











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink

INTRODUCCIÓN
Las infecciones respiratorias agudas (IRA) representan un importante problema de salud, ya que ocupan el segundo lugar a nivel mundial de morbimortalidad en niños menores de 5 años; también son frecuentes en personas de la tercera edad, así como en sujetos inmunocomprometidos1. Se calcula que cada año mueren alrededor del 15% de infantes menores de 5 años por neumonía. Las IRA se presentan en todo el mundo, pero con mayores índices de mortalidad en países en vías de desarrollo2, aproximadamente 20% más que en países desarrollados (3-4%)3. En México, para el 2016, el INEGI reportó que ocupan el tercer lugar de morbilidad con 21,572 muertes en la población general, pero en infantes de 1 a 4 años ocupan el tercer lugar con 500 muertes anuales4.
Los principales agentes causales de las IRA son los virus, y el agente etiológico más frecuente es el ortopneumovirus humano (hOPV), anteriormente nombrado virus sincicial respiratorio humano, seguido del metapneumovirus humano (hMPV), responsable del 5-15% de hospitalizaciones en infantes5, posteriormente respirovirus y ortorubulavirus humano (anteriormente conocidos como virus de parainfluenza humano) 6, influenza virus (IV), coronavirus y rinovirus, entre otros. En la actualidad se han reportado casos de IRA en donde más de un patógeno actúa como agente causal, lo que hace difícil su diagnóstico y aún más cuando alguno de los patógenos no ha sido descrito, como fue el caso de hMPV2,3. En el año 2001, Van den Hoogen y colaboradores reportaron al hMPV como un nuevo agente etiológico, el cual presentaba manifestaciones clínicas similares a las causadas por hOPV; ambos patógenos pueden causar infecciones respiratorias que cursan de leves a severas, con complicaciones como bronquiolitis y neumonía, lo cual hace que su diagnóstico clínico sea difícil7.
En la actualidad, en Estados Unidos las muertes y hospitalizaciones a causa de hOPV y hMPV se han estimado en pérdidas económicas de entre 365 millones y 585 millones de dólares respectivamente8, lo cual pudiera disminuir con tratamiento profiláctico, ya que los tratamientos actuales son de soporte que incluyen disminuir los síntomas, administración de oxígeno, remplazo de fluidos y, en caso de ser necesario, ventilación mecánica9. Por otro lado, los tratamientos antivirales como anticuerpos humanizados (palivizumab, entre otros) debido a su costo (aproximadamente 12,000 pesos por dosis), difícil obtención1 y efectos indeseables, son considerados solo en casos severos9.
Metaneumovirus humano: virología molecular
El hMPV es un virón pleomórfico de 150 nm a 300 nm. Es miembro del género Metapneumovirus, familia Pneumoviridae, que pertenece al orden de los Mononegavirales10 correspondiente a virus de ARN monocatenario negativo no segmentado, con una longitud de aproximadamente 13 kb, que consta de 8 genes (figura 1) en el orden 3’-N-P-M-F-M2-SH-G-L-5’ y codifican 9 proteínas ilustradas en la tabla 1. El hMPV comprende 2 subgrupos A y B, con 4 linajes genéticos (A1, A2, B1 y B2) 11.

En la superficie del virus se expresan 3 proteínas, la F (guinda), G (verde obscuro), SH (naranja). La proteína M (azul) encapsula las nucleoproteínas N (en verde claro), asociadas al genoma. El complejo ribonucleoproteína está asociado con la proteína L (amarillo), P (morado), M2-1 y M2-2 (azul cielo). Modificada de Skiadopoulos et al., 2014.
