











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkOBSERVANDO AL NÚCLEO CELULAR CON OJO HISTOLÓGICO
Una de las primeras preguntas que hacemos los profesores de Biología Celular y Tisular cuando comienza el curso es: ¿cuál es la diferencia entre la célula eucarionte y procarionte? Y los alumnos a coro contestan: que las eucariontes tienen núcleo, y las procariontes, no.
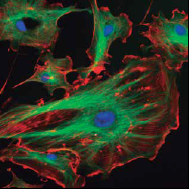
Por supuesto, sabemos que la respuesta va mucho más allá, sin embargo es una diferencia suficiente para comenzar.
EL NÚCLEO DE LA CÉLULA HUMANA
El núcleo suele ser el organelo más evidente de la célula, y es el que contiene la mayor parte del ácido desoxirribonucleico (DNA), ya que una pequeña parte se encuentra en las mitocondrias. La función del núcleo es mantener la integridad del material genético: el DNA nunca sale del núcleo, así que dentro de él se llevan a cabo todas las funciones metabólicas de esta biomolécula, es decir, ahí se lleva a cabo la transcripción, la síntesis y la reparación1-3.
En la interfase, cada célula somática humana contiene en su núcleo 46 cromosomas conformados por 46 hebras de ADN lineal, que suman un total aproximado de 2,000,000 de pares de bases. Si se hiciera una sola hebra de DNA con esos 46 cromosomas, se obtendría un finísimo hilo de 2 metros de longitud. Todo este DNA (junto con una gran cantidad de proteínas) está contenido en un pequeño núcleo que mide 5 micrómetros de diámetro en promedio. Para lograr esta hazaña, la célula confina al DNA dentro del núcleo y lo compacta, desde la doble hélice hasta el cromosoma metafásico (figura 1).
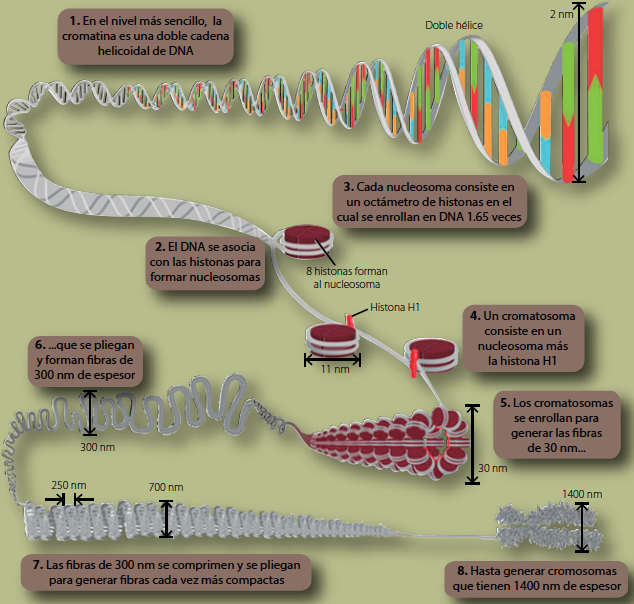
Figura 1 La cromatina es una estructura compleja con varios niveles de organización. En la imagen se describe la compactación desde la doble hélice hasta el cromosoma metafásico (figura modificada de Nature Education4).
En el DNA humano están contenidos aproximadamente 39,000 genes. Cuando un gen se transcribe da origen a un RNA (ácido ribonucléico). Existen muchos tipos de RNA, los más abundantes y los primeros que fueron descubiertos son el RNA mensajero (RNAm), que cuando se traduce da origen a un polipétido, el RNA de transferencia (RNAt), que se encarga de asociarse con los aminoácidos y conducirlos hacia el ribosoma, y el tercero es el RNA ribosomal (RNAr), que forma parte del ribosoma, el organelo que se encarga de llevar a cabo la traducción1-3.
Para que haya expresión génica, el DNA debe estar en su forma laxa, ya que la doble hélice debe abrirse para permitir el paso de la maquinaria metabólica que lleva a cabo la transcripción, tal como las polimerasas, helicasas, ligasas, entre otras. Si la cromatina está en su forma condensada, entonces este proceso no puede ocurrir y no se lleva a cabo la expresión de los genes contenidos en la heterocromatina5.
