











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink

Introducción
Los inflamosomas son complejos proteicos localizados en el citoplasma de las células, que actúan como sensores y son mediadores del desarrollo de la inflamación1. Existen diferentes inflamosomas, pero todos ellos están relacionados con el reconocimiento de patrones conservados de secuencias proteicas a través de los receptores tipo PPR, que son receptores que reconocen patógenos2. Estos últimos se subdividen en 4 categorías: los receptores tipo Toll (TLRs), los receptores tipo RIG-I (derivado del inglés retinoic acid-Inducible gene 1), los receptores de carbohidratos tipo lectina (CLRs) y los receptores tipo NOD (derivado del inglés nucleotide-binding oligomerization domain)3. Estos últimos conforman un grupo especializado de proteínas intracelulares presentes en el citoplasma que se consideran esenciales para detectar patógenos e iniciar la respuesta inmune innata.
A los receptores tipo NOD de aquí en adelante los llamaremos NOD-like receptor (NLR). Los NLRs están divididos en 3 subfamilias clasificadas de acuerdo con los dominios de interacción con proteínas que poseen en la región amino terminal: 1) NOD que consiste de 6 miembros diferentes -5 NOD y 1 CIITA (Class II transactivator)-, 2) IPAF o NLRC, que está formado por 2 miembros, y 3) NLRP, del que se reconocen 14 diferentes miembros (NLRP1 hasta el 14). Esta última subfamilia, la de los NLRP, es la que se relaciona con la formación de inflamosomas, en particular NLRP1, NLRP3, NLRP7 Y NLRC4 (NLR family CARD domain containing 4)2,4. Los NLRs responden a señales exógenas que provienen de bacterias, hongos o virus que contengan PAMPs (pathogen-associated molecular patterns), o a señales endógenas que son moléculas conocidas como DAMPs (damage-associated molecular patterns), también conocidas como alarminas, las cuales son liberadas por las células dañadas ya sea por cáncer, trauma, necrosis o isquemia. Estas moléculas son los mediadores de la inducción de la inflamación asociada a los procesos de estrés5. A la fecha se ha reconocido a la proteína HMGB-1 (high mobility group box-1)6, la calgranulina A y B, y el amiloideo sérico tipo A como inductores de estrés. La señalización que inducen estas moléculas esta mediada por receptores específicos que incluyen a los receptores para productos terminales con glicación avanzada (RAGE), los receptores TREM1 (triggering receptor expressed on myeloid cells 1), TLR2, TLR4 y un receptor transmembranal tipo lectina-C que se conoce como Mincle. El inflamosoma NLRC4, que tiene cierta similitud estructural con NLRP3, se activa por bacterias Gram- que poseen sistemas de secreción tipo III o IV (T3SS, T4SS) o flagelina que entra al citosol a través de los T3SS o T4SS como sucede en la infección por Salmonella, Shigella, Pseudomona o Legionela7.
Los inflamosomas, que como ya mencionamos anteriormente son complejos formados por diferentes proteínas8 que describiremos más adelante, activan a la caspasa-1, también conocida como enzima convertidora de la interleucina-1, cuya función es separar los componentes de la forma precursora de la IL-1b e IL-18, generando así los péptidos maduros. Recientemente se ha descrito que la caspasa-1 también actúa sobre la IL-339; sin embargo, la información es controversial, ya que algunos autores mencionan que el efecto de la casapasa-1 es necesario para generar la molécula biológicamente activa IL-33112-270, mientras que otros mencionan que la forma activa es la IL-331-270 y que el efecto de la caspasa-1 en la asparagina 178 de la secuencia de la IL-33 genera una molécula biológicamente inactiva. De cualquier manera, e independientemente de la controversia funcional, el común denominador de estas 3 interleucinas es que su ácido ribonucleico mensajero (ARNm) es traducido sin requerir de una secuencia señal para ser secretado.