Figura 1 Diagrama de una partícula de metapneumovirus humano
Tabla 1 Proteínas de hMPV
| Nombre de la proteína | Abreviatura | Función |
| Nucleoproteína | N | Encapsida el genoma viral, por lo tanto, lo protege de las nucleasas |
| Fosfoproteína | P | Actúa como un cofactor que estabiliza la proteína L; forma parte del complejo RNP (ribonucleoproteína viral) |
| Proteína de matriz | M | Rodea el genoma del virus, permitiendo el desarrollo y la interacción del complejo RNP |
| Glicoproteína de fusión | F | Se une a receptores heparán sulfato en la superficie celular para la fusión viral |
| Factor de transcripción | M2-1 | Factor transcripcional de elongación |
| Factor regulador de RNA | M2-2 | Juega el papel de mayor virulencia ya que regula los patrones de reconocimiento de los receptores tipo toll y los inducibles por ácido retinoico |
| Glicoproteína pequeña hidrofílica | SH | Probablemente forma un canal iónico protón selectivo que permite el desarrollo de la fusión viral |
| Glicoproteína de anclaje | G | Ayuda en la fusión viral por el reconocimiento de glicosaminoglicanos |
| Polimerasa de ARN | L | Transcribe el RNA genómico, se asegura que el RNA esté protegido en ambos extremos (CAP y poliadenilación) |
Los virus pertenecientes a esta familia constan de la proteína F o glicoproteína de fusión, encargada de fusionar el virón con la membrana mediante el reconocimiento a receptores de la célula blanco y que en la actualidad se utiliza como un blanco terapéutico1; el hMPV y el hOPV comparten 37.9% de identidad en la secuencia de aminoácidos de la glicoproteína de fusión12. Un estudio realizado en la universidad de Maryland demostró que aislados clínicos de 2 de los linajes del hMPV conservan un porcentaje de identidad para la proteína M de 97%, seguido de la nucleoproteína con 96%, glicoproteína F con 95%, y la proteína SH, que es la menos conservada, con 59%10.
La proteína F se sintetiza como un precursor F0 (figura 2), que posteriormente se escinde por una proteasa, en 2 subunidades, F1 y F2, y produce un péptido de fusión hidrofóbico (FP) en el extremo N-terminal de F113. Durante el proceso de fusión FP se inserta en la membrana del hospedero5.
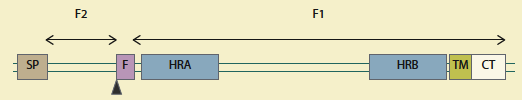
Esquema del precursor de la proteína de fusión, el triángulo negro indica el sitio de corte, F1: subunidad 1; F2: subunidad 2; SP: péptido señal; F: péptido fusión; HRA/HRB: héptada repetida A y B; TM: dominio transmembrana; CT: extremo C terminal. Modificada de Melero y Mas, 2015.
Figura 2 Esquema del precursor F0
El fragmento F1 contiene un péptido de fusión hidrofóbico N-terminal y 2 hidrofílicos que son regiones de heptarrepetidas y conservadas (HR), la héptada N-terminal (HRA), situada inmediatamente adyacente al péptido de fusión y la C-terminal (HRB), que se encuentra proximal al dominio transmembrana (TM). Al activarse la proteína F inserta su péptido de fusión en la membrana blanco y HRA y HRB se rearreglan en un haz estable de 6 hélices (6HB), y posicionan a los péptidos de fusión y los dominios TM próximos a la misma membrana blanco (figura 3); este proceso se conoce como fusogenización14.
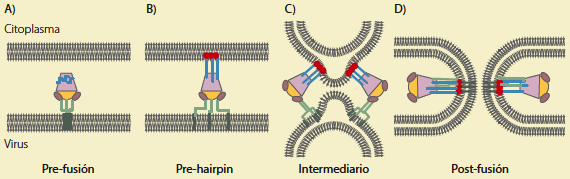
Modelo de fusión hMPV: a) La proteína F en estado prefusogénico, HRA en azul y HRB en verde, b) Reordenamiento de las héptadas repetidas e inserción del péptido de fusión en la membrana de la célula blanco, c) Intermediario de unión entre las membranas por medio de varias proteínas F, d) unión de HRA con HRB dando lugar a la formación de los 6 haces de hélices y por lo tanto la fusión virus-célula. Modificada de Melero y Mas, 2015.
Figura 3 Diagrama que representa la fusión virión-célula de hMPV
Dentro de las principales características que diferencian a la naciente familia Pneumoviridae de su antigua familia Paramyxoviridae, está que sus miembros contienen una proteína de fusión (F) que es independiente de la proteína de adhesión (G) 14 y se ha probado que, por sí sola, la proteína F es suficiente para inducir la fusión con la célula, lo cual sugiere que es una proteína con función dual: unión y fusión, independiente a los posibles receptores de la proteína G (heparán sulfato y quimiocinas).
