










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkGaceta médica de México
versión On-line ISSN 2696-1288versión impresa ISSN 0016-3813
Gac. Méd. Méx vol.141 no.6 Ciudad de México nov./dic. 2005
Artículo original
Efectos contráctiles dependientes del calcio intracelular inducidos por la serotonina en la arteria umbilical humana
Intracellular calcium–dependent contractile effects of serotonin in the human umbilical artery
Jesús Carlos Briones–Garduño,ª,b* Carlos Castillo–Henkel,ª Jorge Skiold López–Canales,ª y Manuel Díaz de León–Ponceb
ª Escuela Superior de Medicina, Instituto Politécnico Nacional, México, D. F. Instituto Materno Infantil del Estado de México, Toluca, Edo. de México, México
*Correspondencia y solicitud de sobretiros:
Dr. Jesús Carlos Briones–Garduño.
Paseo Colón esquina Felipe Ángeles s/n,
Toluca, Edo. de México, México. CP 50170.
Teléfono y Fax 01 722 280 92 12.
Correo electrónico: drcarlosbriones@hotmail.com
Recibido en su versión modificada: 20 de mayo de 2005
Aceptado: 27 de mayo de 2005
Resumen
Antecedentes: Los vasos umbilicales humanos se caracterizan por la ausencia de inervación y por consecuencia, las diferentes sustancias vasoactivas y la respuesta contráctil a través de los iones de calcio son los factores que determinan el flujo sanguíneo al espacio intervelloso en condiciones normales y patológicas, como es el caso de la preeclampsia eclampsia.
Objetivo: Aportar información adicional de la respuesta contráctil de la serotonina en relación al calcio intracelular en la arteria umbilical humana.
Material y métodos: Se utilizaron vasos umbilicales procedentes de mujeres con embarazo normoevolutivo, los cuales una vez disecados se cortaron en anillos de 5 mm y se montaron en cámara de órgano aislado, utilizando solución de Krebs con y sin calcio, burbujeada con carbógeno y la temperatura controlada. Se evaluó el efecto contráctil inducido por serotonina a diferentes concentraciones molares y se contrastó con verapamil, lantano y ácido ciclopiazónico, cuantificando la respuesta contráctil mediante un polígrafo biopac acoplado a un sistema computacional.
Resultados: No se observaron diferencias significativas en la magnitud de la respuesta obtenida en presencia y ausencia de calcio extracelular. Se apreció el efecto contráctil a la serotonina que disminuyó significati vamente como respuesta a la estimulación repetida a la misma; así mismo, se incrementó el tono basal posterior a la adición del calcio al medio de incubación, lo que dependió del tiempo de exposición. También se observó la inhibición parcial del incremento en el tono basal con vera pamil y lantano. Finalmente, el pretratamiento con ácido ciclopiazónico no modificó la respuesta contráctil a la serotonina en un medio sin calcio.
Conclusiones: La contracción inducida por serotonina en la arteria umbilical humana, depende principalmente de calcio intracelular y favorece el ingreso capacitativo de este ion, el cual se incrementa gradualmente a través del tiempo. El ingreso capacitativo del calcio secundario al vaciamiento de los depósitos intracelulares de este ion con serotonina se efectúa a través de canales tipo L y no–L, y no parecen ser sensibles al ácido ciclopiazónico.
Palabras clave: Arteria umbilical, serotonina, respuesta contráctil, depósitos intracelulares de calcio.
Summary
Background: Absence of innervation is a hallmark of human umbilical vessels. Intervillous space blood flow is regulated by vasoactive substances and calcium dependent contractility, both in normal and pathological conditions such as preeclampsia eclampsia.
Objective: To obtain additional information on the intracellular calcium contractile effects of serotonin in human umbilical arteries.
Materials and Methods: Umbilical arteries from normal pregnancies were dissected, cut in 5 mm rings and mounted in a temperature controlled isolated organ chamber, using calciumfree Krebs solution. The contractile effects of serotonin, lantane, verapamil and cyclopiazonic acid were evaluated at different concentrations using a computer coupled biopac polygraph.
