










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
versión On-line ISSN 2448-6698versión impresa ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.1 no.3 Mérida jul./sep. 2010
Artículos
Efecto de una subalimentación prolongada sobre el peso, la condición y la composición corporal de cabras adultas
Effect of a long term feeding restriction on the subsequent body weight, condition score, and tissue composition of mature goats
Manuel Gómez Pasténa, Ofelia Mora Izaguirreb, Rosa María Meléndez Sotoa, José Luis Romano Muñozc, Héctor Vera Avilac, Armando Shimada Miyasakab,c
a Programa de Maestría y Doctorado en Ciencias de la Producción y la Salud Animal, Universidad Nacional Autónoma de México (UNAM).
b Laboratorio de Rumiología y Metabolismo Nutricional (RuMeN), Facultad de Estudios Superiores-Cuautitlán. UNAM., Blvrd. Juriquilla No 3001, Querétaro, Qro., 76230. shimada@unam.mx. Correspondencia al último autor.
c Centro Nacional de Investigación en Fisiología Animal, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP).
Recibido el 26 de noviembre de 2008.
Aceptado para su publicación el 11 de marzo de 2010.
RESUMEN
Para determinar la respuesta de cabras adultas a una restricción alimenticia prolongada y el efecto en su peso, condición y composición corporal, se llevó a cabo un experimento usando 21 cabras hembras encastadas de Nubia, adultas, vacías y secas. Se registró durante nueve semanas el peso, condición corporal y consumo diario. Después se dividieron completamente al azar en tres grupos, para recibir durante 36 semanas los siguientes niveles de alimentación (NA): 100, 80 y 60, como porcentaje del consumo observado previamente. Se analizó químicamente el tejido disectable, corazón, riñón e intestinos. El peso y la condición corporal disminuyeron con la restricción alimenticia y se aumentó la proporción de hueso, se redujo la de tejido disectable (media canal derecha), grasa disectable (TA), grasa peri-renal e hígado, contenido total de materia seca de corazón, riñón y líquido ruminal; proteína del riñón y extracto etéreo del corazón y tejido disectable de la canal; concentración de ADN del corazón, hígado y TA visceral y relación ARN: ADN y proteína: ADN del hígado. En cuanto a la composición del TA visceral, el único ácido graso afectado fue C16:1, teniendo los animales con nivel de NA60 mayor proporción; pero en el TA subcutáneo, la restricción aumentó la proporción del C16:1, C18:1 y de ácidos grasos insaturados y redujo la proporción de los ácidos grasos saturados. Los resultados muestran la capacidad de adaptación de las cabras adultas a una malnutrición a largo plazo, mediante la utilización de combustibles metabólicos provenientes de tejido adiposo, músculo esquelético, hígado, corazón y riñón.
Palabras clave: Condición corporal, Peso corporal, Composición química, Acidos grasos, Cabras.
ABSTRACT
To observe the response of mature goats to long-term feeding restriction and the effect on their body weight, condition and composition, an experiment was conducted with 21 adult, female, non-gestating, non-lactating Nubian goats. For a 9-wk stabilization period, the body weight and body condition score of each animal were measured weekly, along with their voluntary feed intakes. After the stabilization period, goats were randomly assigned to three groups, being offered 100%, 80%, and 60% of the feed intake observed during this period, for a 36-wk restriction period. Chemical analyses were performed on samples of carcass components, heart, kidneys, and intestines. Body weight and body condition score decreased with reduced feeding levels. Moreover, the proportion of bone increased, and that of the carcass soft tissues, kidney fat, and liver decreased, as did the dry matter contents of heart, kidneys, and rumen contents; kidney protein, heart, and ether extracts of the carcass soft tissue; heart and liver DNA; and the RNA:DNA and protein:DNA ratios in hepatic tissue. The C16:1 fatty acid content of the kidney fat of goats on treatment FL60 was increased; in restricted animals, C16:1, C18:1, and unsaturated fatty acids in the subcutaneous fat were all increased. These results suggest that the adaptation capability of adult goats to long-term undernutrition through the utilization of metabolic fuel from adipose tissue, skeletal muscle, liver, heart, and kidneys is an important survival characteristic for free-ranging animals kept in areas where drought conditions can last for several months, with consequent limitations on feed availability.
Key words: Body condition, Body weight, Chemical composition, Fatty acids, Goats.