Congruente con esta hipótesis, se ha encontrado un motivo de unión a integrinas (329RGD331) en dicha proteína y en un estudio realizado en China se demostró que los aminoácidos R329 y G330 del motivo de unión a integrinas son esenciales para la unión de la proteína a las integrinas α5β1 y αν, por lo cual una mutación en estos residuos impide la fusión del virus con la célula5, y por esta razón se puede sospechar de la intervención de esta proteína como receptor para la fusión viral, aunque faltan estudios para poder demostrarlo3.
Epidemiología
Las IRA se distribuyen a nivel mundial como se muestra en la figura 4. En el caso particular del hMPV, los aislados virales2 presentan una distribución dependiente de las estaciones; en invierno y primavera principalmente en el hemisferio norte, y en verano predomina en el hemisferio sur; según algunos estudios el mayor porcentaje de aislados se presentan después de la temporada de infecciones por hOPV3. Se reporta al hMPV como la segunda causa de IRA y responsable de infecciones menos severas que las causadas por hOPV, lo cual no le resta importancia, ya que, en lugares como el sur de Asia y sureste de África, 2 terceras partes de los pacientes mueren a causa de este patógeno. Actualmente, a nivel mundial, la tasa de hospitalizaciones asociada con infecciones de hMPV es de una por cada 1,000 niños, lo cual es bajo en comparación con hOPV, pero es la misma que para respirovirus y ortorubulavirus humano15.

Distribución geográfica mundial promedio de los linajes (A1, A2, B1 y B2) de hMPV. Su incidencia de acuerdo a la región geográfica va de enero a marzo en el hemisferio norte y de junio a julio en el hemisferio sur. Modificado de Panda et al., 2014.
Figura 4 Distribución geográfica mundial de los genotipos de hMPV
Estudios de seroprevalencia han demostrado la presencia de anticuerpos contra hMPV en muestras obtenidas en 1958, lo que sugiere que el virus ha estado en circulación por lo menos hace más de 50 años2, a pesar de que su identificación fue apenas en el 2001; además, análisis evolutivos sugieren que el hMPV provino del metapneumovirus aviar tipo C hace aproximadamente 200 años8.
En México existen pocos estudios epidemiológicos del hMPV, pero un estudio realizado en Guadalajara, Jalisco, reportó que el hMPV fue el agente etiológico más frecuente en niños menores de 5 años, con un porcentaje de 22%16; otros estudios realizados en San Luis Potosí, reportan que ocupan el segundo lugar de incidencia después del hOPV, porque se ha identificado en un porcentaje de 6 a 20% en niños menores de 3 años17.
Existen datos contradictorios respecto a las coinfecciones de hMNV en cuanto al número de virus involucrados, las asociaciones que existen entre estos y la gravedad de la enfermedad; dichas asociaciones también presentan diferentes frecuencias en las poblaciones, por ejemplo, un estudio en Rusia reportó que 14.5% de las IRA causadas por virus fueron coinfecciones, pero en ninguna de estas asociaciones participó el hMPV18. Sin embargo, en otro estudio realizado en la India, 26% de las IRA fueron causadas por coinfecciones, de las cuales el hMPV se presentó en 1.68% de los pacientes formando asociaciones con hRUV-4 y adenovirus (AdV) 19. En nuestro país, 2 estudios reportan frecuencias de coinfecciones parecidas a las ya mencionadas (13.9 y 28%), pero la participación del hMPV es más alta, con un promedio de 5.47% de estas asociaciones, que se detectaron en conjunto con IV, hOPV, hRUV, hREV, AdV y coronavirus20,21.
Factores de riesgo
La severidad de la enfermedad puede deberse a ciertos factores de riesgo, los cuales pueden estar asociados con las características del patógeno y con el estado clínico y el área demográfica donde vive el paciente. Por otro lado, la carga viral se ha asociado con la duración de la enfermedad y con la severidad de las IRA; sin embargo, no existe una correlación precisa entre el genotipo del virus y la severidad de la enfermedad, pues algunos reportes señalan que el genotipo A es más virulento que el genotipo B (2), y otros, como el de Papenburg et al. (2013), indican que es el genotipo B el que se asocia con una mayor severidad de la enfermedad22.
