










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.3 no.5 Texcoco sep./oct. 2012
Artículos
Resistencia a roya amarilla (Puccinia striiformis f. sp. tritici) en variedades de trigo harinero (Triticum aestivum L.)*
Genetics of the resistance to yellow rust (Puccinia striiformis f. sp. tritici) in varieties of bread wheat (Triticum aestivum L.) cultivated in Bajío
Julio Huerta Espino1, Rocío Torres García1, María Florencia Rodríguez García1, Héctor Eduardo Villaseñor Mir1§, Santos Gerardo Leyva Mir2 y Ernesto Solís Moya3
1 Campo Experimental Valle de México. INIFAP. Carretera Los Reyes-Texcoco, km 13.5. Coatlinchán, Texcoco, Estado de México. C.P. 56260. Tel. 015959212657 (huerta.julio@inifap.gob.mx), (rocioiap@hotmail.com), (rodriguez.maria@inifap.gob.mx). §Autor para correspondencia: hevmir@yahoo.com.mx.
2 Universidad Autónoma Chapingo. Carretera México-Texcoco, km 38.5. C. P. 56230. Tel: 015959521500. Ext. 6179. (lsantos@correo.chapingo.mx).
3 Campo Experimental Bajío- INIFAP. C. P. 112.38000, Celaya Guanajuato, México. Tel. 014616115323.
* Recibido: octubre de 2011
Aceptado: agosto de 2012
Resumen
Las variedades de trigo harinero Cortazar S94, Bárcenas S2002, Urbina S2007 y Maya S2007 fueron liberadas para siembras en condiciones de riego en "El Bajío". Estas variedades mantienen resistencia a diferentes razas de roya amarilla que se encuentran en el país; sin embargo, se desconocen las bases genéticas de su resistencia. Para determinar la genética de la resistencia, estas variedades se cruzaron con el progenitor susceptible 'Avocet- YrA' y la determinación de similitud de genes se realizó mediante pruebas de alelismo. Las cuatro variedades y las familias F3 de cada cruza se evaluaron durante verano de 2009 en el Campo Experimental 'Valle de México' (INFAP-CEVAMEX) Chapingo, México, bajo una epifitia artificial creada con el aislamiento MEX96.11. Con los resultados de la evaluación de familias F3 se determinó que la resistencia de planta adulta a roya amarilla en Cortázar S94, Bárcenas S2002 y Maya S2007 está condicionada por dos ó tres genes de efectos aditivos, mientras que en Urbina S2007 la resistencia está condicionada por tres o cuatro genes de la misma naturaleza. La ausencia de familias completamente susceptibles en las cruzas entre las variedades resistentes indica la existencia de al menos un gen de resistencia en común.
Palabras claves: genes aditivos, resistencia, resistencia de planta adulta "El Bajío", variedades de trigo.
Abstract
The varieties of bread wheat, Cortázar s94, Bárcenas S2002, Urbina S2007 y Maya S2007 were liberated for planting under irrigated conditions in "El Bajío". These varieties maintain a resistance to different races of yellow rust that were found in the country; however, the genetic bases of resistance are unknown. To determine the genetics of the resistance, these varieties were crossed with the susceptible parent 'Avocet- YrA', and the determination of similitude ofthe genes was conducted through allele testing. The four varieties and the families F3 of each cross was evaluated during the summer of 2009 in the Experimental Field 'Valle de Mexico', Chapingo, Mexico State, under an artificial epiphyte created with the isolation MEX96.11. With the results of the evaluation of the families F3, it was determined that the resistance of the adult plant to yellow rust in Cortázar S94, Bárcenas S2002 y Maya S2007 is conditioned for two or three genes of additive effects, while in Urbina S2007, the resistance is conditioned for three or four genes of the same composition. The absence of complete acceptable families in the crosses between resistant varieties indicates the existence of at least one gene of resistance in common.
Key words: additive genes, resistance, adult plant resistance "El Bajío", wheat varieties.
Introducción
En México las principales regiones trigueras se ubican en el noroeste y el Bajío, donde se obtiene 53% y 28% de la producción total nacional, respectivamente. En ambas regiones el trigo se produce durante el invierno bajo condiciones de riego (SAGARPA, 2007), es el cultivo de invierno de mayor importancia en este ciclo y la economía regional depende en gran parte de la producción, industrialización y comercialización de dicho cereal (SIAP, 2011). A pesar de que México es un importante productor de este cereal, existe la necesidad de importar trigos panificables debido a que hay un déficit en su producción, ya que el tipo de trigo que más se cultiva es el durum o cristalino, que se destina a la fabricación de pastas (CANIMOLT, 2010).
