ratones por vía intravenosa (iv) con dosis que van de 1x106 a 3x106 unidades formadoras de colonias (UFC) de M. tuberculosis cepaH37Rv y, 20 minutos después, se inicia tratamiento por 12 semanas con isoniazida y pirazinamida. Noventa días después de terminado el tratamiento se pueden recuperar micobacterias del bazo, hígado y pulmón en un tercio de los ratones, y al cabo de 12 semanas se obtienen bacilos cultivables en dos tercios de los ratones infectados; este porcentaje se incrementa al 100% cuando se suprime la respuesta inmune.27
Orme y cols. desarrollaron un modelo en el que los ratones son infectados con aerosoles utilizando dosis de 5-10 UFC de M. tuberculosis /ratón. Después de 3 meses de infección la carga bacilar en pulmón es de entre 3 a 4 log10 y se mantiene así por 15 a 18 meses, después de lo cual se reactiva.28 Arriaga y cols.29 establecieron un modelo experimental utilizando ratones híbridos F1 C57BL/DBA, en el cual la inoculación de una dosis baja (4x103 UFC) de M. tuberculosis H37Rv por vía intratraqueal permite el establecimiento de una infección persistente, que semeja en algunas aspectos a la TBL. Histológicamente se observa la formación de granulomas y pequeños focos de alveolitis, con una elevada expresión de TNF-α, óxido nítrico sintetasa inducible (iNOS), IL-2 e IFN-γ, que se considera el tipo de respuesta inmunológica que induce y mantiene a la TBL. Sabai Phyu y cols.24 propusieron otro modelo utilizando ratones B6D2F1 Bom (C57BL/6JBom x DBA/2JBom); estos ratones se inoculan por vía iv o intraperitoneal (ip) con dosis bajas (4x101 y 4x103) o moderadas (4x104- 4x105), que son controladas por el sistema inmunológico, y los ratones permanecen clínicamente sanos hasta que son inmunosuprimidos con corticosterona. Yukari Manabe y cols.30 propusieron un modelo infectando conejos por aerosol con dosis de 3.17±0.21 log UFC a 3.57 ± 0.05 log UFC de M. tuberculosis H37Rv. En este modelo, a partir de la sexta semana de infección el número de bacilos cultivables y las lesiones granulomatosas decrecen gradualmente a lo largo de 36 semanas, sin reactivación espontánea de la enfermedad.
Suely Kashino y cols.31 propusieron el uso de una cepa mutante denominada 18b, con la cual los ratones infectados vía iv desarrollan un proceso infeccioso que semeja la TBL, caracterizada por la inducción de una potente respuesta de células T que mantiene niveles bajos de bacilos no replicables en los tejidos de los animales por largos periodos de tiempo, además de que los animales desarrollan resistencia a la reinfección exógena con M. tuberculosis H37Rv.
Capuano y cols.32 desarrollaron un modelo que imita la infección latente por tuberculosis en humanos; utilizaron macacos cynomolgus (un huésped muy similar al humano), los cuales fueron infectados con aproximadamente 25 UFC de la cepa virulenta Erdman de M. tuberculosis. En este modelo 50-60% de los animales inoculados desarrolla enfermedad activa y 40% restante no presenta manifestaciones de TB, muy similar a la infección latente que presentan los seres humanos.