Results: No differences in response were observed in the presence and absence of intracellular calcium. The positive contractile effects observed with serotonin were significantly decreased with repeated stimulation. An increase in the basal tone of the vessel was observed after calcium supplementation was added to the solution. This effect was minimized in the presence of verapamil and lantane. The contractile effects of serotonin in the calcium free solution were not affected by the presence of cyclopiazonic acid.
Conclusions: Serotonin contractile effects in the human umbilical artery depend mainly on intracellular calcium levels which favor the gradual entrance of this ion over time. Calcium influx induced by serotonin is possible through L and Non–L channels apparently insensitive to cyclopiozonic acid.
Key words: Umbilical artery, serotonin, contractile response, intracellular calcium.
Introducción
Durante el embarazo normal la paciente se encuentra vasodilatada como un mecanismo fisiológico que permite tolerar los principales cambios hemodinámicos como son el incremento en el volumen circulante y consecuentemente en el gasto cardiaco, lo que asegura no incrementar su presión arterial. Sin embargo, en algunos procesos patológicos como en la preeclampsia–eclampsia se ha demostrado que existen alteraciones en el equilibrio entre estímulos vasopresores y vasodilatadores que puedan explicar el incremento en las resistencias vasculares (hipertensión arterial) y por consecuencia la disminución en el flujo placentario. Cabe señalar que este concepto es básico si recordamos que los vasos placentarios humanos carecen de inervación autónoma y por lo tanto, la resistencia vascular a este nivel dependerá de estos factores humorales.1 La placenta humana (hemocorial) se caracteriza por el intercambio de nutrientes entre madre y feto a través de la barrera placentaria (trofoblasto y endotelio vascular). Por sus características anatómicas (arterias espirales), la eyección al espacio intervelloso es con alta presión y turbulencia; el mecanismo compensador es la vasodilatación propiciada por la invasión trofoblástica que incursiona hasta la capa muscular del vaso arterial, disminuyendo la resistencia vascular así como el impacto del chorro de sangre al espacio intervelloso (cambio que en la paciente con preeclampsia no se logra). Es probable que en este mecanismo estén implicadas variables inmunológicas y genéticas relacionadas a antígenos de histocompatibilidad (HLA) hasta ahora poco explorados, lo que conduce a isquemia e hipoxia utero–placentaria responsable de la disminución en la síntesis de prostaciclina (PGI2) como principal agente vasodilatador y antiagregante plaquetario. En condiciones normales este efecto vasodilatador es más evidente durante el primer trimestre del embarazo (de gran actividad trofoblástica). La isquemia uteroplacentaria origina cuatro efectos principales:
1) Incremento en la peroxidación de lípidos.
2) Deportación de trofoblasto.
3)Activación de la xantina–oxidasa favoreciendo la producción de ácido úrico.
4) Activación de neutrófilos y plaquetas que producen principalmente radicales libres de oxígeno, lo que en conjunto causa daño endotelial multiorgánico de magnitud muy variable.
Clínicamente, lo que la mujer con preeclampsia presenta es el resultado de estos cambios, identificados con tres características clínicas principales:
a) Aumento de la permeabilidad capilar que reconocemos como síndrome de fuga capilar, causando hipovolemia y disminución de la presión coloidosmótica (albuminuria); esto explica el edema somático y visceral que se observa frecuentemente en cara, pies, manos, cerebro, pulmón, hígado, etc.
b) Aumento del tono vascular que se reconoce por la hipertensión que junto con la hipovolemia y la disminución de la presión coloidosmótica conduce a un estado hiperdinámico compensatorio que se presenta con diversos patrones clínicos.