INTRODUCCIÓN
Las necesidades nutritivas de los rumiantes en pastoreo se cubren mediante el consumo de alimentos, sean éstos los disponibles en el agostadero o aquéllos que les son ofrecidos en forma complementaria por el productor. Cuando el aporte de tales alimentos es insuficiente para cubrir sus necesidades básicas, hacen uso de reservas corporales, manifestándose lo anterior como pérdidas de peso, cuya magnitud dependerá de la severidad y la duración de la insuficiencia alimenticia. Las reservas corporales así utilizadas podrán ser subsecuentemente recuperadas(1,2,3,4).
Varios reportes indican que después de períodos de pérdida de peso, los animales reducen sus requerimientos para mantenimiento hasta en un 50%, principalmente mediante la reducción de la tasa metabólica basal(5) y también mediante la reducción en la masa de algunos órganos metabólicamente activos como hígado, riñón, estómago e intestinos(6). Como lo indican la reducción en la relaciones proteína:DNA y RNA:DNA, que sugieren que una privación de nutrimentos reduce la concentración de RNA y de proteínas de las vísceras debido a una disminución aparente del tamaño celular y en la actividad de síntesis de proteína(7,8).
El tejido adiposo blanco está distribuido en diversos tejidos corporales, principalmente en las zonas dérmica, subcutánea, mediastinal, mesenterial, perigonadal, omental, renal, y retropiritoneal(9) y es sabido que aún en el mismo animal, la composición de la grasa subcutánea y el perfil de ácidos grasos contenidos en la misma, no es constante en las diferentes zonas del cuerpo, el grado de instauración depende de la localización anatómica. En general, la grasa subcutánea es la más insaturada, seguida por la grasa intermuscular e intramuscular, mientras que la grasa de los órganos internos es la más saturada(10).
El propósito de este experimento, fue observar en cabras los efectos de una subalimentación prolongada, especialmente en su peso, condición y composición corporal. Además de conocer cómo y en qué medida son utilizados los diferentes órganos y tejidos, para compensar la restricción de nutrimentos.
MATERIALES Y MÉTODOS
El trabajo fue conducido en el estado de Querétaro, a 1,990 msnm, clima semiseco con lluvias en verano, temperatura media anual de 15 °C y precipitación pluvial anual de 450 a 630 mm, principalmente en el verano(11).
Un grupo de 21 cabras hembras, adultas, vacías y secas, con predominancia de sangre Nubia, con un peso corporal inicial de 50.1 ± 5.8 kg y una calificación de condición corporal (CC) de 2.6 ± 0.5 fueron utilizadas. Las cabras permanecieron alojadas en corraletas individuales techadas, con piso de madera, con acceso a bebederos automáticos y comederos individuales.
Al inicio del experimento, todos los animales se alimentaron a libertad durante nueve semanas con una dieta con base en pasta de sorgo (50%), heno de alfalfa (45%), melaza de caña (3.5%) y premezcla mineral-vitamínica (1.5%), de tal manera que cubrieran sus necesidades teóricas de energía metabolizable (EM) y proteína cruda de mantenimiento (PC)(12); nueve semanas de período de estabilización se consideraron suficientes para alcanzar condición corporal y peso constantes(13), y para ajustar la cantidad de alimento según los requerimientos de mantenimiento. El contenido de PC se determinó por el método de Kjeldahl y la EM fue estimada de tablas del NRC(12).
Una vez terminadas las nueve semanas, las cabras se distribuyeron aleatoriamente en cada uno de los siguientes niveles de alimentación que recibieron durante 36 semanas: NA1=100% del consumo observado de alimento; NA2= 80% del consumo observado de alimento; NA3=60% del consumo observado de alimento. Los animales fueron alimentados en forma individual dos veces al día (0900 y 1300).
Los animales se pesaron y su condición corporal se observó(14) al inicio y cada 14 días en días consecutivos, sin ayuno previo y siempre a la misma hora. Empleando los datos de peso corporal, el peso metabólico (W.75) se calculó cada dos semanas y la cantidad de alimento ofrecido fue ajustada; en otras palabras el alimento ofrecido se redujo periódicamente, en respuesta al peso corporal perdido.