El inflamosoma más estudiado hasta la fecha es el NLRP3, también conocido como criopirina10. Este inflamosoma está formado por la proteína NLRP3, la proteína adaptadora ASC (apoptosis-associated speck-like protein) y una procaspasa-1(Figura 1). La estructura de la proteína NLRP3 contiene 3 dominios, un dominio de pirina N-terminal (PYD), en el carboxilo-terminal se encuentra un dominio que posee repeticiones ricas en leucina (dominio LRR), y una zona central que se conoce como dominio NACHT (nucleotide binding and oligomerization domain), el cual se une a nucleótidos y es el responsable del proceso de oligomerización del inflamosoma. El dominio de pirina, que también se le llama TRIM20, censa la perturbación inducida por las toxinas bacterianas de la familia de las RhoGTPasas. Cuando no existen las moléculas activadoras de la formación del inflamosoma el dominio NACHT y el dominio LRR de NLRP3 interactúan entre sí suprimiendo así la interacción entre el NLRP3 y la proteína adaptadora ASC, inhibiendo la formación del inflamosoma.

Figura 1 Componentes del inflamosoma NLRP3: la proteína NLRP3, la proteína adaptadora ASC (apoptosis-associated speck-like protein) y una procaspasa-1. La estructura de la proteína NLRP3 contiene un dominio de pirina N-terminal (PYD), un dominio con repeticiones ricas en leucina (dominio LRR), y un dominio NACHT (nucleotide binding and oligomerization domain), que es el responsable del proceso de oligomerización del inflamosoma.
El dominio CARD es un dominio plegado de muerte, similar a PYD, pero formado por 6 alfa hélices antiparalelas con una región central hidrofóbica, y en la cara externa formada por aminoácidos cargados; PYD está formado por solamente 2 alfas hélices y también tiene una región central hidrofóbica. Es de llamar la atención el hecho de que el dominio PYD en la molécula adaptadora ASC tiene 2 sitios de unión, uno que se une al PYD de NLRP3 y el otro que se une a la proteína moduladora de la formación del inflamosoma llamada POP111; la fuerza de unión de POP1 es pobre, lo que facilita que en presencia de proteínas inductoras de la formación del inflamosoma POP1 se desprenda y ese sitio de unión sea utilizado para reforzar la unión con NLRP3. La lista de proteínas que contienen dominios CARD es muy extensa pero la gran mayoría de ellas (caspasas 1, 2, 4, 5, 9, 12, 13, ICEBERG, MDA-5, MAVS, CRADD, RAIDD-2, RIG-1, CARD 8, 9, 10, 11, 14, APAF1, GLAVA1, etc.) se relaciona con procesos de apoptosis e inflamación.
El dominio pro-caspasa está formado por los 2 componentes activos de la caspasa-1, que se conocen como p10 y p20, que están unidos a un dominio CARD. La Figura 2 es una representación de cómo los 3 componentes básicos del inflamosoma, se unen entre si para formar un dímero y posteriormente el tetrámero que es la forma activa del inflamosoma. La liberación de p10/p20 se debe a la acción de una caspasa-14 que se encuentra en el citoplasma de las células y es parte de la familia de las caspasas inductoras.
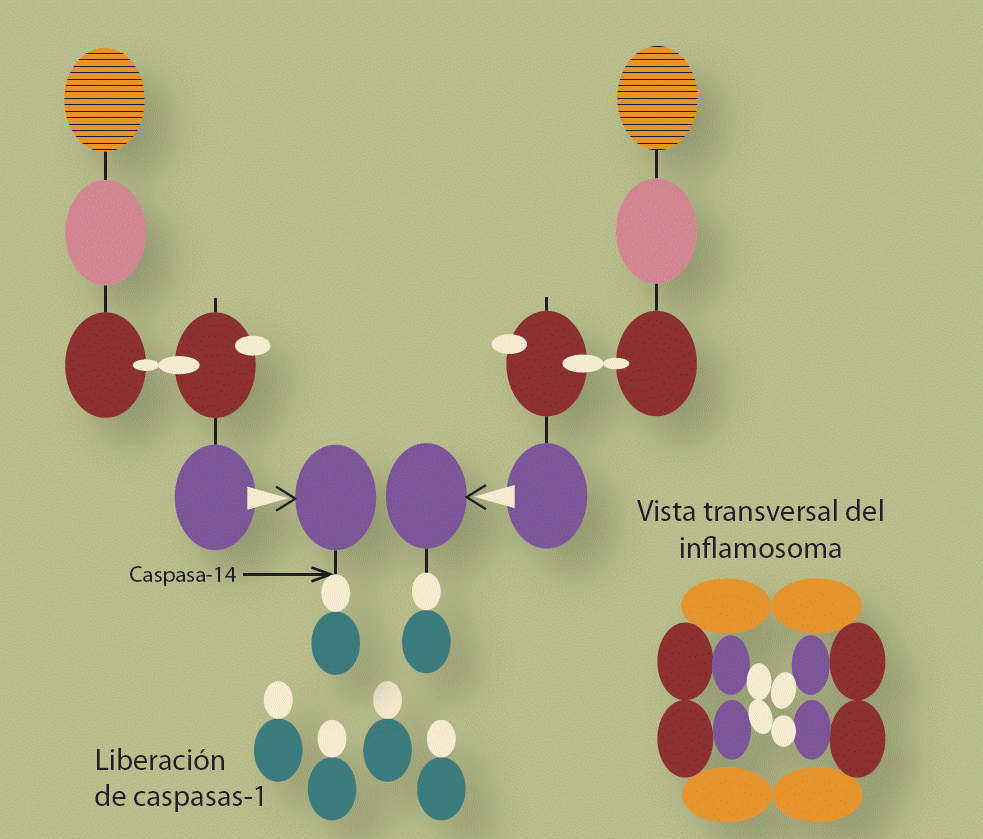