c) Compromiso microcirculatorio y hemorreológico demostrado por deportación de trofoblasto, depósito de micro–agregados, material fibrinoide y agregación plaquetaria, que clínicamente referimos como anemia hemolítica microangiopática, y su variante más frecuente caracterizada por hemólisis, elevación de enzimas hepáticas y plaquetopenia (síndrome de HELLP), comprometiendo la función renal y hepática principalmente (insuficiencia renal aguda y necrosis periportal) o dando paso al síndrome intermedio de coagulación intravascular diseminada (CID) en los casos donde en forma simultánea hay trastornos de los mecanismos de la coagulación y de la fibrinólisis. Los datos clínicos que cada paciente presenta en forma individual dependerán de su reserva orgánica y de lo oportuno del diagnóstico y del tratamiento que requieren estas mujeres críticamente comprometidas.2
En relación al aumento en el tono vascular, la contracción del músculo liso depende de la disponibilidad de calcio libre intracelular,3 el cual puede ser liberado desde el retículo sarcoplásmico o entrar a la célula a través de canales de calcio dependientes de voltaje u operados por receptor.4,5 La fuente de calcio puede variar dependiendo del agente vasoconstrictor y del lecho vascular. Como sucede con las catecolaminas o con la angiotensina II, la serotonina (5–HT) debe su actividad vasoconstrictora a la activación de receptores (del tipo 5–HT2) acoplados a proteínas Gq.6,7 La vasoconstricción asociada a la activación de las proteínas Gq, se relaciona en general con incremento en la disponibilidad de Ca++ libre intracelular que procede tanto de la liberación de depósitos intracelulares como del ingreso desde el espacio extracelular.5,8,9 No obstante que existe evidencia de que lo anterior es cierto, en la mayoría de los vasos sanguíneos estudiados, la información acerca del comportamiento en los vasos umbilicales humanos en este sentido es escasa; es decir, está bien establecido que la 5–HT es uno de los pocos agentes capaces de contraer vasos umbilicales, pero la relación que dicha contracción tiene con los depósitos intra y extracelulares de calcio en las células de músculo liso de dichos vasos no se ha definido con claridad. Por lo anterior, el presente trabajo intenta aportar información adicional al respecto, ya que esta patología (preeclampsia–eclampsia) continúa siendo la primera causa de mortalidad materna en nuestro país25 y dentro de su fisiopatogenia la reactividad vascular incrementada que determina el factor principal de la hipertensión arterial2 representa un campo abierto para la búsqueda de fármacos con propiedades suficientes y adecuadas para el tratamiento antihipertensivo y simultáneamente para mejorar la perfusión intervellosa (uteroplacentaria) que favorezca el desarrollo del producto de la gestación.
Material y métodos
Se utilizaron cordones umbilicales procedentes de 15 mujeres jóvenes que se atendieron en la Unidad de Tococirugía del Hospital de Ginecología y Obstetricia del Instituto Materno Infantil del Estado de México. Al momento del nacimiento (parto o cesárea) se obtuvieron de 10 a 15 cm, de tercio medio de cordón umbilical, los cuales se colocaron en solución de Krebs burbujeada con carbógeno, para ser transportados después en un frasco hermético y dentro de un contenedor de unicel mediante geles congelantes, hasta el laboratorio de farmacología en donde se procedió a disecar la arteria umbilical retirándole todo el tejido adyacente. La arteria se cortó en anillos de aproximadamente 5 mm de longitud, cada uno de los cuales se montó en dos ganchos de acero inoxidable colocándolos después en cámaras para órgano aislado de 10 mL llenadas con solución de Krebs–bicarbonato de la siguiente composición (mM): NaCl, 118; KCl, 4.7; KH2 PO4, 1.2; MgSO4, 1.2; CaCl2, 2.5; NaHCO3, 25; dextrosa, 11.7; y EDTA disódico cálcico, 0.026, burbujeada con carbógeno, con temperatura de 37° y pH 7.4. La solución libre de Ca2+ tenía la misma composición salvo que el CaCl2 fue omitido y se añadió EDTA (2 mM). La solución en el baño se mantuvo a 37°C, pH 7.4 y se burbujeó con carbógeno (95% de O2 y 5% CO2). Cada anillo se fijó por un extremo al fondo de la cámara y por otro a un transductor de tensión, lo que permitió registrar los cambios de tensión en la preparación. Se les dio una tensión inicial de 2 gramos a los anillos y se les dejó estabilizar por dos horas antes de iniciar el experimento.
Efecto contráctil de la serotonina en presencia o ausencia de calcio extracelular
Con el objeto de establecer la participación del calcio extra–celular en el efecto contráctil de la 5–HT, se compararon los efectos producidos por la concentración submáxima (10–5 M) de esta amina en anillos de arteria umbilical incubados en presencia o ausencia de calcio en el medio de incubación.
