











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkLas enfermedades complejas como la diabetes mellitus tipo 2, dislipidemias, obesidad, hipertensión, cardiopatía coronaria, problemas microangiopáticos, demencias, entre otras, se desarrollan por el efecto aditivo entre factores ambientales, como el estado nutricional, y factores genéticos. El elemento común es la afección simultánea de varios órganos o resistencia orgánica múltiple. En este sentido, el mapeo de genes asociados requiere considerar genes pleiotrópicos, que tienen efectos simultáneamente en diversos órganos y sistemas; uno de ellos es el gen que codifica para la altaxina-2 (ATXN2).1-2
La ataxina-2 es una proteína de unión al ARN, interactúa con PABPC1 y se localiza en los ribosomas y retículo endoplásmico rugoso. En situaciones de estrés celular, ambas proteínas son reubicadas en gránulos, donde se protegen los ARNm de la traducción y degradación. También modifica la abundancia de factores de traducción basal, la traducción global y es modulador de la biogénesis ribosomal a través de la regulación de factores de transcripción como Nop10, Rps10, rps18, RPL14, Rpl18 y Gnb2l1; así como también la iniciación de la traducción a través de los factores de Eif2s2, EIF3S6, eIF4B, Pabpc1, Srp14, βR1, SEC61B y la translocasa REr.3
El gen para la ataxina-2 es regulado por el gen A2BP1, que codifica para una proteína de unión a esta, cuya deficiencia en células hipotalámicas embrionarias de ratón ocasiona disminución de los genes atxn2, Insr y Mc4r, lo que se asocia con desarrollo de obesidad.4 Este estudio sugiere que ATXN2 está involucrado en el balance energético, a nivel hipotalámico, lo que se apoya con otros dos modelos murinos: uno que presenta deficiencia y pérdida de la función de la ataxina-2, la cual se traduce en marcada hiperfagia;5-7 el segundo, un modelo knock-out. El fenotipo clínico de estos ratones es obesidad severa, hígado graso, anormalidades del metabolismo del colesterol y disminución de la expresión del receptor de insulina en hígado y cerebro.8 El mecanismo por el cual se producen estas alteraciones todavía sigue en estudio.
En modelos animales se ha reportado que la sobreexpresión de atxn2 deriva en neurotoxicidad, en conjunto con las proteínas atxn1, atxn3, tdp-43, y mapt (también responsables de la neurodegeneración en las ataxias espinocerebelosas [SCA], esclerosis lateral amiotrófica [ELA], demencias frontotemporales y parálisis supranuclear progresiva en humanos). La atrofia neuronal se da por ganancia de la función de ATXN2, relacionada con aumento del tracto de poliglutaminas codificadas en el exón 1, lo que se explica porque la ataxina-2 interactúa con el dominio SH3 del complejo modulador de endocitosis RTK.9 Por su parte, la deficiencia de ATXN2 mitiga la neurotoxicidad y deriva en la apoptosis de líneas celulares de neuroblastoma, asociada con disminución de los niveles de Grb2 y SRC (Figura 1), lo que se ha demostrado en ratones mutantes y en cultivos de fibroblastos humanos de piel.