Figura 2 Representación gráfica de la formación del inflamosoma. Está claro que se requieren al menos 2 elementos de cada uno de los componentes del inflamosoma. La liberación de p10/p20 de la proteína NLRP3 se debe a la acción de una caspasa. El ensamble de los componentes dispara una modificación conformacional que induce su activación y oligomerización en un tetrámero que conlleva a la activación de la caspasa-1 y ésta a su vez a la secreción de IL-1b, IL-18 e IL-33.
El ensamble de los diferentes componentes que conforman el inflamosoma NLRP3 dispara una modificación conformacional que induce su activación y oligomerización en un tetrámero (Figura 2), lo cual a su vez conlleva a la activación de la caspasa-1 y ésta a su vez, a la secreción de IL-1b e IL-18 que resulta en inflamación y también en un proceso de lisis celular llamado piroptosis que se da cuando el proceso de oligomerización no se lleva a cabo12. Al parecer la proteína de choque térmico conocida como HSP90 y la proteína asociada con la ligasa de ubiquitina conocida como SGT1 regulan la formación y la activación del inflamosoma NLRP313.
La activación del inflamosoma se inicia con la fosforilación del receptor específico que se activa ya sea porque reconoce PAMPs o DAMPs;14 la fosforilación del receptor activa al factor de transcripción NF-kB, el cual se trasloca al núcleo de la célula y promueve la transcripción de la proteína NLRP3, proIL-1b, proIL-18, proIL33 y HMGB-1, que se mantienen en el citoplasma de la célula de forma inactiva15. Este proceso se reconoce como la primera señal de inducción para la formación del inflamosoma. La presencia de una segunda señal de activación está dada por la presencia de especies reactivas de oxígeno en el citoplasma, la salida de potasio a través de poro purogénico P2X7 y la entrada de calcio, daño mitocondrial, la ruptura de vesículas de fagolisosomas que liberan catepsinas al espacio intracelular, o bien la existencia de estrés en el microambiente celular; cualquiera de estas vías de activación favorece que se “abra” NLRP3 e interactúe el dominio PYD de NLRP3 con la proteína adaptadora ASC induciendo así la oligomerización del NLRP3 con la proteína ASC (apoptosis-associated speck-like protein); posteriormente el dominio CARD (caspase activation and recruitment domain) de ASC se une al dominio CARD de la procaspasa-1 dando lugar a la formación del inflamosoma NLRP3 que es en realidad un complejo. Este complejo funciona como catalizador para que la caspasa-14, que es una forma de caspasa de activación corte a la procaspasa-1, entre el dominio CAD y las regiones activas p10/p20 y genere el tetrámero activo caspasa-1/p10/p20, que es una proteasa de cisteínas que al activarse corta a la pro-ILb, pro-IL-18 y pro-IL-33, lo que contribuye a la activación y secreción de IL-1b, IL-18 e IL-33 maduras (Figura 3)16,17. Recientemente se describió que la fracción ASC del NLRP3 se libera hacia la circulación donde funciona como un complejo oligomérico extracelular que genera señales de daño amplificando así la respuesta inflamatoria18.