Efectos de la administración repetida de serotonina
Después del período de estabilización se administró una concentración submáxima de serotonina (10–5 M) para contraer los anillos arteriales incubados en solución libre de Ca2+; a continuación las preparaciones arteriales se lavaron y se repitió la administración de 5–HT dos veces más, al final de la cual se añadió Ca2+ (2.5 mM) al medio. La intención era el establecer si como sucede con otros vasos sanguíneos y con otros agentes contráctiles, la administración repetida de 5–HT conduce a vaciamiento de los depósitos intracelulares de Ca2+ y esto a su vez da lugar al denominado "ingreso capacitativo" de Ca2+ desde el espacio extracelular cuando este ion es añadido al medio de incubación, ocasionando incremento en el tono basal.10,11 Cuando se requirió y una vez que se observó el incremento en tono basal debido a la adición de calcio, los anillos se lavaron con solución conteniendo Ca2+, permaneciendo en ella por una hora (período de reequilibrio) antes del estudio siguiente.
Curso temporal de la respuesta a la adición de calcio
Con la intención de establecer si la incubación en un medio sin calcio daba lugar a la apertura gradual de canales de Ca2+ que permitiera el ingreso del ion cuando se añade al medio de incubación, se evaluó el incremento en el tono basal asociado con la adición de calcio a los 15, 30, 45 y 60 minutos de exponer las preparaciones arteriales a una solución libre de calcio.
Efecto del verapamil y el lantano en el incremento del tono basal ocasionado por la adición de calcio
Con el objeto de establecer si el incremento en el tono basal ocasionado al administrar calcio al medio de incubación de los anillos pretratados con 5–HT, ocurría a través de canales de calcio tipo L o tipo no–L, se recurrió al verapamil y al lantano respectivamente. Estos agentes se administraron 15 min. antes de reincorporar calcio al medio de incubación.
Análisis de la capacidad del ácido ciclopiazónico para inducir la contracción de anillos de arteria umbilical
El ácido ciclopiazónico depleta los depósitos intracelulares de calcio al inhibir su captura por el retículo sarcoplásmico a través de la inhibición de la Ca2+ ATPasa. La intención de utilizar este fármaco fue establecer si los depósitos intracelulares de calcio sobre los que actúa la serotonina eran sensibles al ácido ciclopiazónico y si como sucede en otros vasos sanguíneos, este compuesto provoca en las arterias umbilicales incubadas en un medio con calcio a contracción por ingreso capacitativo de este ion.
Resultados
De las principales variables clínicas de la población de mujeres estudiadas, destacaron: edad de 24.2 ± 6.1, semanas de gestación de 36.7 ± 4.0, peso de los recién nacidos de 2877.6 ± 633.9 g, primigestas 53% y la resolución obstétrica por la vía vaginal mediante parto eutócico, 67%.
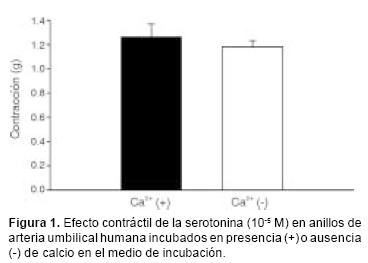
La 5–HT (10–9 – 10–4 M) ocasionó contracción dependiente de la concentración en los anillos de arteria umbilical humana con un efecto máximo de 1.42 ± 0.12 g. La concentración de 10–5M ocasionó aproximadamente 90% de la respuesta máxima; esa concentración submáxima es la que se usó en el desarrollo del presente estudio. En la figura 1 se presentan los resultados obtenidos al administrar la concentración submáxima de 5–HT en anillos de arteria umbilical incubados en un medio con o sin calcio. No se encontraron diferencias significativas en la magnitud de la respuesta en presencia y ausencia de calcio extracelular, siendo los efectos máximos de 1.27 ± 0.10 y de 1.82 ± 0.5 g, respectivamente.