Diagnóstico de la TBL
Históricamente, la prueba más utilizada para el diagnóstico de la TBL es la prueba cutánea de hipersensibilidad retardada al Derivado Proteico Purificado (PPD, por sus siglas en inglés).33 Esta prueba, también conocida como de la tuberculina (TST por sus siglas en inglés), es el principal método utilizado en México para identificar la exposición a M. tuberculosis. La prueba no permite diferenciar entre infección, enfermedad y sensibilización con micobacterias no tuberculosas (MNTs) debido a que el PPD es una mezcla cruda de antígenos, muchos de los cuales son conservados y forman parte de la estructura antigénica de M. tuberculosis, M. bovis, BCG y diversas MNTs,34 no obstante en muchas regiones del mundo es el único método disponible para el diagnóstico de TBL.34-37
Los avances en el campo de la genómica y las técnicas de inmunodetección han permitido el desarrollo de sistemas de diagnóstico basados en la producción de IFN-γ por células T estimuladas con ESAT-6 y CFP-10, dos antígenos codificados en la región de diferencia 1 (RD1) que se perdió en la vacuna BCG durante su atenuación y se encuentran casi exclusivamente en el complejo M. tuberculosis.35,38
Los ensayos de liberación de interferón gamma (IGRAs por sus siglas en inglés interferon-γ-release assays) han surgido como una alternativa para la detección de TB. Actualmente existen versiones comerciales como el QuantiFERON-TB Gold (Cellestis, Carnegie, Australia) y el T-SPOT.TB (Oxford Immunotec, Oxford, United Kingdom), que consisten en la cuantificación del IFN-γ producido por leucocitos de sangre total estimulados con los antígenos PPD, ESAT-6, CFP-10 y TB 7.7 en el kit QuantiFERON-IT de tercera generación.10,39 Estas pruebas de diagnóstico tienen mayor especificidad, especialmente en población vacunada con BCG.40 Diversos estudios han demostrado que el QuantiFERON-TB no presenta interferencia por la vacunación con BCG y puede diferenciar la infección con M. tuberculosis de la sensibilización con MNTs (cuadro III).41,42

El ELISPOT ha demostrado tener una sensibilidad de entre el 92 y 96% en pacientes con TB confirmada por cultivo, incluyendo pacientes VIH+.15 Al comparar la sensibilidad y especificidad de la prueba en contactos de pacientes con TB, se encontró que 53% de los contactos con TST+ fueron negativos por ELISPOT y 7% de los positivos por ELISPOT fueron TST-, lo que sugiere que muchos de ellos fueron vacunados con BCG o estuvieron expuestos a MNTs.35 En un estudio realizado en Zambia se demostró por ELISPOT que la prevalencia de TBL era de 69% (37/54) en sujetos VIH-, mientras que en los VIH+ era de 90% (35/39),43 lo que demuestra la mayor especificidad del ELISPOT en comparación con el TST.35,43 No obstante, aun se ha cuestionado el uso de ESAT-6 y CFP-10 como marcadores de latencia, toda vez que estas proteínas también se expresan regularmente durante la fase activa de crecimiento microbiano, por lo que en el mejor de los casos la reactividad hacia estas proteínas constituye de momento un indicador de infección tuberculosa.44
Establecimiento de la TBL en humanos
Después de la primo-infección, en más de 90% de los casos se controla la infección en sujetos inmunocompetentes, sin embargo, la eliminación del patógeno es lenta y difícil de llevar a cabo, por lo que se estima que en 30% de los sujetos sin tratamiento se establece TBL,5 pudiendo reactivarse y producir enfermedad progresiva después de años o incluso, décadas. Estudios epidemiológico-moleculares utilizando RFLP-IS6110 sugieren que en 40% de los casos la infección primaria nunca es contenida adecuadamente.9 En personas con TBL sin tratamiento, se calcula un riesgo acumulable de desarrollar TB del 10% durante el tiempo de vida, con un riesgo acumulado de 50-80% de que ocurra durante los primeros dos años seguidos a la infección.45,46 En pacientes VIH+ se incrementa el riesgo anual de reactivación de la TBL en 5-10%.10
Además de la coinfección con el VIH, la diabetes mellitus (DM), que se estima afecta 180 millones de personas en el mundo, es un factor que incrementa el riesgo de desarrollar TB o reactivación. En un metaanálisis realizado recientemente se determinó que el riesgo de desarrollar TB es tres veces mayor en sujetos con DM.47 En México, en un estudio prospectivo realizado en 12 municipios de la jurisdicción sanitaria de Orizaba en el estado de Veracruz, se encontró que la incidencia de TB es siete veces mayor en los sujetos que padecen DM, lo cual podría deberse a una reactivación de TBL o por infección transmitida recientemente.48
Tratamiento antituberculoso e infección latente
Los antibióticos para tratar la TB eliminan eficientemente a los bacilos que se multiplican activamente, sin embargo, los bacilos con baja actividad metabólica y replicativa, como sucede con los que están en estado quiescente, son muy resistentes a la quimioterapia; es por esto que el tratamiento convencional de la tuberculosis activa incluye cuatro fármacos (isoniazida, rifampicina, pirazinamida, etambutol) que deben ser administrados al menos por 6 meses.9 Estos fármacos constituyen la fase inicial del tratamiento; su uso por dos meses permite la rápida eliminación de bacilos con replicación activa, promoviendo la negativización del esputo. La segunda fase del tratamiento usualmente consiste en la administración de isoniazida y rifampicina por un periodo de 4 ó 7 meses; esta etapa del tratamiento busca eliminar los bacilos remanentes en estado durmiente o con baja actividad replicativa.