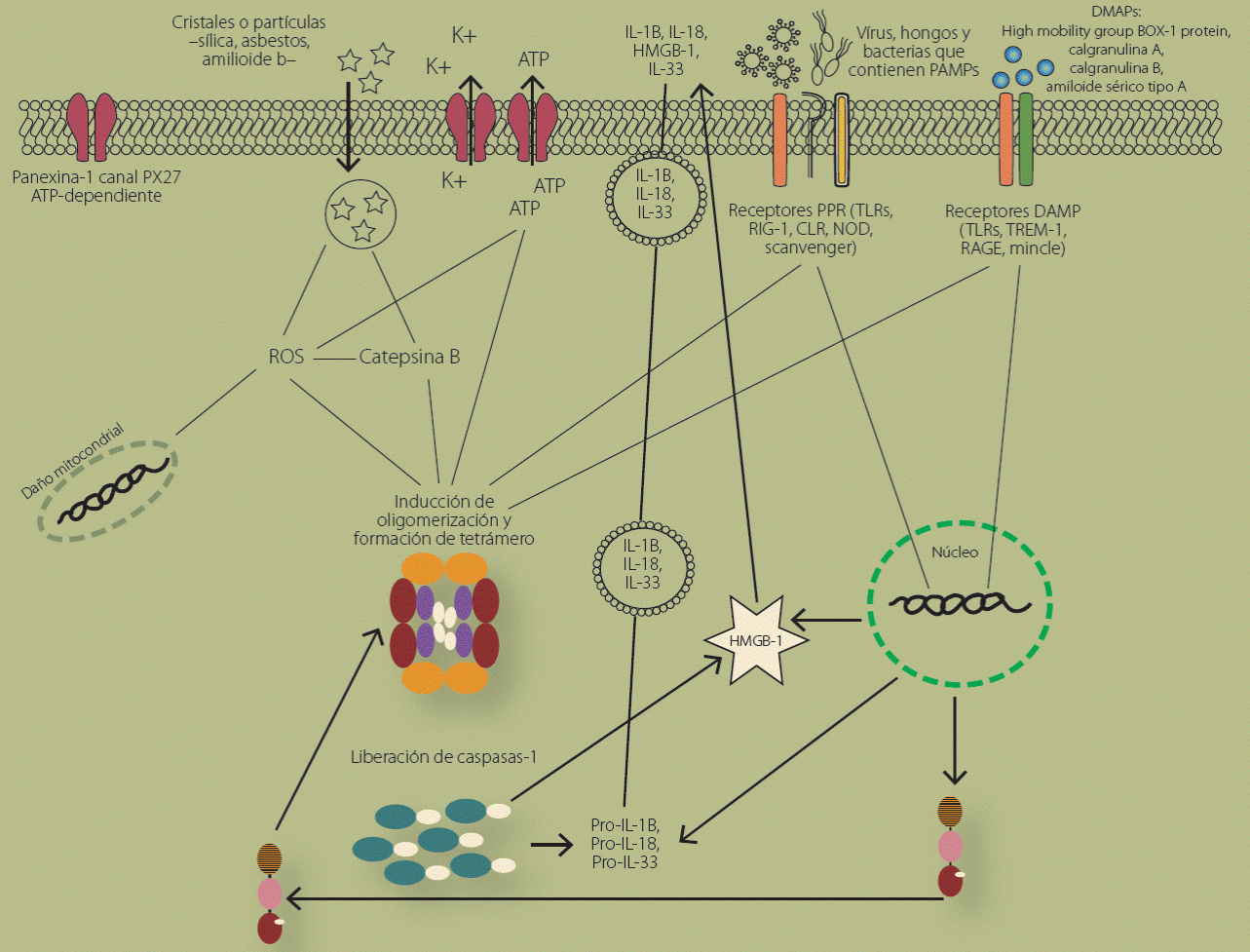
Figura 3 Esquema que representa las moléculas y los mecanismos que activan la formación del inflamosoma. La explicación más detallada de todos estos procesos y su influencia se describe con detalle en el texto.