Al término del periodo de restricción (PR), las cabras fueron pesadas y sacrificadas por el método del émbolo cautivo, y desangradas. Se registraron los pesos de piel, patas, cuernos, cabeza, pulmones, útero, hígado, riñón, corazón, intestino delgado, colon, compartimentos gástricos y contenido retículo-ruminal. Se determinó la longitud de los intestinos; y se tomaron muestras de órganos, que se mantuvieron en congelación (-70 °C) para análisis de MS (materia seca)(15), EE (extracto etéreo), proteína total(16); para determinar ARN total y ADN genómico, se emplearon reactivos de TRIzol and DNAzol®, respectivamente (Invitrogen®, Carisbad, CA, USA); la extracción de ADN consiste básicamente en el uso de una solución lisante de detergente de guanidina, que permite la precipitación selectiva del ADN del lisado celular. En el caso del ARN se emplea el mismo reactivo, luego se añade cloroformo, que separa la solución en una fase acuosa y una orgánica.
Se registró el peso de la canal caliente y se realizó un corte longitudinal de la misma, evitando la columna vertebral, resultando una media canal izquierda con la totalidad de la columna vertebral y una media canal derecha sin la misma. De la media canal derecha se registraron los pesos total, del hueso y del tejido disectable, y se muestrearon para su análisis químico.
Se registró el peso de la grasa disectable y la perirenal (riñón derecho). Se obtuvieron muestras de tejido adiposo peri-renal y subcutáneo (esternal) para determinación de DNA y perfil de ácidos grasos (PAG); las muestras fueron hidrolizadas con una solución al 20% KOH en metanol a 80 °C, durante 40 min, para realizar la extracción de la fracción lipídica con acetonitrilo/tetrahidrofurano (ACN/THF). Los ácidos grasos se cuantificaron con cromatografía de líquidos de alta resolución (Hewlett Packard 1100 System con detector de diodos), con un programa de derivatización en línea con bromuro de bromofenacilo. Se usó una columna MOS de 150 x 4.6 mm, 5 |m, con una fase móvil, en gradiente, a base de agua (70%) y ACN adicionado de 1% THF (30%).
El análisis estadístico se realizó mediante un diseño completamente al azar(17), con tres tratamientos: NA100, NA80 y NA60 con 8, 7, y 6 repeticiones por tratamiento, respectivamente. Para determinar diferencias entre tratamientos se utilizó el procedimiento de modelos lineales generales (GLM) del Sistema de Análisis Estadístico(18) y la opción LSMEANS se empleó para comparar las medias de tratamientos.
Las medidas tomadas a partir de la canal y los pesos de las vísceras se registraron como peso en gramos y como porcentaje del peso al sacrificio. Para su análisis estos porcentajes fueron sometidos a una transformación del arco seno de la raíz cuadrada de su proporción.
RESULTADOS Y DISCUSIÓN
Peso y condición corporal
Los efectos de la restricción alimenticia sobre los pesos y condiciones corporales se muestran en el Cuadro 1. Aunque los pesos y valores no mostraron diferencias significativas (P>0.05), los cambios globales fueron diferentes entre tratamientos (P<0.01).
Diversos autores(19,20,21) señalan que el peso corporal de animales restringidos no difiere del de aquéllos alimentados a libertad, indicando que los primeros pueden disminuir su tasa metabólica basal, y sus requerimientos para mantenimiento y así mantener el peso corporal. No obstante, las restricciones en todos los casos anteriores fueron de menor duración y en algunos casos se consideró como restricción alimenticia consumos de alimento similares a los indicados para mantenimiento de peso.
En contraste, cuando la restricción alimenticia es más severa y con una duración de meses, se ha encontrado que sí tienen efectos significativos sobre el peso corporal. En un trabajo previo(8), en el que se utilizaron cabras de características similares a las empleadas en el presente estudio y restringidas al 80 y 60% de sus requerimientos nutricionales, se observaron pérdidas de peso corporal significativas en comparación con el testigo no-restringido. Ovejas adultas que iniciaron con buena condición corporal (3.8) y que estuvieron sujetas a una restricción severa (80%) por 161 días, tuvieron pérdidas significativas de peso(19).
Aunque la CC es una medida subjetiva, junto con el peso dan una buena referencia del estado nutricional del animal. La pérdida de CC de los restringidos se debe tanto a la disminución de grasa como de masa muscular, principalmente de la primera.
Peso de órganos y tejidos viscerales
Los datos de pesos de órganos y tejidos se muestran en el Cuadro 2. Exceptuando al hígado, ninguno de los componentes medidos difirió entre tratamientos (P> 0.05), lo que concuerda con lo anteriormente informado(8,13). En contraste, otros autores(22) trabajando con borregas adultas con una restricción alimenticia (RA) del 80%, indican que el peso de la cabeza, del retículo-rumen y del colon, presentan una reducción significativa después de 161 días de tratamiento.