La isoniazida requiere ser metabolizada por la catalasa-peroxidasa bacteriana para inhibir la biosíntesis de ácidos micólicos, los cuales son componentes esenciales de la pared micobacteriana. La isoniazida tiene efecto bacteriostático contra bacilos quiescentes.49 Este fármaco es utilizado para dar tratamiento profiláctico durante 6 ó 12 meses, lo cual reduce la tasa de reactivación de la TB en un 65 y 90% respectivamente.50,51 La rifampicina ha mostrado poca actividad contra bacilos quiescentes, ya que inhibe la trascripción al interactuar con la subunidad beta de la RNA polimerasa. Por otro lado, la pirazinamida requiere ser convertida a ácido pirazinoico para ser activo. Su actividad bactericida parece ser específica contra M. tuberculosis;es activa contra bacilos persistentes y bacilos que no se están dividiendo, incluso aquellos que se localizan intracelularmente. El etambutol es un fármaco que afecta la biosíntesis de la pared celular y contribuye a incrementar la susceptibilidad de M. tuberculosis a otros fármacos.25-27,52, 53
El esquema de tratamiento para la TBL generalmente consiste en la administración de monoterapias. Recientemente se encontró que la terapia con rifampicina por cuatro meses produce menos eventos adversos que la terapia de nueve meses con isoniazida, observándose un menor número de casos de hepatitis y mayor apego al tratamiento.54 Las investigaciones enfocadas a encontrar fármacos más eficaces para el tratamiento de la TBL generalmente se basan en evaluaciones de cultivos in vitro en condiciones anaerobias. Heifets y cols.55 evaluaron la actividad antituberculosa de diversos fármacos y demostraron que la capreomicina tenía actividad bactericida contra bacilos en un estado no replicativo, similar al obtenido con metronidazol. No obstante, es necesario realizar más estudios en los modelos animales apropiados para probar la efectividad de este fármaco contra bacilos quiescentes.
El metronidazol, que no tiene efecto bactericida contra bacilos en crecimiento exponencial, es bactericida para bacterias en fase estacionaria y en estado persistente no replicativo. Este fármaco se ha evaluado de manera individual y en combinación con otros fármacos antituberculosos, sin embargo, se observó que el tratamiento con metronidazol (por seis semanas a 100mg/kg de peso), no tuvo efecto en la reducción de la carga bacilar en los granulomas pulmonares de cobayos infectados y contribuyó significativamente a empeorar la lesión inflamatoria en los pulmones cuando se combinó con isoniacida, rifampizina y pirazinamida.56
Discusión
La tuberculosis latente ocurre después de que se genera una respuesta inmunológica contra el bacilo durante la infección primaria, ante lo cual el bacilo entra en un estado quiescente en el que la bacteria puede persistir en los tejidos sin ser destruida por la respuesta inmunológica del hospedero, y en un nicho celular que le puede proporcionar una barrera muy efectiva para protegerse de la acción bactericida de los fármacos.
En general, se asume que un porcentaje significativo de los casos de TB activa es el resultado de la reactivación de una infección latente. Este fenómeno es particularmente frecuente en sujetos que presentan coinfección con el VIH. Según la OMS se estima que una tercera parte de la población tiene TBL, sin embargo no se sabe cuántos de los sujetos que han estado expuestos a M. tuberculosis eliminan totalmente la infección. Estudios moleculares han demostrado la presencia de ADN micobacteriano en tejido pulmonar, tejido adiposo,4,57 bazo, hígado, intestino, riñón y cerebro macroscópica e histológicamente normal de sujetos que murieron por causas ajenas a tuberculosis.* Lo anterior muestra los diversos nichos celulares que puede tener el bacilo tuberculoso además del pulmón, lo cual demanda el desarrollo de alternativas terapéuticas específicas para eliminar bacterias en estado quiescente o para evitar su reactivación.