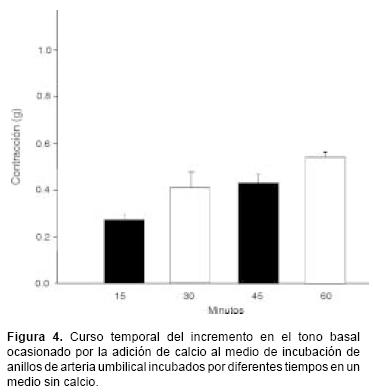
En la figura 2 se muestran los resultados obtenidos al administrar consecutivamente en tres ocasiones 5–HT 10–5M (con lavado después de cada administración) en anillos de arteria umbilical incubada en un medio sin calcio y al añadir calcio (2.5 mM) después de la última administración de la amina. Se encontró que el efecto contráctil de la 5–HT fue significativamente menor al repetir su administración, tendiendo prácticamente a desaparecer a la tercera aplicación. Por otra parte, se observó que la adición del calcio al medio de incubación después de la administración repetida de 5–HT condujo a un incremento en el tono basal. Dicho aumento fue de magnitud muy superior (Figura 3) en comparación con el que espontáneamente se presentó en respuesta a la adición de calcio en anillos control que fueron incubados en un medio sin calcio, pero sin ser expuestos a la serotonina. Este incremento espontáneo en el tono basal (Figura 4) se presentó gradualmente con el tiempo de exposición al medio sin calcio pero siempre fue inferior al que se presentó después de pretratar los anillos de la arteria umbilical con serotonina (Figura 3).


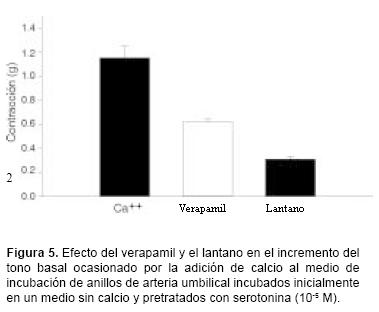
En la figura 5 se muestran los resultados obtenidos al añadir calcio al medio de incubación, después de pretratar los anillos de la arteria umbilical con 5–HT, en ausencia o presencia de verapamil o de lantano. Se aprecia que tanto el verapamil como el lantano provocaron inhibición parcial del incremento en el tono basal ocasionado por la adición de calcio al medio de incubación. La inhibición provocada por el lantano fue superior a la producida por el verapamil.
Finalmente, en la figura 6 se aprecia que el pretratamiento con el ácido ciclopiazónico no modificó significativamente la respuesta contráctil a la 5–HT en anillos incubados en un medio sin calcio. El ácido ciclopiazónico tampoco fue capaz de inducir contracción en anillos arteriales incubados en un medio con calcio.

Discusión
La ausencia de diferencias significativas al comparar la contracción inducida por una concentración submáxima de la 5–HT en anillos de arteria umbilical incubados en un medio en presencia o ausencia de calcio, sugiere que el efecto contráctil de la serotonina en este vaso es dependiente esencialmente del calcio intracelular. La 5–HT, al igual que otros agonistas de receptores acoplados a proteínas Gq, puede al unirse a su receptor, y como resultado de la activación de la proteína Gq estimular a la fosfolipasa C, enzima que hidroliza al fosfolípido de membrana fosfatidilinositol 4,5–bifosfato dando lugar a los dos segundos mensajeros inositol 1,4,5 trifosfato (IP3) y diacilglicerol.12–14 El IP3 se une a receptores localizados en el retículo sarcoplásmico y libera Ca2+ almacenado,14,15el cual es probablemente responsable de la respuesta contráctil a la 5–HT observada en los anillos de arteria umbilical incubados en solución libre de Ca2+.
Se ha establecido que la administración de vasoconstrictores que son agonistas de receptores acoplados a proteínas Gq, conduce a vaciamiento de los depósitos intracelulares de Ca2+ y como consecuencia de este fenómeno se abren canales de Ca2+ en la membrana celular, lo que permite el ingreso de Ca2+ extracelular que incrementa el tono basal.11 En consecuencia, los depósitos intracelulares de Ca2+ desempeñan un papel crucial en la respuesta contráctil de las células de músculo liso vascular, puesto que los depósitos vacíos de Ca2+ son la señal para el denominado ingreso capacitativo u operado por depósitos de Ca2+ extracelular16,17 y se ha demostrado que dependiendo del vaso estudiado, los canales de Ca2+ dependientes de voltaje están involucrados en este proceso.18–23 Los resultados del presente estudio indican que el incremento en el tono basal en las arterias umbilicales en respuesta al vaciamiento de los depósitos intracelulares de Ca2+ con serotonina, se debe al influjo capacitativo de Ca2+ a través de canales de Ca2+ tanto de tipo L sensibles a verapamil como de tipo no–L, sensibles a lantano.