Todas las células vivas expresan genes, unas más y otras menos, esto depende del tipo celular; por ejemplo, una neurona o un hepatocito son células metabólicamente muy activas y la evidencia de ello la observamos claramente en sus núcleos: la cromatina que predomina en ellos es la eucromatina. Por el contrario, los linfocitos en sangre periférica son células que sólo están de paso en ese tejido, por lo tanto sus actividades metabólicas están al mínimo, lo que es observable a través del microscopio, ya que su núcleo se observa teñido de manera intensa, esto es, la cromatina que predomina está condensada (heterocromatina)1-3.
EL NÚCLEO VISTO CON EL MICROSCOPIO DE CAMPO CLARO
Durante la interfase, el núcleo está constituido por la envoltura nuclear, el núcleoesqueleto, el nucléolo, la matriz nuclear y la cromatina1-3.
La “envoltura nuclear” está formada por la membrana nuclear externa, el espacio perinuclear, la envoltura nuclear interna y los complejos del poro nuclear. La membrana nuclear externa es una continuación del retículo endoplásmico rugoso, por ello es común encontrar ribosomas adosados a su superficie. La membrana nuclear interna es la membrana que está asociada con la superficie interna del núcleo, a ella se ancla el núcleoesqueleto. Los poros nucleares son espacios en ambas membranas, estos poros están ocupados por proteínas y juntos, el poro y las proteínas, conforman al “complejo del poro nuclear”. Casi todo el tránsito molecular núcleo-citoplasma y citoplasma-núcleo depende de dichos complejos y de su interacción con las proteínas transportadoras (importinas y exportinas).
El “núcleoesqueleto” es el “citoesqueleto” del núcleo, está compuesto por láminas nucleares (A y B, principalmente) que son proteínas de la familia de los filamentos intermedios. Las láminas nucleares constituyen una red proteica que se encuentra asociada a la membrana nuclear interna. Este componente es el responsable de la morfología nuclear y de proveer la estructura sobre la cual se ordena la cromatina.
El “nucléolo” es un organelo que está dentro del núcleo. En el nucléolo se lleva a cabo la síntesis y el ensamblaje de las subunidades ribosomales. Es un organelo altamente basófilo, porque es una zona rica en RNA ribosomal. La información genética necesaria para hacer RNA ribosomal y proteínas ribosomales, se encuentran en los cromosomas organizadores nucleolares (pares 13, 14, 15, 21 y 22). Cada célula tiene al menos un nucléolo (que puede o no ser evidente al microscopio de campo claro), porque todas necesitan ribosomas para llevar a cabo la traducción de RNAm. Otro indicador de la actividad metabólica celular, es el número de nucléolos visibles.
La “matriz nuclear” es el “citoplasma” de este organelo, en ella se encuentran metabolitos y proteínas como las polimerasas.
La “cromatina” es el DNA y sus proteínas asociadas (principalmente histonas). Dentro del núcleo, la cromatina está en dos estados de compactación diferentes: en forma de cromatina laxa, denominada “eucromatina” y la cromatina compacta o condensada llamada “heterocromatina”. A su vez, la heterocromatina, se clasifica de acuerdo con su actividad transcripcional y puede ser heterocromatina constitutiva (actividad génica nula) o facultativa, es decir cromatina silenciada, como es el caso de uno de los cromosomas X en las mujeres, que forma al corpúsculo de Barr (figura 2).
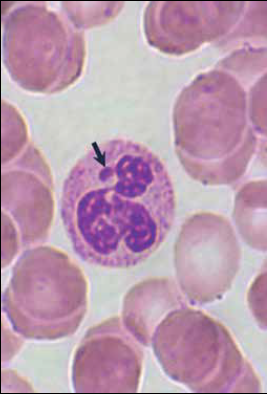
Fotos: Cortesía del autor
Figura 2 Neutrófilo de sangre periférica en el que la flecha señala al cromosoma X condensado, denominado corpúsculo de Barr (tinción de Wright, magnificación 330X. Fotografía: Armando Zepeda Rodríguez y Francisco Pasos Nájera. La preparación histológica pertenece a la colección para docencia del Departamento de Biología Celular y Tisular, Facultad de Medicina, UNAM).
De acuerdo con el tipo celular, los núcleos pueden tener predominancia de eucromatina o de heterocromatina, en la mayoría de las células la cantidad de eucromatina y de heterocromatina son similares en el núcleo.