El uso de monoterapia como medida profiláctica para eliminar la infección latente es una estrategia riesgosa en el control de la tuberculosis, ya que el costo de tratar personas de las que no se sabe si presentarán reactivación de la infección es mayor al beneficio que se podría obtener, y hasta el momento no se conocen fármacos eficientes en el tratamiento de la TBL; además, el uso de un solo antibiótico facilita el desarrollo de cepas drogoresistentes. Es por esto que se ha propuesto el uso de vacunas terapéuticas que refuercen la inmunidad protectora y, en consecuencia, eviten la reactivación.58,59
Conclusiones
La latencia es uno de los principales obstáculos para erradicar la TB, por lo tanto, el entendimiento de los mecanismos inmunológicos y genéticos que operan durante la persistencia y la reactivación de este estado infeccioso nos ayudará a crear medidas más efectivas para controlar y erradicar a este patógeno. Es importante desarrollar terapias eficaces, que sean de corta duración, y que carezcan de efectos colaterales y toxicidad, tanto en sujetos inmunocompetentes como inmunosuprimidos, con un costo accesible y, sobre todo, que estén al alcance de personas que viven en países en vías de desarrollo con zonas endémicas de tuberculosis. El desarrollo de técnicas de laboratorio que ayuden a diferenciar acertadamente la tuberculosis activa de la latente y el desarrollo de biomarcadores que permitan determinar la eficacia del tratamiento será también de gran ayuda para controlar esta importante enfermedad infecciosa.
Referencias
1. Daniel TM. The origins and precolonial epidemiology of tuberculosis in the Americas: can we figure them out? Int J Tuberc Lung Dis 2000;4:395-400. [ Links ]
2. Leyten EM, Lin MY, Franken KL, Friggen AH, Prins C, van Meijgaarden KE, et al. Human T-cell responses to 25 novel antigens encoded by genes of the dormancy regulon of Mycobacterium tuberculosis. Microbes Infect 2006;8:2052-2060. [ Links ]
3. Cardona PJ, Ruiz-Manzano J. On the nature of Mycobacterium tuberculosis-latent bacilli. Eur Respir J 2004;24:1044-1051. [ Links ]
4. Hernandez-Pando R, Jeyanathan M, Mengistu G, Aguilar D, Orozco H, Harboe M, et al. Persistence of DNA from Mycobacterium tuberculosis in superficially normal lung tissue during latent infection. Lancet 2000;356:2133-2138. [ Links ]
5. Gomez JE, McKinney JD. M. tuberculosis persistence, latency, and drug tolerance. Tuberculosis (Edinb) 2004;84:29-44. [ Links ]
6. World Health Organization. Global tuberculosis control: Surveillance, planning, financing.Genova: WHO, 2004. [ Links ]
7. Opie E, Aronson J. Tubercle bacilli in latent tuberculous lesions and in lung tissue without tuberculous lesions. Arch Pathol Lab Med 1927;4:21. [ Links ]
8. Amberson J. The significance of latent forms of tuberculosis. N Engl J Med 1938;219:5. [ Links ]
9. Parrish NM, Dick JD, Bishai WR. Mechanisms of latency in Mycobacterium tuberculosis. Trends Microbiol 1998;6:107-112. [ Links ]
10. Tuberculosis, TMACftEo. Latent tuberculosis infection: A guide for Massachusetts providers. Massachusetts, USA: The Medical Advisory Committee for the Elimination of Tuberculosis, 2005. [ Links ]
11. Khomenko AG. The variability of Mycobacterium tuberculosis in patients with cavitary pulmonary tuberculosis in the course of chemotherapy. Tubercle 1987;68:243-253. [ Links ]
12. Stanford JL. Much's granules revisited. Tubercle 1987;68:241-242. [ Links ]