Los sujetos considerados de alto riesgo son los niños menores de 5 años, con mayor incidencia en infantes de 6 meses de edad9, debido a la falta de maduración de su sistema inmunológico; los niños con nacimiento prematuro, bajo peso, asma, infecciones nosocomiales preexistentes23, enfermedades crónico-pulmonares o cardiacas, o con desórdenes neuronales. Muchos estudios han reportado al hMPV en coinfección con otros patógenos como hOPV, bocavirus, rinovirus, coronavirus, Respirovirus y Ortorubulavirus humano IV A y B3. También se han encontrado aislados en personas de edad adulta que, sin haberlo diagnosticado antes, alguna vez fueron infectadas por este virus, y también en personas inmunocomprometidas2, con mayor frecuencia si han recibido trasplantes o si tienen complicaciones hematológicas (como leucemias) 3.
Manifestaciones clínicas
El diagnóstico diferencial basado exclusivamente en la clínica dificulta determinar al agente etiológico hMPV o hOPV, ya que tanto los factores de riesgo como las manifestaciones clínicas son similares y, a pesar de esto, en México el diagnóstico se hace de acuerdo con la guía práctica clínica en la cual se describe que este tipo de infecciones cursan frecuentemente con tos, sibilancias, estertores, dificultad para respirar y, en algunos casos, se produce la muerte por falla respiratoria11. En algunos países, como en algunos hospitales de México, las pruebas para la identificación del agente etiológico responsable se basan en el uso de anticuerpos específicos marcados con fluorocromos para realizar los ensayos de inmunofluorescencia directa, indirecta o el uso de ensayos moleculares como el RT-qPCR.
Las infecciones causadas por hMPV se presentan en la versión asintomática y la sintomática. Esta última generalmente presenta signos tales como: fiebre (< 38 °C), tos, hipoxia, sibilancias, rinorrea, disnea3, linfopenia con monocitosis; en adultos inmunocomprometidos son comunes los dolores de cabeza, la adenopatía cervical, la faringitis y el herpes labial recurrente11.
Patogénesis
La proteína F es la responsable de la fusión del virus con la célula hospedera, y se sabe que el mecanismo de fusión es diferente al del resto de los miembros de las familias Pneumoviridae y Paramyxoviridae, los cuales realizan este proceso a pH neutro11.
Como primeras células diana, el hMPV infecta a las células epiteliales del tracto respiratorio inferior, además de los leucocitos circulantes, como monocitos y células dendríticas, que principalmente se encuentren en los pulmones24, y provoca la fusión de la célula infectada con las células vecinas, lo que da lugar a la formación de sincicios, cuyo tamaño dependerá del linaje25. En el caso de una infección severa, se presentan cambios a nivel de pulmones, que incluyen daño en la arquitectura epitelial respiratoria, desprendimiento de células epiteliales, pérdida de los cilios, producción de moco e inflamación exacerbada de la zona intersticial del pulmón24.
La primera acción de defensa ante la infección que se presenta es la secreción de interferones (INF) del tipo I, con lo que se establece un estado antiviral en las células vecinas no infectadas, además de que se promueve la eliminación de células infectadas mediante el mejoramiento de la respuesta del antígeno. La inducción de dicho INF, no podría llevarse a cabo sin el reconocimiento de estructuras de ARN de doble cadena, evento que se presenta durante el ciclo de replicación viral (genoma-antigenoma). Esto provoca una señalización celular conocida como MAVS (señalización antiviral mitocondrial), que desencadena la expresión de receptores y secreción de citocinas11, como la Interleucina 2 (IL-2), IL-6, IL-8, IL-10, el factor de necrosis tumoral α (TNF-α), entre otros. Finalmente la infección persistente por hMPV se atribuye a una mínima respuesta inmunológica, ya que la depuración del virus mediada por los linfocitos T citotóxicos (CD8+) es ineficiente en la infección primaria; aunado a esto, el hMPV interfiere con la inducción de antígenos timodependientes, debido a que las células dendríticas (a pesar de no ser las primeras células infectadas) son infectadas conforme el virus se disemina, lo que afecta en la inducción de inmunidad a largo plazo, mejor conocida como “de memoria” 3. Se sabe que este tipo de virus también modula negativamente la respuesta de las citosinas; por ejemplo, por un mecanismo desconocido aún, el hMPV evita la fosforilación de Tyk2 y STAT3 y la subsecuente fosforilación positiva de IL-6; la proteína M2-2 promueve la evasión global de los TLR en monocitos; la proteína G interfiere con la producción de INF tipo I porque impide que una de las proteínas (RIG-1) de la cascada de las MAVS se desacople, lo que evita la activación de la señal11.