Morfología nuclear: Aunque siempre imaginamos que las células tienen núcleo esférico, existen otras muchas formas nucleares que son específicas para el tipo celular, por ejemplo, los monocitos tienen núcleo en forma de riñón (figura 3a), los neutrófilos tienen núcleo multilobulado (figura 3b), y las células musculares lisas y estriadas, alargados.
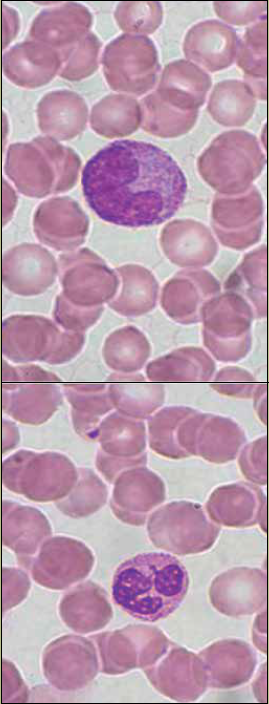
Figura 3 Morfología nuclear. a) En el centro de la imagen se observa un monocito de sangre periférica, con el núcleo en forma de riñón. b) se muestra el núcleo multilobulado de un neutrófilo (tinción de Wright, magnificación 330X. Fotografía: Armando Zepeda Rodríguez y Francisco Pasos Nájera. La preparación histológica pertenece a la colección para docencia del Departamento de Biología Celular y Tisular, Facultad de Medicina, UNAM).
¿POR QUÉ EL NÚCLEO SE VE AZUL-MORADO?
Características tintoriales: El DNA al ser una biomolécula de naturaleza ácida se tiñe con el componente básico (hematoxilina) de la tinción convencional (H-E: hematoxilina-eosina). Sin embargo, las características tintoriales del núcleo se observan de acuerdo con la actividad metabólica de la célula, ya que la heterocromatina se tiñe intensamente (figura 4c) y la eucromatina se observa pálida (figura 4a).
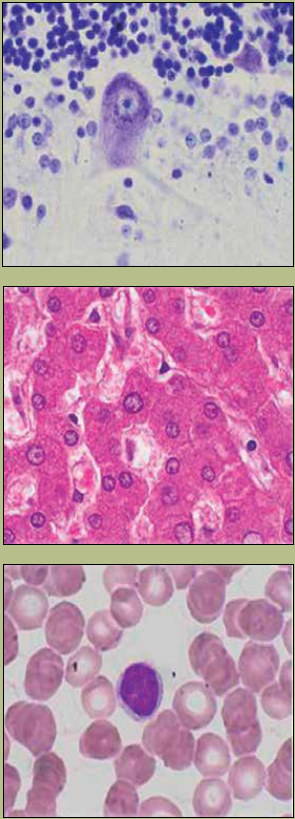
Figura 4 Se muestran diferentes tipos celulares: a) Neurona de Purkinje en cerebelo: se observa el núcleo de tinción pálida y nucleolo evidente (tinción Kluver-Barrera, magnificación 132X). b) Hepatocitos: se observa la eucromatina pálida y la heterocromatina de tinción intensa en la periferia del núcleo (tinción hematoxilina-eosina, magnificación 132X). c) Linfocito de sangre periférica: se observa el núcleo de coloración intensa (tinción de Wright, magnificación 330X). Fotografías: Armando Zepeda Rodríguez y Francisco Pasos Nájera. Las preparaciones histológicas pertenecen a la colección para docencia del Departamento de Biología Celular y Tisular, Facultad de Medicina, UNAM.
La heterocromatina constitutiva se observa como una línea intensamente teñida en la periferia del núcleo (figura 4b), ya que este tipo de cromatina tiene funciones estructurales, es decir, se asocia con el núcleoesqueleto. Esta cromatina es muy importante, porque al no tener actividad transcripcional (genes) cuando esta cromatina sufre daños, éstos pueden ser reparados, y si hay errores durante la reparación, la célula no tiene consecuencias graves.
La heterocromatina facultativa es cromatina que tiene genes útiles, sin embargo, la célula la silencia hipercompactándola. Pero, si es necesario que la célula exprese genes que están en esa forma, simplemente los descondensa para poder transcribirlos. Es decir, este tipo de cromatina tiene la “facultad” de ser eucromatina en un momento de la vida de la célula o heterocromatina en otro.