13. Sherman DR, Voskuil M, Schnappinger D, Liao R, Harrell MI, Schoolnik GK. Regulation of the Mycobacterium tuberculosis hypoxic response gene encoding alpha -crystallin. Proc Natl Acad Sci U S A 2001;98:7534-7539. [ Links ]
14. Demissie A, Leyten EM, Abebe M, Wassie L, Aseffa A, Abate G, et al. Recognition of stage-specific mycobacterial antigens differentiates between acute and latent infections with Mycobacterium tuberculosis. Clin Vaccine Immunol 2006;13:179-186. [ Links ]
15. Richeldi L, Ewer K, Losi M, Hansell DM, Roversi P, Fabbri LM, et al. Early diagnosis of subclinical multidrug-resistant tuberculosis. Ann Intern Med 2004;140:709-713. [ Links ]
16. Cole ST, Brosch R, Parkhill J, Garnier T, Churcher C, Harris D, et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 1998;393:537-544. [ Links ]
17. Wayne LG, Hayes LG. An in vitro model for sequential study of shiftdown of Mycobacterium tuberculosis through two stages of nonreplicating persistence. Infect Immun 1996;64:2062-2069. [ Links ]
18. Boon C, Dick T. Mycobacterium bovis BCG Response Regulator Essential for Hypoxic Dormancy. J Bacteriol 2002;184:6760-6767. [ Links ]
19. Park HD, Guinn KM, Harrell MI, Liao R, Voskuil MI, Tompa M, et al. Rv3133c/dosR is a transcription factor that mediates the hypoxic response of Mycobacterium tuberculosis. Mol Microbiol 2003;48:833-843. [ Links ]
20. Rustad TR, Harrell MI, Liao R, Sherman DR. The enduring hypoxic response of Mycobacterium tuberculosis. PLoS ONE 2008;3:e1502. [ Links ]
21. Kumar A, Toledo JC, Patel RP, Lancaster JR, Steyn AJC. Mycobacterium tuberculosis DosS is a redox sensor and DosT is a hypoxia sensor. Proc Natl Acad Sci 2007;104:11568-11573. [ Links ]
22. Roupie V, Romano M, Zhang L, Korf H, Lin MY, Franken KLMC, et al. Immunogenicity of eight dormancy regulon-encoded proteins of Mycobacterium tuberculosis in DNA-vaccinated and tuberculosis-infected mice. Infect Immun 2007;75:941-949. [ Links ]
23. Beste DJV, Laing E, Bonde B, Avignone-Rossa C, Bushell ME, McFadden JJ. Transcriptomic analysis identifies growth rate modulation as a component of the Adaptation of Mycobacteria to survival inside the macrophage. J Bacteriol 2007;189:3969-3976. [ Links ]
24. Phyu S, Mustafa T, Hofstad T, Nilsen R, Fosse R, Bjune G. A mouse model for latent tuberculosis. Scand J Infect Dis 1998;30:59-68. [ Links ]
25. McCune RM Jr, Tompsett R. Fate of Mycobacterium tuberculosis in mouse tissues as determined by the microbial enumeration technique. I. The persistence of drug-susceptible tubercle bacilli in the tissues despite prolonged antimicrobial therapy. J Exp Med 1956;104:737-762. [ Links ]
26. McCune RM Jr, McDermott W, Tompsett R. The fate of Mycobacterium tuberculosis in mouse tissues as determined by the microbial enumeration technique. II. The conversion of tuberculous infection to the latent state by the administration of pyrazinamide and a companion drug. J Exp Med 1956;104:763-802. [ Links ]
27. McCune RM, Feldmann FM, McDermott W. Microbial persistence. II. Characteristics of the sterile state of tubercle bacilli. J Exp Med 1966;123:469-486. [ Links ]
28. Orme IM. A mouse model of the recrudescence of latent tuberculosis in the elderly. Am Rev Respir Dis 1988;137:716-718. [ Links ]
29. Arriaga AK, Orozco EH, Aguilar LD, Rook GA, Hernandez-Pando R. Immunological and pathological comparative analysis between experimental latent tuberculous infection and progressive pulmonary tuberculosis. Clin Exp Immunol 2002;128:229-237. [ Links ]
30. Manabe YC, Kesavan AK, Lopez-Molina J, Hatem CL, Brooks M, Fujiwara R, et al. The aerosol rabbit model of TB latency, reactivation and immune reconstitution inflammatory syndrome. Tuberculosis (Edinb) 2008;88:187-196. [ Links ]