Las variedades de trigo harinero que ocupan mayor superficie sembrada en el Bajío son Bárcenas S2002 con 29%, Cortázar S94 con 22% y Saturno S86 con 11%. (Singh et al, 2001) indican que la roya amarilla y de la hoja constantemente amenazan la producción de trigo de temporal y riego en el país, debido a la capacidad de vencer la resistencia especifica de variedades nuevas, a través de la evolución hacia nuevas razas fisiológicas con nuevos genes de virulencia.
Entre los principales problemas que enfrenta el cultivo de trigo en el Bajío, destaca la roya amarilla, enfermedad que se ha presentado severamente en siembras comerciales de trigo harinero en el ciclo otoño-invierno (Solís et al., 2007). Huerta y Singh (2000) indican que por mucho tiempo la raza 14E14 fue la más común en las zonas de el Bajío durante el invierno y se caracterizaba por su virulencia para los genes Yr2, 3, 6, 7, Sk (Yr 27) y A, entre otros. Durante el ciclo otoño-invierno 2004-2005 se observó una nueva raza identificada con el aislamiento número MEX03.37 que se caracterizó por infectar la espiga de diversas variedades que había sido resistentes incrementando los niveles de la enfermedad de las variedades recomendadas para riego como Salamanca S75, Pavón F76, Saturno S86 y Cortázar S94 entre otras, causando graves pérdidas en el rendimiento. Este aislamiento también se identificó en regiones como el Valle del Yaqui, Valle del Mayo, Costa de Hermosillo, Valle de Mexicali en Baja California Norte y en El Bajío, lo que ocasiono la necesidad de aplicación de fungicidas para reducir las pérdidas (Rodríguez et al, 2009). Ocasionando incremento en los costos de producción.
La estrategia que más ha apoyado el control de esta enfermedad es el uso de variedades resistentes, requiriéndose constantemente de fuentes de resistencia. Al recombinar una variedad susceptible por una resistente es posible determinar el tipo de acción génica que se presenta, ya que se puede observar si los genes actúan en forma dominante o en forma recesiva. Los trabajos de mejoramiento genético realizados en los últimos años tienen como objetivo generar genotipos con mayor potencial de rendimiento que las variedades comerciales de la región, resistentes a royas y de alta calidad industrial. Una alternativa para lograr mayor durabilidad de la resistencia es generar variedades que posean resistencia durable basada en genes que confieren resistencia de enroyamiento lento (slow rusting).
En el caso de la roya amarilla se deben combinar de 4 a 5 genes para reducir el progreso de la enfermedad a niveles tan bajos que sólo se puedan observar trazas al tiempo de madurez, bajo una presión alta de la enfermedad (Singh et al., 2001). Las variedades de trigo para siembras de riego Cortázar S94, Bárcenas S2002 (Solís et al., 2003), Urbina S2007 (Solís et al, 2008 a) y Maya S2007 (Solís et al, 2009) liberadas por el Programa de Mejoramiento Genético de Trigo (INIFAP-CEBAJ) mantienen diferentes niveles de resistencia a las razas de roya amarilla que se encuentran en el Bajío; sin embargo, se desconoce cómo opera la resistencia y los genes que están confiriéndola en estas variedades, para un uso más eficiente de las fuentes de resistencia que poseen los genotipos de trigo en México, es necesario conocer sus modos de herencia, similitud y cuantía de genes. Ante tal situación, se realizó la presente investigación con el objetivo de determinar la genética de la resistencia de planta adulta a roya amarilla de las cuatro variedades de trigo harinero antes indicadas.
Materiales y métodos
Material genético: se utilizaron las variedades de trigo Cortázar S94, Bárcenas S2002, Urbina S2007 y Maya S2007, que fueron liberadas para siembras de riego por el INIFAP-CEBAJ desde 1994 hasta 2007, estas variedades sobresalen por sus niveles de resistencia a roya amarilla. La cruza e historia de selección de dichas variedades se presentan en el Cuadro 1 y sus características se mencionan a continuación.