Llama la atención que a diferencia de otros vasos sanguíneos, en las arterias umbilicales la incubación en un medio sin calcio por un tiempo inferior a los 70 min dio lugar al ingreso del calcio al interior de las células cuando se incorporó este ion al medio de incubación. La magnitud del ingreso se incrementó gradualmente con el tiempo de exposición al medio sin calcio, pero en todo caso fue inferior al que se produjo cuando las preparaciones arteriales fueron pretratadas con serotonina. Es decir, el ingreso capacitativo ocasionado por el vaciamiento de los depósitos intracelulares con el agonista contráctil fue superior al que se presentó espontáneamente por la exposición a un medio sin calcio.
También es interesante el hecho de que los depósitos intracelulares de Ca++ de las arterias umbilicales parecen ser insensibles al efecto del ácido ciclopiazónico. Este compuesto ocasiona vaciamiento de los depósitos intracelulares de calcio al inhibir la Ca2+–ATPasa y consecuentemente la captación de calcio por el retículo sarcoplásmico.24 A diferencia de lo que ocurre en otros vasos, el pretratamiento de las arterias umbilicales incubadas en un medio sin calcio con el ácido ciclopiazónico no inhibió el efecto contráctil de la serotonina, indicando que dicho compuesto no vació los depósitos intracelulares de calcio. Congruente con lo anterior, el ácido ciclopiazónico no ocasionó contracción de las arterias umbilicales incubadas en un medio con calcio, indicando que al no vaciar los depósitos intracelulares de éste no se presenta el ingreso capacitativo de este ion que conduce a la contracción.11
En resumen, la contracción inducida por la 5–HT en las arterias umbilicales humanas depende principalmente del calcio intracelular. La 5–HT ocasiona vaciamiento de los depósitos intracelulares de calcio y en consecuencia favorece el ingreso capacitativo de este ion, lo cual se traduce en el incremento en el tono basal observado al reincorporar calcio al medio de incubación.
La sola exposición a un medio sin calcio favorece, al añadir este ion, su ingreso al músculo liso de arterias umbilicales, el cual se incrementa gradualmente a través del tiempo. Sin embargo, la magnitud de este ingreso parece ser menor al que se produce como consecuencia del pretratamiento de las preparaciones arteriales con 5–HT. El ingreso capacitativo de calcio consecutivo al vaciamiento de los depósitos intracelulares de este ion con 5–HT, parece efectuarse en las arterias umbilicales a través de canales de calcio tipo L y de tipo no–L. Los depósitos intracelulares de calcio de las arterias umbilicales no parecen ser sensibles al ácido ciclopiazónico.
Referencias
1. Abad A, Estaño L, Morales FJ, Serra V. Estudio de la contractilidad de 14. los vasos coriónicos humanos in vitro en la preeclampsia. Prog Obstet Ginecol 2000; 43:177–181. [ Links ]
2. Briones GJC, Díaz de León PM, Gómez Bravo TE, Ávila EF, Briones VCG, Urrutia TF. Protocolo de manejo en la preeclampsia–eclampsia. Cirugía y Cirujanos 1999; 67:6. [ Links ]
3. Walsh MP. Regulation of vascular smooth muscle tone. Can J Physiol Pharmacol 1993; 72:919–936. [ Links ]
4. Minneman, KP, Esbenshade TA. a1 adrenergic receptor subtypes. Annu Rev Pharmacol Toxicol 1994; 34:117–133. [ Links ]
5. Martin GR. Vascular receptors for 5–hydroxytryptamine: distribution, function and classification. Pharmac Ther 1994; 62:283–324. [ Links ]
6. Fain JR, García–Sainz JA. Role of phosphatidylinositol turnover in a, and of adenylate cyclase inhibition in a2 effects of catecholamines. Life Sci 1980; 26:1183–1194. [ Links ]