El inflamosoma NLRP3 se encuentran primordialmente en macrófagos, monocitos, células dendríticas y neutrófilos, y ayuda en la defensa contra infecciones, pero también pueden inducir choque séptico o empeorar la ateroesclerosis cuando se activa de manera no controlada o se inhibe a la cinasa kB19. El lipopolisacarido de la pared de bacterias gramnegativas o el ATP extracelular activan a la caspasa-1 del inflamosoma NLRP320, así como también el amiloide b fibrilar, el hidróxido de aluminio, los asbestos, los cristales de colesterol, las lipoproteínas oxidadas de baja densidad, la hiperglicemia, los ácidos grasos libres o los cristales de acido úrico. Existe una larga lista de virus que activan al inflamosoma NLRP3, entre los que sobresalen el virus Sendai, el adenovirus, el virus de la encefalomiocarditis y el virus de la influenza. La presencia de ATP extracelular estimula el canal iónico purogénico P2X7, lo que dispara la salida de K hacia el espacio extracelular e induce de esta manera el reclutamiento del poro de membrana llamado panexina-1, que facilita el ingreso de moléculas de ATP hacia el espacio intracelular. Es importante hacer notar que existe una vía de inflamosoma no canónica en la que la caspasa-4 y la -5 ejercen el efecto sensor y el efecto efector. La caspasa 4 o 5 detectan la presencia de LPS en el citosol a través del reconocimiento del lípido A, que es parte constitutiva del LPS bacteriano, con el dominio CARD; esta vía no canónica solo induce el proceso de piroptosis y no activa la secreción de citosinas proinflamatorias debido a que no favorece la oligomerización del inflamosoma. Esta vía no canónica requiere que las vacuolas citoplásmicas que contienen al patógeno se lisen para que se libere el LPS del patógeno al citoplasma.
Afortunadamente existen una serie de inhibidores del inflamosoma que incluyen al hidroxibutirato, los interferones tipo 1, el micro RNA223 y el resveratrol21. Este último es una molécula antioxidante formada por un fenol y una fitoalexina que se encuentra en la pared de las uvas negras y los frutos rojos, y por lo tanto es un componente muy importante del vino tinto22.
¿Cuál es la relevancia clínica de los inflamosomas?
Constantemente, nuestra actividad médica está enfrentándonos a eventos mediados por los inflamosomas; aun más, no existe proceso inflamatorio en el que no esté involucrado alguno de los múltiples inflamosomas que hemos mencionado6,23. De manera muy particular se puede decir que la activación del inflamosoma NLRP3 se relaciona con alteraciones en la concentración intracelular de potasio, la presencia del virus de la influenza, la bacteria Neisseria gonorrhoae, las partículas contaminantes ambientales, los cristales de colesterol o los cristales de uratos, los asbestos, la progresión de la esclerosis múltiple, la encefalomielitis autoinmune, la enfermedad de Alzheimer24, la colitis ulcerativa crónica inespecífica, la enfermedad de Crohn25, el síndrome de fiebre periódica asociada a criopirina y muchos síndromes clínicos más. Mutaciones en los genes de NLRP3 también se asocian con el desarrollo de enfermedades autoinmunes y autoinflamatorias, entre estas últimas sobresale el síndrome autoinflamatorio familiar frío o síndrome neurocutáneo infantil crónico2.
En el caso particular de la enfermedad de Alzheimer se sabe que la fracción 1-42 del amiloide-b en su forma soluble es internalizado en la neurona por el CD36, que es un receptor scanvenger; este amiloide-b, así como fibras de amiloide-b intracelulares y extracelulares, estas últimas internalizadas por los receptores para glutamato NMDA (que son receptores de N-metil-D-aspartato), forman un fagolisosoma que se modifica y libera al espacio intracelular catepsina B la cual activa la formación del inflamosoma. Es obvio que los inhibidores de catepsina B (como el ibuprofeno)26 tienen aplicación clínica en esta enfermedad, ya que al prevenir la activación del inflamosoma disminuirá la secreción hacia el espacio extracelular de especies reactivas de oxígeno, que tienen el potencial de inducir la formación de fibrillas de amiloide-b y de citocinas proinflamatorias (IL-1b, IL-6, IFNy, IL-8), que se mantienen en el cerebro o son drenadas a la circulación periférica un mediante mecanismo de drenaje antigénico bien descrito. La acumulación de más fibrillas de amiloide-b se da, entre otras causas, por la función activadora del IFNg sobre las g-secretasas, que son las enzimas que liberan el fragmento tóxico amiloide-b 40/42.
Recientemente se observó en ratones con deficiencia de NLRP3 que las células de la microglia, que en realidad son macrófagos cerebrales, se derivan hacia el fenotipo M2 que se relaciona con mayor secreción de IL-4 (típica de respuestas Th2, es decir, antiinflamatorias) y reducción de depósitos cerebrales de amiloide-b27. Toda esta información deja de manifiesto que la activación del inflamosoma NLRP3 juega un papel primordial en el desarrollo de la enfermedad de Alzheimer1, lo cual lo convierte en un blanco terapéutico interesante (Figura 4).