En el caso del hígado, tanto en este trabajo como en el de otros autores(13) el hígado como porcentaje de peso al sacrificio, se reduce significativamente (P<0.01), después de una RA severa (40%) de 18 semanas de duración y moderada (20%) y severa (40%) de 36 semanas de duración. En cuanto a otros órganos, los resultados coinciden también en forma parcial con los otros autores(7,8,23) que utilizando cerdos, ratas, ovinos y bovinos, observaron que una disminución en el plano nutricional consistentemente produce una reducción relativa (como porcentaje del peso al sacrificio) en el peso de hígado y otros órganos y concluyen que existe una buena relación entre los pesos del hígado y el tejido intestinal y la estimación de los requerimientos de energía de mantenimiento.
Pero, al utilizar una restricción alimenticia más severa (80%) por 161 días en ovejas adultas, se incrementan los órganos afectados, siendo además de hígado, riñón, compartimientos gástricos y colon; afectados también bazo y corazón; reduciéndose además la MS del líquido ruminal(22).
Análisis químicos
La composición química de las muestras, mostró que hígado, corazón, riñón, duodeno y el contenido retículo-ruminal, fueron afectados por la RA (Cuadros 3,4). En hígado se encontró mayor concentración de ADN (mg g-1 de tejido) a medida que se incrementaba la severidad de RA, lo que provocó que las relaciones ARN: ADN y proteína: ADN fueran mayores en el grupo testigo, comparados con los grupos con RA. En corazón se encontró menor cantidad de ADN y EE (mg g-1 de tejido) y MS total en los animales restringidos; debido a la dimensión de la pérdida de MS, es muy probable que tal reducción se deba no sólo a la pérdida de EE, sino también a la de proteína muscular; por otro lado al estar formado éste por células poli nucleadas, el tener más ADN no necesariamente nos hace pensar en que tiene mayor número de células. En riñón, aunque su peso no se mostró afectado por la RA, el análisis químico indica que las células renales de las restringidas tienen menor MS, lo que probablemente se deba a una utilización moderada de proteína. En duodeno, semejante a lo ocurrido en hígado, se encontró una menor relación de proteína:ADN en los restringidos, lo que hace pensar que de las tres porciones que integran el intestino delgado, éste resulta ser el más afectado por la RA. Finalmente, los animales restringidos tuvieron menos MS en el contenido ruminal en comparación con los testigos.
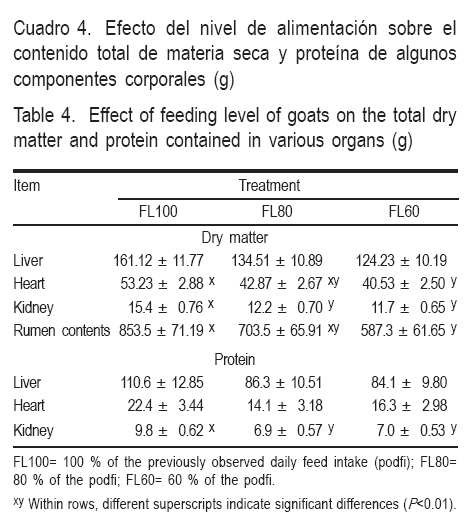
En general, los resultados de hígado coinciden con los de otros autores(24), que al estudiar el comportamiento de los ácidos nucleicos en novillos restringidos en el suministro de alimento, determinaron que la reducción en el tamaño del hígado se debió principalmente a una reducción en el tamaño celular (proteína:ADN), con pequeños cambios en el número de células (ADN. En cuanto a la concentración de ARN encontraron que se relacionaba directamente con el peso total del hígado, indicando que es un buen indicador de la capacidad de síntesis de proteína, ya que normalmente el 80% del total de ARN es ribosomal.
En intestino, la relación proteína:ADN no se redujo en los restringidos, pero sí la concentración total de ADN, lo cual sugiere que la reducción encontrada en el peso del intestino se debió a una reducción en el número celular.