La eucromatina es la cromatina con actividad transcripcional, es la que contiene a las secuencias génicas que se están expresando en esa célula. Al estar en la forma laxa, la cantidad de colorante que puede captar es menor que en el caso de la heterocromatina, es por ello que es pálida cuando se observa con microscopía de campo claro, como es el caso de la cromatina en células como las neuronas.
Y… ¿TODAS LAS CÉLULAS TIENEN NÚCLEO?
Cuando decimos: “todas las células eucariontes tienen núcleo”, es posible que algún alumno levante tímidamente la mano y pregunte: ¿entonces el eritrocito es una célula procarionte porque no tiene núcleo?
Cabe hacer la aclaración que hay dos tipos celulares en nuestro organismo que están tan diferenciados que sacrifican al núcleo (y al resto de sus organelos) en pos de llevar a cabo su función: los eritrocitos (figura 5) y las células que conforman al cristalino. Sin embargo, siguen siendo células eucariontes.
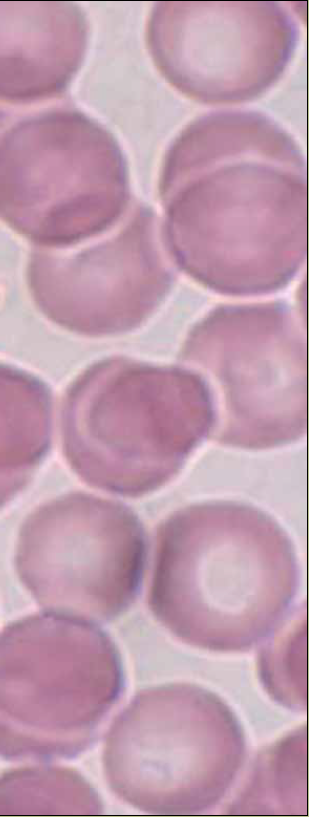
Figura 5 Eritrocitos de sangre periférica, el tipo celular anucleado más abundante del organismo (tinción de Wright, magnificación 330X. Fotografía: Armando Zepeda Rodríguez y Francisco Pasos Nájera. Las preparaciones histológicas pertenecen a la colección para docencia del Departamento de Biología Celular y Tisular, Facultad de Medicina, UNAM).
Y AHORA… ¿CÓMO ENTENDER EL PASO DE LA HISTOLOGÍA A LA BIOQUÍMICA?
La sección anterior explica las características del núcleo cuando la célula está en interfase. Pero cuando la célula va a entrar en la fase de división celular, además de generar nuevos organelos para repartirlos de manera equitativa entre las células hijas, también debe duplicar su material genético, ya que cada célula nueva, debe contener un juego completo, y de preferencia perfecto, de cromosomas (46 en células somáticas y 23 en células germinales).
El ciclo celular se divide en las fases G1, S, G2 y M (mitosis). G1 es el período durante el cual la célula sintetiza biomoléculas y crece. La fase S se caracteriza porque la célula sintetiza una copia exacta de su DNA; este proceso no modifica el aspecto del núcleo. En esta fase también se duplica el centrosoma y los dos centriolos. La fase G2 es período de crecimiento y de revisión: todo debe estar perfecto para iniciar el proceso de división. La fase M a su vez se divide en 5 etapas: profase, prometafase, metafase, anafase y telofase1-3.
La profase se distingue porque en el núcleo se vuelve evidente que la cromatina se está condensando en su forma de cromosomas. La prometafase se caracteriza por la desaparición de la envoltura nuclear. Durante la metafase la cromatina se encuentra en su máximo nivel de compactación, es decir en forma de cromosomas metafásicos, que se alinean en el ecuador de la célula. Durante la anafase, las dos cromátidas que conforman a cada cromosoma, son separadas y conducidas por el huso mitótico hacia los polos. La telofase es el período en el cual se forman nuevas envolturas nucleares, los cromosomas de las células hijas se descondensan, y forman regiones heterocromáticas y eucromáticas iguales a las de la célula que las originó1.
EL OJO BIOQUÍMICO OBSERVANDO AL NÚCLEO CELULAR
Cuando el DNA está en su forma de “cromosoma metafásico” es porque ha llegado al máximo nivel de compactación que puede alcanzar y esto ocurre porque la célula se encuentra en el proceso de división. La información genética se encuentra contenida en el DNA, la estructura de esta estructura fue descrita por Watson y Crick en 1953; este modelo cumple con cuatro puntos indispensables para cualquier material genético que son:
Se puede explicar el proceso de replicación o duplicación del material genético.