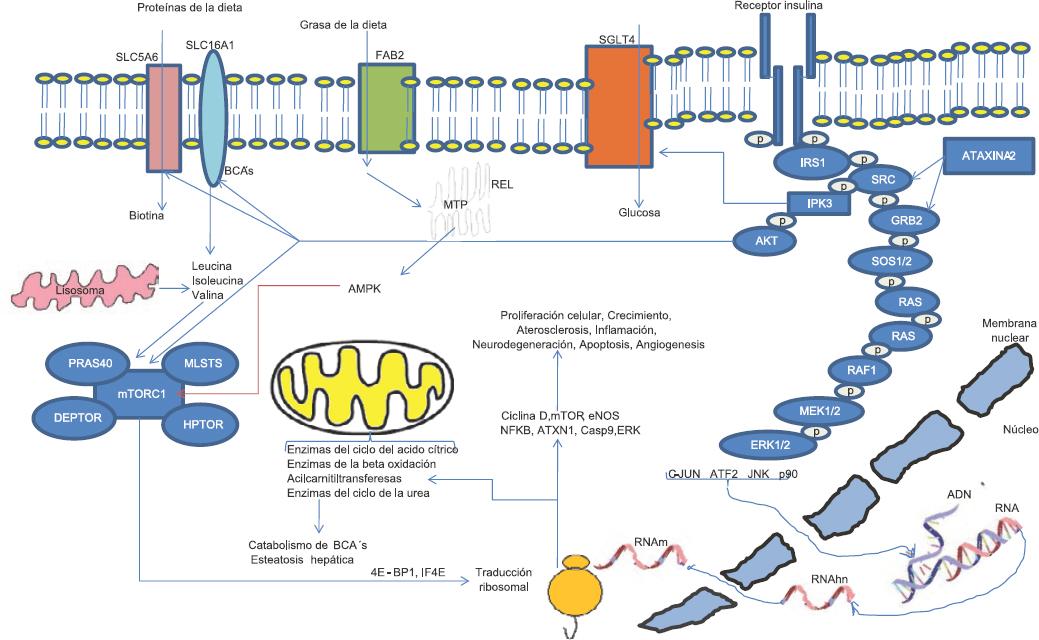
Las observaciones anteriores sugieren que ATXN2 es un modulador de la atrofia neuronal por la vía de la señalización STK; a través de AKT regula la ataxina-1, relacionada con la neurodegeneración en la ataxia espinocerebelosa tipo 1 (Figura 1).9
Los estudios de genética molecular en humanos muestran la relación directa del gen ATXN2 con el desarrollo de trastornos neurodegenerativos como ataxia espinocerebelosa tipo 2 (SCA2), ELA o síndrome pospolio, por expansión de la repetición CAG inestable en el exón 1 del gen, que codifica una poliglutamina, aunque también las interrupciones CAA en la expansión de los tripletes CAG contribuyen el fenotipo clínico de estos desórdenes. El ligamiento del locus de ATXN2, donde tiene su ubicación el VNTR, predispone al desarrollo de diabetes mellitus tipo 1 y enfermedad celiaca, lo cual también se ha relacionado con desregulación de SH2B3.10 En población inglesa se ha observado que la expansión CAG en el rango de la premutación (alelos mayores a 22 repetidos) y el estado homocigoto C para el SNP rs695872 que flanquea al VNTR están asociados con el desarrollo de obesidad severa (análoga a las mutaciones en los genes para la leptina, receptor de leptina y el receptor tipo 4 de melanocortina).5 Recientemente en un estudio de casos y controles se mostró la asociación con diabetes mellitus tipo 2 pura, con las repeticiones CAG en el rango de la premutación.11
Estudios de búsqueda exhaustiva del genoma humano han revelado la asociación del gen ATXN2 con los parámetros metabólicos e inflamatorios del síndrome metabólico,12-13 así como con parámetros hematológicos (niveles de hemoglobina, conteo y volumen de eritrocitos, leucocitos y plaquetas) en el desarrollo de enfermedad arterial coronaria e infarto al miocardio.14 En población caucásica, el locus está asociado con el calibre vascular venular retineano, cardiopatía coronaria e hipertensión.15 Un estudio epidemiológico en población europea mostró correlación con rasgos de los eritrocitos, concentración media de hemoglobina, concentración de hemoglobina, hematocrito, volumen corpuscular medio, concentración de hemoglobina corpuscular y conteo de eritrocitos.16
Las investigaciones anteriores sugieren que la ataxina-2 citoplasmática tiene un papel clave en la homeostasis metabólica en diferentes tejidos, ya que es un modulador nutricional y metabólico sensible a estrés celular. En fibroblastos embrionarios lleva al incremento de la fosforilación del factor de traducción 4E-BP1, así como de la proteína ribosomal S6, vía PI3K-mTOR.7 Por otra parte, altera el procesamiento del ARN e internalización de receptores tróficos.10
Estas observaciones se apoyan en los resultados de un modelo knock-out para ataxina-2, en el cual el hígado de los ratones presenta disminución significativa de proteínas relacionadas con la oxidación de las grasas, degradación de los aminoácidos ramificados, cetogénesis y ciclo del ácido cítrico, tales como ACADS, ALDH6A1, ALDH7A1, IVD, MCCC2, PCCA y OTC.17 Por todo ello, el gen ATXN2 en humanos, como su homólogo en ratones, participa en anormalidades relacionadas con la resistencia a la insulina, obesidad, alteraciones del metabolismo de lípidos, hipertensión, desórdenes de la homeostasis de la glucosa y polifagia severa, entre otros rasgos metabólicos inflamatorios y vasculares.18