Al estudiar la concentración de componentes tisulares en ratas, otros autores(6), señalan que al comparar animales ayunados 72 h con testigos, los primeros mostraron menor ARN (mg g-1 de tejido) y más alto ADN en hígado, riñón e intestinos, y reducción de ARN y masa proteica en intestino; lo que derivó en la menor relación de ARN: ADN y proteína: ADN en hígado, estómago y duodeno de las ratas en ayuno. Al hacer estas determinaciones en hígado de cabras subalimentadas por 36 semanas, se encontró disminución en la proteína, incremento en ADN, que resultó en decremento de las relaciones ARN:ADN y proteína:ADN. Se sugiere que la privación de nutrimentos reduce la concentración de ARN y proteínas de las vísceras debido a disminución del tamaño celular y la síntesis de proteína, como fue previamente sugerido(8).
Canales
Aunque el NA no tiene efecto sobre el peso o el rendimiento en canal, al hacer la separación de los componentes de la mitad derecha de ésta (Cuadro 5), se notan diferencias significativas en la proporción de hueso y tejido disectable. Las subalimentadas presentan mayor proporción de hueso y menor de tejido disectable que las testigo.
Las cabras subalimentadas mostraron mayores proporciones de hueso y niveles menores de tejidos blandos (especialmente músculo esquelético), estos últimos pudieron ser empleados activamente como fuentes de energía y nitrógeno metabólicos. Anteriormente nuestro grupo informó de menores rendimientos en canal después de 36 semanas de restricción, tuvieron menores rendimientos en canal que aquéllas subalimentadas por 18 semanas (13).
Otros autores(25) informan que en novillos, el peso de la canal caliente se redujo linealmente (P<0.04) por la RA de 10 y 20%, comparándolos con controles a libertad. Lo mismo sucede en corderos después de siete semanas de restricción de energía o proteína(26).
El tejido disectable de los restringidos presentó menor cantidad de EE (Cuadro 5), observación similar a otros trabajos(21) donde concluyeron que cuatro meses de RA provocaba menor proporción de grasa y mayor de hueso en vaquillas; con novillos(25) indican que la grasa contenida en la canal se reduce, mientras la proteína y el agua se incrementan como resultado de la reducción en el consumo de alimento durante 168 días. Finalmente nuestro grupo(6) mostró que el contenido de EE del músculo esquelético también se redujo por afecto de una RA de 36 semanas Todo lo anterior hace suponer que la grasa intramuscular es de los primeros depósitos en ser empleados como fuente energética en animales restringidos. En ese sentido Fattet et al(27) informan que la pobre utilización de la EM y el aparente efecto substitutivo de la proteína suplementaria, podría ser explicada bajo el supuesto que el metabolismo energético basal es dependiente de la ingestión de energía y, que una deficiencia proteica sería causa de una mayor pérdida de energía para el animal. Si uno o ambos supuestos fueran ciertos, entonces la totalidad del concepto de que el metabolismo en ayuno como base de los sistemas de alimentación se podría cuestionar, dado que el animal en ayuno, de la misma manera que moviliza reservas corporales de energía, se vuelve también deficiente en proteína.
Se ha informado que la suplementación proteica en animales en balance energético negativo resultaría en la acumulación de proteína y depleción de grasa corporal, respectivamente. Se puede esperar que animales maduros o jóvenes utilicen muy eficientemente la proteína suplementaria para contrarrestar pérdidas endógenas, y esos resultados podrían explicar los aquí mostrados.
Tejido adiposo
Los animales restringidos tuvieron menor cantidad de grasa disectable y de grasa peri-renal que los animales del tratamiento testigo (Cuadro 6). Las cabras restringidas presentaron mayor cantidad (mg g-1 de tejido) de ADN en riñón y tejido adiposo subcutáneo, siendo el primero estadísticamente significativo (P<0.01), lo que hace suponer que los adipocitos reducen su tamaño por efecto de la RA. Otros autores(28) en estudios sobre la variación de la actividad de la lipoproteína lipasa (LLP) en tejido adiposo blanco de ratas, durante un ayuno de tres días, informan que los adipocitos disminuyen de tamaño, lo mismo que la actividad de la LLP, con la realimentación a libertad. La actividad de la LLP retornó a valores normales en cuatro días y posteriormente en siete días el adipocito regresó a su tamaño inicial.
En cuanto al perfil de los ácidos grasos contenidos en el tejido adiposo, el único afectado por la RA más severa fue el C16:1 renal (P< 0.05) (Cuadro 7). Sin embargo, en el TAS el C16:1, C18:1 y AGI se encuentran en mayor proporción con un decremento en los ácidos grasos saturados totales, en los animales más restringidos (P<0.01). Los resultados obtenidos parecen ser muy diferentes a los de otras investigaciones, debido a que anteriormente, aún conociendo la gran importancia de los ácidos grasos de cadena corta(29), no se había incluido su cuantificación en forma completa o muestreado otro tipo de tejido adiposo(29,30,31).