31. Kashino SS, Ovendale P, Izzo A, Campos-Neto A. Unique model of dormant infection for tuberculosis vaccine development. Clin Vaccine Immunol 2006;13:1014-1021. [ Links ]
32. Capuano SV III, Croix DA, Pawar S, Zinovik A, Myers A, Lin PL, et al. Experimental Mycobacterium tuberculosis infection of Cynomolgus macaques closely resembles the various manifestations of human M. tuberculosis infection. Infect Immun 2003;71:5831-5844. [ Links ]
33. Society AT. Targeted tuberculin testing and treatment of latent tuberculosis infection. This official statement of the American Thoracic Society was adopted by the ATS Board of Directors, July 1999. This is a Joint Statement of the American Thoracic Society (ATS) and the Centers for Disease Control and Prevention (CDC). This statement was endorsed by the Council of the Infectious Diseases Society of America. (IDSA), September 1999, and the sections of this statement. Am J Respir Crit Care Med 2000;161:S221-S247. [ Links ]
34. Huebner RE, Schein MF, Bass JB Jr. The tuberculin skin test. Clin Infect Dis 1993;17:968-975. [ Links ]
35. Codecasa L, Mantegani P, Galli L, Lazzarin A, Scarpellini P, Fortis C. An in-house RD1-based enzyme-linked immunospot-gamma interferon assay instead of the tuberculin skin test for diagnosis of latent Mycobacterium tuberculosis infection. J Clin Microbiol 2006;44:1944-1950. [ Links ]
36. Fine PE, Bruce J, Ponnighaus JM, Nkhosa P, Harawa A, Vynnycky E. Tuberculin sensitivity: conversions and reversions in a rural African population. Int J Tuberc Lung Dis 1999;3:962-975. [ Links ]
37. Diagnostic standards and classification of tuberculosis in adults and Children. This official statement of the American Thoracic Society and the Centers for Disease Control and Prevention was adopted by the ATS Board of Directors, July 1999. This statement was endorsed by the Council of the Infectious Disease Society of America, September 1999. Am J Respir Crit Care Med 2000;161:1376-1395. [ Links ]
38. Goletti D, Vincenti D, Carrara S, Butera O, Bizzoni F, Bernardini G, et al. Selected RD1 peptides for active tuberculosis diagnosis: comparison of a gamma interferon whole-blood enzyme-linked immunosorbent assay and an enzyme-linked immunospot assay. Clin Diagn Lab Immunol 2005;12:1311-1316. [ Links ]
39. Connell TG, Ritz N, Paxton GA, Buttery JP, Curtis N, Ranganathan SC. A three-way comparison of Tuberculin skin testing, QuantiFERON-TB Gold and T-SPOT.TB in Children. PLoS ONE 2008;3:e2624. [ Links ]
40. Pai M, Zwerling A, Menzies-D. Systematic review: T-cell-based assays for the diagnosis of latent tuberculosis infection: an update. Ann Intern Med 2008;149:177-184. [ Links ]
41. Pottumarthy S, Morris AJ, Harrison AC, Wells VC. Evaluation of the Tuberculin Gamma Interferon Assay: potential to replace the Mantoux Skin Test. J Clin Microbiol 1999;37:3229-3232. [ Links ]
42. van Pinxteren LAH, Ravn P, Agger EM, Pollock J, Andersen P. Diagnosis of Tuberculosis based on the two specific antigens ESAT-6 and CFP10. Clin Diagn Lab Immunol 2000;7:155-160. [ Links ]
43. Chapman AL, Munkanta M, Wilkinson KA, Pathan AA, Ewer K, Ayles H, et al. Rapid detection of active and latent tuberculosis infection in HIV-positive individuals by enumeration of Mycobacterium tuberculosis-specific T cells. Aids 2002;16:2285-2293. [ Links ]
44. Shi L, North R, Gennaro ML. Effect of growth state on transcription levels of genes encoding major secreted antigens of Mycobacterium tuberculosis in the mouse lung. Infect Immun 2004;72:2420-2424. [ Links ]