Permite explicar la especificidad del material y que éste se preserve a lo largo del proceso de duplicación.
El DNA es la molécula en donde se guarda la información genética.
Por último, el modelo explica cómo el DNA puede mutar6.
En todos los organismos, el DNA se encuentra organizado; en los procariontes, aunque carecen de núcleo, el DNA no se encuentra disperso en el citoplasma celular sino que se organiza en una sección del citosol. Uno de los procesos que permite mantener y transmitir la información genética de una célula a otra es la duplicación o replicación del DNA. Se han realizado diversos estudios para comprender como se lleva a cabo este proceso y los primeros organismos para estudiar, por su relativa facilidad, son las bacterias. En ellas, una vez que se reconoce el sitio de “origen de la replicación” (oriC) que consta al menos de 245 pares de bases (pb). El oriC consta de secuencia repetida de adenina (A) y timina (T) (13 pb), que se denomina caja TATA, en esta secuencia se unirá la proteína iniciadora DnaA, una helicasa Dna B y el cargador de la helicasa Dna C a lo largo del proceso se van uniendo estos complejos hasta que se logran tener sobre el DNA juegos Dna B con Dna C, cada una de estas unido a ATP. Estos complejos proteicos cumplen funciones específicas, la cuales son: Dna C permite la unión de Dna B a la cadena de DNA, esto va asociado con la hidrólisis de ATP; y el Dna B rompe los puentes de hidrógeno que mantienen unidas a las cadenas sencillas de DNA, y abre la “burbuja de replicación”.
En cada hebra de DNA hay un hexámero de DnaB, que permite que se lleve la replicación en ambas direcciones. Para evitar que las cadenas de DNA se vuelvan a unir, interviene una nueva enzima SSBP (proteína de unión de cadena sencilla), que impide que se cierre el DNA y pueda avanzar la burbuja de replicación, pero cada giro en la molécula genera tensión sobre la hebra por lo que es necesario que una topoisomerasa DNA girasa vaya liberando la tensión de la horquilla de replicación. Para poder completar la replicación se requiere que se genere una plantilla, por lo cual cuando la Dna B avanza se une a ella Dna G (primasa) que se encarga de sintetizar un cebador (plantilla) o “primer” de RNA siendo éste de 10 a 12 ribonucleótidos y aporta el extremo 3’-OH para que la DNA polimerasa pueda unir los demás desoxinucleótidos. Se han caracterizado cinco DNA polimerasas (DNA Pol):
DNA Pol I: lleva a cabo la reacción de polimerización en dirección 5’ - 3’. Tiene actividad de exonucleasa en ambas direcciones y se encarga de eliminar y reemplazar a los “primers de RNA”.
DNA Pol II: tiene actividad de polimerización 5´- 3´, pero carece de actividad de exonucleasa 5’-3’. Repara al DNA.
DNA Pol III: Elonga a la cadena de DNA y tiene actividad de polimerización 5’ - 3’.
DNA pol IV: Repara al DNA y tiene actividad de polimerización 5’- 3’, pero no tiene actividad de exonucleasa.
DNA pol V: solo tiene actividad de polimerización 5’- 3’ y se encarga de reparar daños en el DNA.
Debido a que durante la replicación hay una cadena de DNA que se va replicando de manera discontinua, es necesario pegar los fragmentos de DNA que se fueron generando y que reciben el nombre de “fragmentos Okazaki”, esto se lleva a cabo por la DNA ligasa.
Como ya se mencionó, la replicación en eucariontes es más compleja, aunque sigue el mismo patrón para que se lleve a cabo. Una diferencia es que el número de enzimas que intervienen en el proceso es mayor, lo que implica que está altamente conservado y regulado para impedir que el DNA ubicado en los cromosomas se puede replicar en forma independiente de las funciones y de la actividad celular.
En el núcleo celular de los eucariontes el DNA se encuentra altamente organizado gracias a que existe una fuerte y repetida interacción entre el DNA y las histonas que forman al “nucleosoma”. Éste es la unidad fundamental de la cromatina, el cual se constituye por 147 pb que van a envolver a una histona. El DNA se enrolla sucesivamente para hacer una estructura compacta. Al ir enrollando al DNA quedan regiones expuestas o desnudas (20 a 90 pb), estas regiones sirven como sitios de reconocimiento para la unión con otras proteínas que permiten el inicio de la transcripción del material genético7.