Los cambios adaptativos pueden ser regulados, en parte, por la actividad de la D9 desaturasa (esteroil-CoA desaturasa), una enzima presente en los tejidos donde tiene lugar la síntesis de novo de ácidos grasos, siendo la enzima responsable de la introducción de dobles D9 en los ácidos grasos C16 y C18(31,32). Además se ha demostrado que la presencia de la enzima está altamente correlacionada con la cantidad de C18:1 en los depósitos grasos de la canal y del epicardio, pero no en los de omento y riñones(32,33).
CONCLUSIONES E IMPLICACIONES
Las cabras adultas tienen la capacidad de adaptarse a una restricción alimenticia prolongada, haciendo uso de combustibles metabólicos de diferentes tejidos; principalmente adiposo, como fuente de energía, y músculo esquelético y órganos viscerales metabólicamente activos como fuente de energía y aminoácidos, sin olvidar que con la reducción de éstos órganos se disminuyen los requerimientos de energía, lo que contribuye a la adaptación de estos animales a una reducción en el aporte de nutrimentos. Sin embargo, cabe aclarar que la CC inicial y por lo tanto la cantidad potencial de combustibles metabólicos disponibles, serán el principal factor para determinar el tiempo que estos animales soportarán dichas condiciones.
AGRADECIMIENTOS
Esto es parte de la tesis presentada por M. Gómez-Pastén, como requisito parcial para la obtención de grado de Doctor en Ciencias de la UNAM. Se realizó con apoyos de: Sistema de Investigación Miguel Hidalgo-Consejo Nacional de Ciencia y Tecnología (Proyecto ALIM-11/96); Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica-Dirección General de Asuntos del Personal Académico-UNAM (Exp.IN210396)
LITERATURA CITADA
1. Berg R, Butterfield R. New concepts of cattle growth. New York: John Wiley and Sons; 1976. [ Links ]
2. Birkelo C, Johnson D, Phetteplace H. Maintenance requirements of beef cattle as affected by season on different planes of nutrition. J Anim Sci 1991;69;1214-1222. [ Links ]
3. Aziz N, Murray D, Bell R. The effect of live weight gain and live weight loss on body composition of Merino wethers: dissected muscle, fat and bone. J Anim Sci 1992;70:1819-1828. [ Links ]
4. Freetly HC, Nienaber JA, Brown-Brandl T. Partitioning of energy in pregnant beef cows during nutritionally induced body weight fluctuation. J Anim Sci 2008;86:370-377. [ Links ]
5. Ledger H, Sayers A. The utilization of dietary energy by steers during periods of restricted food intake and subsequent realimentation. J Agric Sci 1977;88:11-26. [ Links ]
6. Ferrell CL, Koong L, Nieraber J. Effect of previous nutrition on body composition and maintenance energy costs of growing lambs. Br J Nutr 1986;56:595-605. [ Links ]
7. Burrin D, Britton R, Ferrell C. Visceral organ size and hepatocyte metabolic activity in fed and fasted rats. J Nutr 1988;118:1547-1552. [ Links ]
8. Gómez-Pastén M, Mora O, Pedraza-Chaverri J, Shimada A. The effect of a long term feed restriction on metabolism and tissue composition of goats. J Agric Sci 1999;132:227-232. [ Links ]
9. Moreno M, Martínez JA. El tejido adiposo: órgano de almacenamiento y órgano secretor. ANALES Sistema San Navarra 2002(25):29-39. [ Links ]
10. Cummins KA, Solaiman SG, Bergen WG. The effect of dietary copper supplementation on fatty acid profile and oxidative stability of adipose depots in Boer x Spanish goats. J Anim Sci 2008;86:390-396. [ Links ]
11. Instituto Nacional de Estadística, Geografía e Informática. Síntesis Geográfica, Nomenclator y Anexo Cartográfico del Estado de Querétaro. Secretaría de Programación y Presupuesto. México, 1986. [ Links ]
12. NRC. Nutrient Requirements of Goats. Natl. Acad. Press, Washington, DC. 1982. [ Links ]