El espectro clínico nosológico de la deficiencia del gen ATXN2 también se relaciona con problemas conductuales y obesidad, como se ha demostrado en humanos y ratones. Los ratones knock-out desarrollan reducción de la fertilidad, hiperactividad, disminución del miedo innato, deterioro de la potenciación a largo plazo de la amígdala; la plasticidad de la amígdala alterada se asocia con reducción del miedo acondicionado con claves y contextual. Este modelo animal puede ser útil para el estudio de los trastornos de ansiedad y debería fomentar la investigación de la ansiedad en pacientes con ataxia espinocerebelosa tipo 2 (SCA2).19 En humanos, los estudios de ligamiento revelan que el locus 12q24.12 del gen de la ataxina está asociado con la ganancia de peso por el consumo de antipsicóticos.5
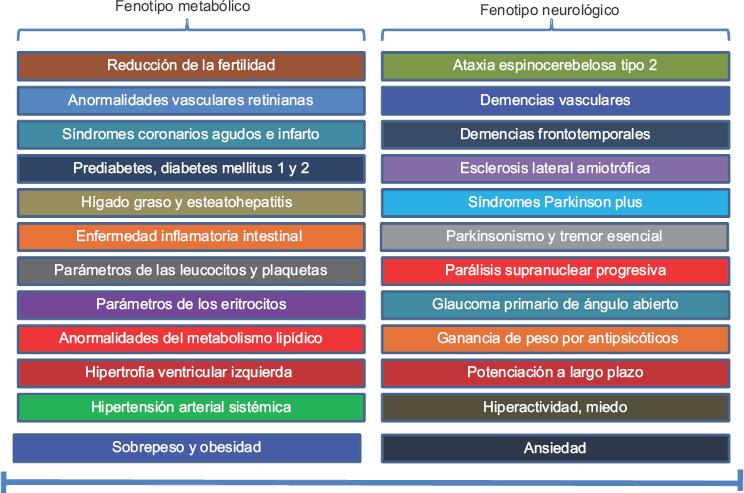
Ciertamente el espectro fenotípico clínico de la ataxina-2 es amplio (Figura 2); su variación y expresión genética están relacionadas con el desarrollo clínico de dos fenotipos: metabólico-inflamatorio-vascular y neurológico-conductual, reflejo del efecto pleiotrópico. Por todas las implicaciones fisiológicas de la ataxina-2, su gen se postula como candidato en estudios epidemiológicos de genética, epigenéticos, silenciamiento, ARN de interferencia, terapia génica, terapia antisentido para enfermedades complejas relacionadas con la nutrición, como el síndrome metabólico, trastornos neurodegenerativos y de conducta humana, que constituyen problemas de salud pública en México.
Los estudios experimentales in vitro con terapia antisentido dirigida ATXN2, parecen ser prometedores en SCA2 y ELA.20-23 Recientemente en Parkinson el blanco es la interactómica entre la ataxina-2 y EIF4G1/PARK18.23 Uno de los genes responsables de ELA, GSTO2, se ha relacionado con la edad de inicio de la SCA2, particularmente el estado heterocigoto del rs2297235.24 En un sentido similar, el locus ATXN2 es un modulador de la edad de inicio de la ataxia espinocerebelosa tipo 3.25 Estos resultados revelan que el fenotipo complejo de las enfermedades neurodegenerativas puede estar influido por la interactómica de genes y de proteínas.
En estudios con células madre se han generado varios isotipos de líneas celulares para ataxia espinocerebelosa tipo 2 IPSC H196, 221, 227 y 266, si bien continúan en investigación como opción terapéutica.24-29 Por análisis de búsqueda de larga escala genómica o de búsqueda exhaustiva del genoma con SNP se ha explorado la relación con la estructura y función cardiaca, donde el polimorfismo rs10774625 de ATXN2 se asocia con el tamaño del ventrículo izquierdo, mientras que rs7137828 se asocia con el desarrollo de glaucoma, con una razón de momios de 1.17 para el alelo T de esta variante genética.22,29 Recientes avances en el estudio del gen ATXN2 revelan que su deficiencia en el modelo knock-out provoca desregulación del metabolismo del calcio, lo cual tendrá que ser explorado en futuros estudios en pacientes con ataxia espinocerebelosa tipo 2.30
En conclusión, el gen ATXN2 puede ser usado como marcador genético en la práctica clínica como pronóstico o diagnóstico, sobre todo de enfermedades metabólicas, vasculares, inflamatorias y neurológicas, independientemente de la etiología.