45. Ferebee SH. Controlled chemoprophylaxis trials in tuberculosis. A general review. Bibl Tuberc 1970;26:28-106. [ Links ]
46. Styblo K, Meijer J, Sutherland I. [The transmission of tubercle bacilli: its trend in a human population]. Bull World Health Organ 1969;41:137-178. [ Links ]
47. Jeon CY, Murray MB. Diabetes mellitus increases the risk of active Tuberculosis: a systematic review of 13 observational studies. PLoS Med 2008;5:e152. [ Links ]
48. Ponce-de-Leon A, Garcia-Garcia MdL, Garcia-Sancho MC, Gomez-Perez FJ, Valdespino-Gomez JL, Olaiz-Fernandez G, et al. Tuberculosis and diabetes in Southern Mexico. Diabetes Care 2004;27:1584-1590. [ Links ]
49. Palomino JC, Leão SC, Ritacco V. Tuberculosis 2007 from basic science to patient care. Alemania: Amedeo Challenge, 2007. [ Links ]
50. Comstock GW, Baum C, Snider DE Jr. Isoniazid prophylaxis among Alaskan Eskimos: a final report of the bethel isoniazid studies. Am Rev Respir Dis 1979;119:827-830. [ Links ]
51. Comstock GW, Woolpert SF. Preventive treatment of untreated, nonactive tuberculosis in an Eskimo population. Arch Environ Health 1972;25:333-337. [ Links ]
52. Hobby GL, Lenert TF. The in vitro action of antituberculous agents against multiplying and non-multiplying microbial cells. Am Rev Tuberc 1957;76:1031-1048. [ Links ]
53. McCune RM, Feldmann FM, Lambert HP, McDermott W. Microbial persistence. I. The capacity of tubercle bacilli to survive sterilization in mouse tissues. J Exp Med 1966;123:445-468. [ Links ]
54. Menzies D, Long R, Trajman A, Dion MJ, Yang J, Al Jahdali H, et al. Adverse events with 4 months of Rifampin therapy or 9 months of Isoniazid therapy for latent Tuberculosis infection: a randomized trial. Ann Intern Med 2008;149:689-697. [ Links ]
55. Heifets L, Simon J, Pham V. Capreomycin is active against non-replicating M. tuberculosis. Ann Clin Microbiol Antimicrob 2005;4:6. [ Links ]
56. Hoff DR, Caraway ML, Brooks EJ, Driver ER, Ryan GJ, Peloquin CA, et al. Metronidazole lacks antibacterial activity in guinea pigs infected with Mycobacterium tuberculosis. Antimicrob Agents Chemother 2008;52: 4137-4140. [ Links ]
57. Neyrolles O, Hernandez-Pando R, Pietri-Rouxel F, Fornes P, Tailleux L, Barrios-Payan JA, et al. Is adipose tissue a place for Mycobacterium tuberculosis persistence? PLoS ONE 2006;1:e43 [ Links ]
58. Campuzano J, Aguilar D, Gonzalez-y Merchant J, Salas P, Espitia C, Hernández-Pando R. The PGRS domain of Mycobacterium tuberculosis. PE-PGRS-Rv1759 is an efficient subunit vaccine to prevent reactivation in a murine model of chronic tuberculosis. Vaccine 2007; 25 (18): 3722-3729. [ Links ]
59. Rook GAW, Lowry D, Hernández-Pando R. Immunotherapeutics for tuberculosis in experimental animals: Is there a common pathway activated by effective protocols? J Infect Dis 2007; 196: 191-198 [ Links ]
 Solicitud de sobretiros:
Solicitud de sobretiros:
Dr. Rogelio Hernández Pando
Departamento de Patología, sección de Patología Experimental, Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán
Vasco de Quiroga 15, col. Sección XVI
14000, Delegación Tlalpan, Distrito Federal, México
Correo electrónico: rhdezpando@hotmail.com
Fecha de recibido: 7 de enero de 2009
Fecha de aceptado: 27 de octubre de 2009
* Barrios-Payán J, Aguilar LD, Castañón M, Hernández-Pando R. Persistance of DNA of Mycobacterium tuberculosis in superficially normal pulmonary and extrapulomonary human tissue. Manuscrito en preparación.


{kind=link}