Diagnóstico
Debido a las similitudes clínicas y epidemiológicas existentes entre hOPV y hMPV, es difícil determinar al agente etiológico responsable. La forma de detección era el aislamiento mediante cultivo celular a partir de muestra biológica (aspirados, hisopado nasal o nasofaríngeo), que se cultivaba para después infectar células inmortales (como HEP-2), para observar la formación de sincicios, pero su diagnóstico era lento.
Posteriormente se han probado “variantes” de la reacción en cadena de la polimerasa retro transcriptasa (RT-PCR) 26, como RT-PCR multiplex para poder identificar 13 posibles patógenos causantes de enfermedad, el cual ha demostrado ser sensible y específico al no arrojar falsos negativos, pero los costos son elevados27, por lo que en la búsqueda de un diagnóstico más rápido y económico se intentó la evaluación de proteínas como anticuerpos (IgG-hMPV) presentes en los aspirados nasofaríngeos por inmunofluorescencia directa (DIF), la cual resultó no ser tan sensible en comparación con RT-PCR pero más accesible26. El diagnóstico sugerido por la Norma Oficial Mexicana 031-SSA-1999 para la atención a la salud del niño comienza con el interrogatorio, la inspección física orientada a la identificación de neumonía, de otitis media aguda, faringoamigdalitis purulenta u otra entidad nosológica de etiología bacteriana28; por otro lado, se reporta que además de la exploración de rutina también se realiza cultivo celular a partir de aspirados nasofaríngeos29.
Tratamiento
En la actualidad el tratamiento de elección contra las enfermedades causadas por hMPV es el que se utiliza para hOPV y son anticuerpos monoclonales murinos humanizados como palivizumab o motavizumab, que es una variante de segunda generación de palivizumab. Ambos se administran de manera profiláctica en infantes en situación de riesgo, que tienen altas probabilidades de contraer la enfermedad12. Además del tratamiento de soporte (nebulización con solución salina hipertónica al 3% y salbutamol) que indica el Catálogo Maestro de Guías de Práctica Clínica IMSS-032-08, se han propuesto la ribavirina, inhibidores de fusión y pequeños ácidos ribonucleicos interferentes como tratamiento en el control contra hMPV, a partir de estudios realizados en diferentes países del mundo como Chile y Canadá3; sin embargo, la ribavirina ya no se usa en México por sus efectos colaterales.
No existe aún una vacuna contra el hMPV que esté aprobada y sea utilizada por el sector salud en el mundo30, pero se han realizado varios intentos por desarrollar la vacuna ideal, la cual, además de ser inmunoprotectora contra hMPV, no cause efectos adversos en el proceso de inmunización.
Vacunas candidatas
Los primeros intentos para obtener una vacuna capaz de inducir una respuesta inmune con base en anticuerpos fueron los virus atenuados, virus inactivados y partículas tipo virales (VLP, por sus siglas en inglés,). Podemos citar un trabajo con virus atenuado que presenta la deleción de la proteína SH de un hMPV (HPMVΔSH), con el que inmunizaron hámsteres dorados. Los resultados mostraron respuesta inmunológica, aunque no lo suficiente para conferir protección, y en cultivo celular demostró replicación eficiente, sugiriendo que la proteína SH no era esencial para la entrada del virus a la célula31. Posteriormente se eliminó la proteína G y se probó en roedores, pero no mostró conferir suficiente protección. Un ensayo más reciente de las VLP fue el realizado por Lévy et al. (2013), donde insertaron proteína F y G de linajes A y B en un core de un retrovirus que mimetizaba las propiedades de ambos linajes de hMPV y cuando se probó en ratones indujo anticuerpos neutralizantes contra hMPV, mostrando protección de los ratones32. Otro ensayo con virus inactivados por formaldehído (FI-HMPV) mostró inducir anticuerpos neutralizantes (los que se unen al virus e inhiben su fusión) en cotton rats y en otros animales; sin embargo, en macacos fue menos efectiva, por lo cual se debe mejorar33.