En las células eucariontes el proceso de duplicación es complejo, porque la duplicación empieza en cientos o miles de sitios “oriC”, la activación de estos sitios ocurre durante la fase G1 de la división celular. Un modelo que permite estudiar y acercarse al proceso de duplicación de una célula eucariótica es el hongo Saccharomyces cerevisiae. En este organismo se han identificado varias secuencia conservadas de sitios de iniciación denominadas “secuencias autónomas de replicación” (ARS, autonomously replication sequences) que son secuencias no codificantes de 100-200 bp8. Estas secuencias se encuentran altamente conservadas en las levaduras y en organismos emparentados, pero en otros eucariontes la presencia de estas secuencias permitió aislar al complejo de reconocimiento del origen (ORC), que está formado por 6 proteínas (Orc1-6) y tiene un papel análogo a la DnaA bacteriana9.
Una vez que se une el complejo ORC al sitio de origen de la duplicación, es necesario empezar a unir otras proteína como son Cdc6 y Cdt1. Para que se lleve a cabo la duplicación, se deben reclutar a las 6 proteínas que se denomina “minicromosomas de mantenimiento” (MCM) al ORC10. El complejo MCM es un heteroxámero que presenta actividad de helicasa, lo que permite ir abriendo la doble cadena de DNA, para que se pueda llevar a cabo la duplicación del DNA generando una nueva hebra hija por cada cadena de DNA que se duplique. A pesar de que las enzimas encargadas de llevar a cabo la duplicación del DNA son altamente eficientes, pueden existir errores en el copiado. Estos errores son reparados por enzimas encargadas de esta actividad. Sin embargo, en ocasiones no es posible reparar y por lo tanto el error permanece y se genera una mutación en el DNA. Cuando éste se duplica, la mutación es transferida a las células hijas. Las mutaciones se pueden generar durante la duplicación del material génetico y también cuando la célula se enfrenta con factores o agentes externos capaces de modificar al material genético. Este último punto se abordará más adelante.
¿LA CÉLULA PUEDE TENER ERRORES AL REPLICAR SU DNA?
El DNA es susceptible (al igual que las otras biomoléculas) a presentar daños en su estructura como consecuencia de una gran diversidad de factores. Uno de los problemas que esto conlleva, es que cuando la célula se divide, puede heredar estos daños a las células hijas.
“Genotoxicidad” es el término que se utiliza para designar al daño que sufre el DNA. Los agentes que pueden provocar daño, son “agentes genotóxicos”: la generación de daño al ADN es considerado como un evento inicial importante en la carcinogénesis11, porque es más probable que las células con DNA dañado desarrollen mutaciones después de la exposición a los agentes genotóxicos12.
Hoy día, se utilizan diferentes pruebas, con un alto grado de sensibilidad para evaluar diversos tipos de daño al DNA13. Existe una considerable batería de técnicas para la detección de efectos genotóxicos11 tanto in vitro como in vivo en sistemas celulares procarióticos y eucarióticos13-14 que son utilizadas para evaluar diferentes agentes ya sea en condiciones experimentales, ambientales u ocupacionales11.
La prueba validada internacionalmente para detectar actividad genotóxica, es el ensayo de micronúcleos (MN). Un MN es un cuerpo citoplásmico de naturaleza nuclear que corresponde a material genético no incorporado correctamente a las células hijas durante la división celular15; por tanto, es un pequeño núcleo separado del núcleo principal de la célula, que se produce durante la mitosis (en telofase)16. Los MN (conocidos también como cuerpos de Howell-Jolly) son de forma redonda y de contorno liso17. Reflejan aberraciones cromosómicas que pueden ser debidas a errores durante la replicación del material genético o por la exposición a agentes genotóxicos15. Los MN contienen material genético que puede provenir de fragmentos cromosómicos o cromosomas completos16. El propósito de este ensayo, es identificar agentes o circunstancias que inducen formación de MN, que son resultado de rompimientos cromosómicos (clastogenicidad) o de cromosomas completos (aneugenicidad)17.
El ensayo de MN con laminillas cubiertas con naranja de acridina (NA), aprovecha el proceso de eritropoyesis (producción de eritrocitos) para identificarlos. Durante la eritropoyesis, las células deben pasar por diversos estadios de proliferación y maduración. Si el DNA o el aparato mitótico están dañados, esto se podrá ver reflejado en forma de micronúcleos.