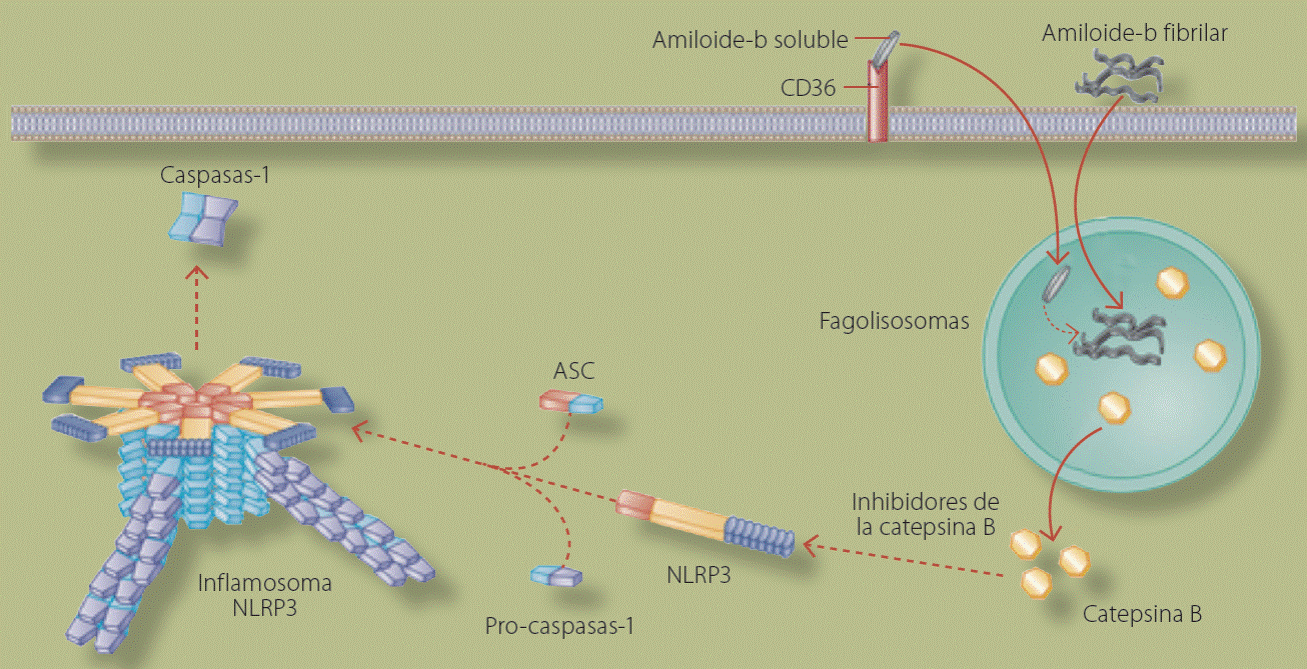
Figura 4 Representación esquemática de la formación y activación del inflamosoma NLRP3 en la enfermedad de Alzheimer. Se observa el efecto inductor del amiloideo betasoluble y del amiloideo estructurado en forma de fibras que son fagocitadas por un fagolisosoma que libera catepsina B que inicia el proceso de inducción de la formación del inflamosoma. Como se puede observar, el uso de inhibidores de catepsina podría tener beneficios terapéuticos en pacientes con enfermedad de Alzheimer, ya que disminuiría la inflamación y ésto evitaría la agregación y formación de nuevos depósitos de amiloide intracerebral. Fuente: Guo H et al, 2015.
En conclusión, es importante enfatizar que los inflamosomas son estructuras intracelulares que al tener dos pasos de activación pueden considerarse como reguladores del proceso de la inflamación; sin embargo, la homeostasis que mantienen se puede perder en condiciones de agresión por patógenos o por medicamentos. Un buen ejemplo de esto último es el uso indiscriminado de medicamentos antiinflamatorios, los cuales pueden inducir efectos contrarios a los deseados ya que los medicamentos de este estilo, especialmente los no esteroides, pueden activar al inflamosoma a través de vías dependientes no solo de P2X7 sino también de TLR4.