13. Mora O, Shimada A, Ruiz FJ. The effect of the length and severity of feed restriction on weight, carcass measurements and body composition of goats. J Agric Sci 1996;127:549-553. [ Links ]
14. Santucci PM, Branca A, Napoleone M, Bouche R, Poisot F, Aumont G, Alexandre, G. Body condition scoring of goats in extensive conditions. In: Goat nutrition. Morand-Fehr P. editor. Wageningen, The Netherlands: Pudoc Publ; 1991. [ Links ]
15. AOAC. Official Methods of Analysis. 15th ed. Assoc. Offic. Anal. Chem., Arlington, VA. 1990. [ Links ]
16. Bradford M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem 1976;72:248-254. [ Links ]
17. Steel RG, Torrie JH. Bioestadística: Principios y procedimientos. 2nd ed., Bogotá: McGraw-Hill; 1988. [ Links ]
18. SAS User's Guide: Statistics. Statistical Analysis System Institute Inc., Cary, NC. 1994. [ Links ]
19. Burrin D, Ferrell C, Britton R, Bauer M. Level of nutrition and visceral organ size and metabolic activity in sheep. Br J Nutr 1990;64:439-448. [ Links ]
20. Drouillard JS, Klopfestein TJ, Britton RA, Bauer ML, Gramlich SM, Wester TJ, Ferrell CL. Growth, body composition, and visceral organ mass and metabolism in lambs during and after metabolizable protein or net energy restrictions. J Anim Sci 1991;69:3357-3375. [ Links ]
21. Yambayamba E, Price M. Growth performance and carcass composition in beef heifers undergoing catch-up (compensatory) growth. Can J Anim Sci 1991;71:1021-1029. [ Links ]
22. Atti N, Nozière P, Doreau M, Kayouli C, Bocquier F. Effects of underfeeding and refeeding on offals weight in the Barbary ewes. Small Ruminant Res 2000;38:37-43. [ Links ]
23. Kerr BJ. Considerations in the use of crystalline amino acids in swine diets [Ph.D. Thesis], Chicago, USA: Univ Illinois; 1988. [ Links ]
24. Sainz RD, Bentley BE. Visceral organ mass and cellularity in growth-restricted and refed beef steers. J Anim Sci 1997;75:1229-1236. [ Links ]
25. Murphy TA, Loerch SC. Effects of restricted feeding of growing steers on performance, carcass characteristics, and composition. J Anim Sci 1994;72:2497-2507. [ Links ]
26. Wester TJ, Britton RA, Klopfenstein TJ, Ham GA, Hickok DT, Krehbiel CR. Differential effects of plane of protein or energy nutrition on visceral organs and hormones in lambs. J Anim Sci 1995;73:1674-1688. [ Links ]
27. Fattet I, Deb F, Hovell D, Ørskov E, Kyle D, Pennie K, Smart R. Undernutrition in sheep. The effect of supplementation with protein on protein accretion. British J Nutri 1984;52:561-574. [ Links ]
28. Fried S, Hill J, Nickel M, DiGirolamo M. Prolonged effects of fasting-refeeding on rat adipose tissue lipoprotein lipase activity: influence of caloric restriction during refeeding. J Nutr 1983;113:1861-1869. [ Links ]
29. Bas P, Rouzeau A, Morand-Fehr P. Changes in the content of branched-chain fatty acids of the adipose tissue from different sites in growing goats. Proc. 4th Int. Conf Goats, Beijing, China. 1996. [ Links ]
30. Casey NH, Van Niekerk WA. Fatty acid composition of subcutaneous and kidney fat depots of Boer goats and the response to varying levels of maize meal. S Afr J Anim Sci 1985;15:60-62. [ Links ]
31. Banskalieva V, Saúl T, Goetsch AL. Fatty acid composition of goat muscles and fat depots: a review. Small Rum Res 2000;37:255-268. [ Links ]
32. Beswick NS, Kennelly JJ. Influence of bovine growth hormone and growth hormone releasing factor on messenger RNA abundance of lipoprotein lipase and steroyl-CoA desaturase in the bovine mammary gland and adipose tissue. J Anim Sci 2000;78:412-419. [ Links ]
33. Barber MC, Ward RJ, Richards SE, Salter AM, Buttery PJ, Vernon RG, Travers M. Ovine adipose tissue monounsaturated fat content is correlated to depot-specific expression of the stearoyl-CoA desaturase gene. J Anim Sci 2000;78:62-68. [ Links ]