Proteínas de hMPV como posibles vacunas
De las 9 proteínas traducidas por el virus, solamente 3 (F, G y SH) se encuentran insertas en la membrana con una extremidad saliente, por lo cual son posibles candidatas a ser reconocidas por alguna célula presentadora de antígeno, para desencadenar reacciones inmunológicas que confieran protección al hospedero. Se han realizado varios intentos por encontrar el mayor determinante antigénico (de las 3 proteínas ancladas a la envoltura viral) buscando el epítopo que indujera inmunidad contra hMPV. En un principio se consideró a la proteína G de acuerdo con algunos reportes8,33, pero fue hasta el trabajo de Skiadopoulos et al. (2006), en el que se demostró que la proteína F era más importante debido a que la entrada del virus está mediada por la proteína de fusión. En ese trabajo desarrollaron un modelo de vacuna, insertando en una partícula vacía del virus de parainfluenza humano (hPIV) recombinante (rPIV) las proteínas a probar; un diseño fue con la proteína F, y se obtuvo que la inducción de anticuerpos fue similar a la inducida por un hMPV WT, lo cual sugirió protección ante el patógeno, mientras que cuando se insertó solamente la proteína G, la respuesta fue menor con respecto a la que induce hMPV WT y no indujo anticuerpos neutralizantes; finalmente, con la proteína SH no se indujeron anticuerpos neutralizantes34.
A partir de esto, se han diseñado vacunas que contengan la proteína F de hMPV o un fragmento de ella, ya sea sola o en compañía de su versión en hOPV, y finalmente una versión quimérica, como una opción de vacuna que confiera protección y no tenga efectos secundarios graves. Por ejemplo, en un estudio realizado por Wen et al. (2016), diseñaron una vacuna quimérica (RPM-1) a partir de las secuencias de aminoácidos conservadas entre las proteínas F de hMPV y hOPV (figura 5), capaz de inducir anticuerpos neutralizantes para ambos virus, pero según los autores, debido a su estructura tridimensional, mostró menor efectividad para hMPV, por lo cual sugieren diseñar otra vacuna que contenga ambas proteínas F de los 2 virus sin ser quiméricas, o sea por separado12. Otro ejemplo de una investigación con un enfoque diferente fue el realizado en Canadá por Skiadopoulos et al. (2004), donde a partir de 2 aislados clínicos de pacientes con infección por hMPV pertenecientes cada uno a uno de los linajes de hMPV (A y B), evaluaron en hámsteres siberianos si ambas cepas podían proteger para su linaje o para ambos linajes y obtuvieron que solo la cepa correspondiente al linaje A protegía para ambos. Posteriormente introdujeron la secuencia de la proteína F en el genoma de PIV recombinante y eliminaron la propia proteína F de PIV; así se obtuvo una vacuna que confiere inmunidad para ambos linajes de hMPV y además para PIV10. Sin embargo, en otro estudio realizado en Francia, también utilizaron aislados de ambos linajes de hMPV para producir VLP con proteínas F de cada cepa, y se demostró que estas proteínas F conferían inmunización cruzada, además de la correspondiente a su linaje, por lo que ellos proponen que esta protección se debe a que dichas proteínas se administraron en su conformación nativa y funcional32. Otro estudio realizado en Pittsburgh demuestra que no es necesaria toda la proteína F para poder conferir inmunidad, ya que con una héptada (HRA-A2) es suficiente en ratones para ser reconocida como un epítopo y obtener inmunidad35. En la tabla 2 se resumen los tratamientos mencionados en esta sección.

Secuencias de alineamiento de la proteína F de hOPV y hMPV junto con la secuencia mutante RPM-1, donde los cuadros amarillos representan los contactos más probables con el anticuerpo comercial motavizumab y los rojos, los sitios de menor probabilidad de contacto, debido a que son menos accesibles. En la parte superior de las secuencias, las letras (H/L) indican el contacto de dicho aminoácido con la cadena pesada o ligera (H o L respectivamente, por sus siglas en inglés), en las regiones CDR (hipervariables) de la parte Fab del anticuerpo, en la parte inferior de las secuencia RPM-1, las letras C representan los aminoácidos cargados, el guión medio (-) los no cargados, el punto (.) sobre la secuencia de hMPV indica los que son de la misma carga hOPV y hMPV. Tomado de Wen et al., 2016.