Durante la diferenciación del linaje eritroide, cuando un eritroblasto se desarrolla en un eritrocito policromático o reticulocito (eritrocito joven que aún contiene ácido ribonucleico o RNA), el carácter que define esta etapa es la expulsión del núcleo principal, por tanto, cualquier micronúcleo que se haya formado permanece en el citoplasma de la célula, que ahora es una célula anucleada, lo que facilita la visualización de los micronúcleos17(figura 6).
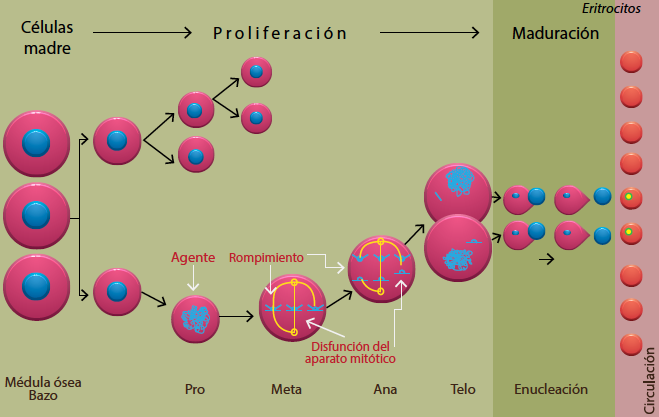
Figura 6 Mecanismo de generación de micronúcleos durante la eritropoyesis en ratón (modificado de Krishna y Hayashi17).
Los reticulocitos, con el tiempo, degradan al RNA y contienen primordialmente hemoglobina, entonces se conocen como eritrocitos normocrómicos (ENC) o eritrocitos maduros. Los reticulocitos y los ENC se tiñen de manera diferencial cuando se fluorocromizan con NA17. Los reticulocitos son células que tienen menos de 24 h de haber sido liberadas a la circulación, por lo tanto, el fluorocromo al interaccionar con el RNA de los ribosomas, emite fluorescencia rojo-naranja, por tanto, pueden ser distinguidos fácilmente de los maduros que no emiten fluorescencia, debido a que éstos no presentan ningún tipo de ácido nucleico en su citoplasma. Los MN son el resultado de daño cromosómico o daño en el aparato mitótico de los eritroblastos16(figura 7).

Figura 7 Los reticulocitos (rojos) contienen un micronúcleo (amarillo) en su citoplasma. Los MN son la evidencia de eventos genotóxicos durante la formación de estas células (coloración supravital con naranja de acridina. La imagen fue obtenida a 100 X. Fotografía: Marcela Rojas Lemus, Laboratorio de Daños a la salud por metales pesados, BCyT, Facultad de Medicina, UNAM).
El incremento en la frecuencia reticulocitos micronucleados (R-MN), es una indicación de daño genético, de inestabilidad cromosómica y actualmente, tiene valor predictivo en la predisposición a desarrollar cáncer17.
CONCLUSIÓN
El material genético, independientemente del organismo del que se trate, se encuentra agrupado en una sección particular de la célula, dentro los organismo eucariontes se encuentran en una estructura muy caracterizada y compleja que le permite guardar y proteger la información genética contenida dentro del DNA. Queda claro que es más sencillo llevar a cabo los estudios a nivel molecular en organismos procariontes, porque ambos sistemas comparten similitudes: ambos requieren diversas proteínas, pero en los eucariontes el proceso es más complejo por la gran variedad de proteínas involucradas y los niveles de regulación para evitar errores en la duplicación del material.
El núcleo celular es una estructura en constante actividad. Una buena aproximación de lo que está sucediendo en la célula (o de lo que sucedió) la podemos identificar observando las características de este organelo y su contenido.
Por último, vale la pena el esfuerzo de hacer hincapié con los estudiantes, en que las estructuras y componentes que se revisan en los temarios son exactamente los mismos, porque pertenecen a las mismas células (a las que conforman su cuerpo y el de sus futuros pacientes), y la única diferencia es que son explicados, vistos y analizados a través de diferentes perspectivas. Lo importante, es que lleven a cabo esa integración con el fin de que puedan concretar la comprensión de su sujeto de estudio, el ser humano.