7. García–Sainz JA, Vázquez–Prado J, Villalobos–Molina R. c^–adrenoceptors: subtypes, signaling, and roles in health and disease. Arch Med Res 1999; 30:449–458. [ Links ]
8. Timmermans PB, Wong PC, Chiu AT, Herblin WF, Benfield P, Carini DJ, et al. Angiotensin II receptors and angiotensin II receptor antagonists. Pharmacol 21. Rev 1993; 45:205–251. [ Links ]
9. Suematsu E, Hirata M, Sasaguri T, Hashimoto T, Kuriyama H. Roles of Ca2+ on the inositol 1,4,5–triphosphate–induced release of Ca2+ from saponin– permeabilized single cells of the porcine coronary artery. Comp Biochem Physiol 1985; 82:645–649. [ Links ]
10. Noguera MA, Ivorra MD, Chulia S, D'Ocon M P. Capacitative Ca2+ entry associated with a1–adrenoceptors in rat aorta. Naunyn Schmiedeberg's Arch 23. Pharmacol 1996;355:1–7. [ Links ]
11. Noguera MA, Madrero Y, Ivorra MD, D'Ocon P. Characterization of two different Ca2+ entry pathways dependent on depletion of internal Ca2+ pools in rat aorta. Naunyn–Schmiedeberg's Arch Pharmacol 1998; 357:92–99. [ Links ]
12. Terzic A, Puceat M, Vassort G, Vogel SM. Cardiac c^–adrenoceptors: an overview. Pharmacol Rev 1993; 45:147–175. [ Links ]
13. Graham RM, Perez DM, Hwa J, Piascik MT. a,–Adrenergic receptor subtypes. Molecular structure, function and signaling. Circ Res 1996; 78:737–49. [ Links ]
14. Varma DR, Deng X–F. Cardiovascular a,–adrenoceptor subtypes: functions and signaling. Can J Physiol Pharmacol 2000; 78:267–292. [ Links ]
15. Karaki H, Ozaki H, Hori M, Mitsui–Saito M, Amano K, Harada K, et al. Calcium movements, distribution, and functions in smooth muscle. Pharmacol Rev 1997; 49(No 2):157–230. [ Links ]
16. Putney JW. A model for receptor–regulated calcium entry. Cell Calcium 1986; 7: 1–12. [ Links ]
17. Putney JW. Capacitative calcium.entry revisited . Cell Calcium 1990; 11:611–634. [ Links ]
18. Felder CC, Singer–Lahat D, Mathes C. Voltage–independent calcium channels. Regulation by receptors and intracellular calcium stores. Biochem Pharmacol 1995; 48:1997–2004. [ Links ]
19. Dessy C, Godfraind T. The effect of L–type calcium channel modulators on the mobilization of intracellular claim stores in guinea–pig intestinal smooth muscle. Br J Pharmacol 1996; 119:142–148. [ Links ]
20. Asano M, Kuwako M, Nomura Y, Ito KM, Ito K, Uyama Y, et al. Possible mechanism of the potent vasoconstrictor actions of ryanodine on femoral arteries from spontaneously hypertensive rats. Br J Pharmacol 1996; 118:1019–27. [ Links ]
21. Noguera MA, D'Ocon MP. Evidence that depletion of internal calcium stores sensitive to noradrenaline elicits a contractile response dependent on extracellular calcium in rat aorta. Br J Pharmacol 1993; 110: 861–7. [ Links ]
22. Sekiguchi F, Shimamura K, Akashi M, Sunano S. Effects of cyclopiazonic acid and thapsigargin on electromechanical activities and intracellular Ca2+ in smooth muscle of carotid artery of hypertensive rats. Br J Pharmacol 1996; 118: 857–864. [ Links ]
23. Fasolato C, Innocenti B, Pozzan T. Receptor–activated Ca2+influx: how many mechanisms for how many channels? Trends Pharmacol Sci 1994; 15:77–83. [ Links ]
24. Okoro EO. Overlap in the pharmacology of L–type Ca2+channel blockers and 5– HT2 receptor antagonists in rat aorta. J Pharm Pharmacol 1999; 51 (8):953–957. [ Links ]
25. Gómez Bravo TE, Briones GJC, Morales EJG, Rodríguez RM. Mortalidad materna en el Estado de México. Rev Asoc Mex Med Crit y Ter Int 2003; 17(6): 199–203. [ Links ]