Figura 5 Proteína quimérica (RPM-1)
Tabla 2 Estrategias experimentales de tratamiento contra hMPV
|
Estrategia
de control |
Producto | Modelo usado | Resultado | Referencia |
| VLP | HPMVΔSH | Hámster dorado | Mostró respuesta inmunológica, aunque no lo suficiente para proteger al espécimen, además de que la deleción en realidad no atenuó en su totalidad al virus | Biacchesi et al., 2004 |
| Virus atenuados | FI-HMPV | Cotton rats/ macacos | Mostraron inducir anticuerpos neutralizantes en cotton rats, sin embrago, en macacos fue menos efectiva | Schuster and Williams, 2014 |
| VLPs | rHPIV-1 | Hámster dorado | Cuando se insertó proteína F en el virus recombinante, la inducción de anticuerpos fue similar a la inducida por un hMPV WT, lo cual sugirió protección ante el patógeno, mientras que cuando se insertó la proteína G la respuesta fue menor con respecto a la que induce hMPV WT y no indujo anticuerpos neutralizantes | Skiadopoulos et al., 2006 |
| Vacuna | RPM-1 | Ratones BALB/c | Fue capaz de inducir anticuerpos neutralizantes para ambos virus, pero según los autores, debido a su estructura tridimensional, con menor efectividad para hMPV, por lo cual sugieren diseñar otra vacuna con los mismos epítopos, pero separados | Wen et al., 2016 |
| Proteína | Proteína F en una partícula de PIV | Hámster siberiano | Confirió inmunidad para ambos linajes de hMPV y además para PIV | Skiadopoulos et al., (2004) |
| Péptido | HRA-A2 | BALB/c | Demostró que es capaz de inhibir la fusión viral en células HEp-2 y la enfermedad en ratones BALB/c | Deffrasnes et al., (2008) |
BALB/c: cepa de ratones albinos; FI-hMPV: metapneumovirus inactivados por formaldehido; HPMVΔSH: metapneumovirus humano con la proteína SH deletada; HRA-A2: fracción de la proteína F expresada en bacterias; rHPIV-1: partícula recombinante de respirovirus humano -1 (anteriormente parainfluenza virus humano tipo 1); RPM-1: proteína quimérica de proteínas F de hMPV y hOPV; VLP: Partículas similares a virus (por sus siglas en inglés).
Péptidos antivirales
Los péptidos antivirales son una alternativa profiláctica y emergente en enfermedades infecciosas causadas por virus, los cuales tienen el objetivo de inhibir la infección de las células por el virus. Estos péptidos actúan profilácticamente y, debido a su tamaño pequeño, no confieren protección inmunológica de memoria, por lo que deben aplicarse en la mucosa en varias dosis.
La inhibición de la fusión virus/célula se puede lograr ya sea porque el péptido se una al virus y así evita que este se una a su receptor en la célula -como el reportado por Ortega-Berlanga et al. (2016) para el hOPV, en donde el péptido antiviral RhoA al unirse a la proteína F de hOPV inhibía la fusión hOPV/células HEp-2 (36)- o porque el péptido compita con el receptor celular con el virus, como el reportado por Deffrasnes et al. (2008). Con la finalidad de inhibir la fusión del hMPV a sus células blanco se desarrollaron péptidos sintéticos y recombinantes, basándose en los péptidos HRA y HRB; en los ensayos de inhibición realizados en cultivos celulares, descubrieron que las héptadas A recombinantes eran más efectivas en bajas concentraciones en comparación con las HRB y con las propias HRA sintéticas. Posteriormente realizaron retos virus/péptido en ratones BALB/c y reportaron que el péptido HRA2 (compuesto por 50 aa del péptido HRA original del virus más 52aa del péptido pET donde fue clonado) era capaz de inhibir la fusión y por lo tanto la replicación viral37.
CONCLUSIONES
El hMPV es considerado como un virus emergente responsable de las IRA, principalmente en pacientes pediátricos con altos índices de morbimortalidad. La identificación de este patógeno como agente causal de forma individual o en coinfección, es difícil, a menos que se cuente con infraestructura y personal adecuado para su diagnóstico utilizando métodos inmunológicos o moleculares, además, no existe tratamiento antiviral específico ni vacunas para la profilaxis aprobado por el sector salud contra hMPV, por lo que es un nicho de investigación adecuado. Existen varias propuestas de vacunas tales como: VLPs, Virus atenuados, proteína F y péptidos; los resultados reportados en modelos animales particularmente con VLPs con la proteína F demostraron que este tipo de candidato a vacuna, induce los 2 tipos de respuesta Th1 y Th2 con una reacción inflamatoria leve y debe seguirse trabajando en ella. No obstante, también se propone el uso de proteínas o péptidos profilácticos como HRA-A2 que al ser una réplica exógena de un fragmento de la proteína F, puede impedir la infección viral, pero debe suministrarse en varias dosis por aspersión en mucosa respiratoria.