Cortázar S94: se obtuvo por hibridación y selección genealógica en el Instituto de Investigaciones Forestales Agrícolas y Pecuarias (INIFAP), de una cruza simple realizada en 1994. Se considera resistente a roya amarilla (Puccinia striiformis) y a roya de la hoja (Puccinia triticina Ericks.) (Solis et al, 2000).
Urbina S2007: se logró por hibridación y selección a través del método de cruzas masivas F3 En el progenitor femenino intervinieron cuatro genotipos recombinados en tres cruzamientos, y el progenitor masculino fue la cruza Casilda/Centella. Se considera resistente a roya amarilla (Puccinia striiformis) y a roya de la hoja (Puccinia triticina Ericks.) (Solis et al, 2008 b).
Maya S2007: se produjo por hibridación y selección genealógica en el INIFAP, de una cruza simple realizada en 1997. Se considera resistente a roya amarilla (Puccinia striiformis) y a roya de la hoja (Puccinia triticina Ericks.) (Solis et al, 2009).
Bárcenas S2002: se recabó por hibridación y selección genealógica en el INIFAP. Se considera resistente a roya amarilla (Puccinia striiformis) y a roya de la hoja (Puccinia triticina Ericks.) (Solis et al, 2003).
Se utilizó el genotipo Avocet-YrA que es susceptible en plántula y planta adulta a todas las razas fisiológicas de roya amarilla que existen en México, y que alcanza una severidad de 100%. Raza del patógeno la fórmula de avirulencia/virulencia del aislamiento MEX96.11, es: Yrl, 4, 5, 8,10,15,17, 24, 26, Sp/Yr2, 3, 6, 7,9,27, basado en las líneas diferenciales derivadas de Avocet (Singh et al, 2000).
Obtención de progenies las cruzas se hicieron durante el ciclo otoño-invierno 2007-2008 en los invernaderos del Campo Experimental Valle de México (CEVAMEX), perteneciente al Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias (INIFAP), ubicado en Chapingo, México, 19° 53' latitud norte 19' y 99° 53' longitud oeste, a una altitud de 2 250 msnm, con promedio anual de precipitación de 640 mm y 15 °C de temperatura media anual (García, 1981). Se hicieron 10 cruzas como un dialelo parcial sin las cruzas reciprocas, de las cuales cuatro fueron, susceptible x resistente y las seis restantes fueron cruzas resistente x resistente Cuadro 2.
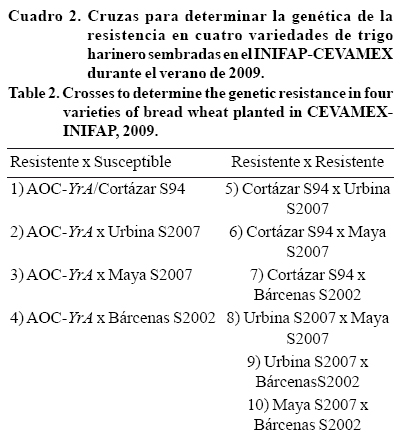
Obtención de F1, F2 y Familias F3 de las cruzas resistente por susceptible la generación F1 de cada una de las cruzas se sembró en campo en forma mateada durante el ciclo primavera-verano 2008 en el CEVAMEX. De cada cruza se cosecharon de cuatro a cinco plantas al azar, tres de las cuales dieron origen a la generación F2 y las semillas restantes se guardaron como reserva. Las progenies de cada planta F1 fueron sembradas individualmente en forma graneada en el ciclo otoño-invierno 2008-2009 en el CEVAMEX. Para la obtención de las familias F3, las poblaciones F2 de las tres plantas de cada cruza se inspeccionaron visualmente, y como no hubo discrepancia entre sus progenies, se seleccionaron 40 plantas individuales de cada una, para la obtención de 120 familias F3.
Obtención de F1, F2 y familias F3 de las cruzas resistente por resistente la generación F1 de cada una de las cruzas se sembró en campo en forma mateada durante el ciclo primavera-verano 2008 en el CEVAMEX. De cada cruza se cosecharon de dos a cuatro plantas al azar, dos de las cuales dieron origen a la generación F2 y las semillas restantes se guardaron como reserva. Las progenies de cada planta F1 fueron sembradas individualmente en forma graneada en el ciclo otoño-invierno 2008-2009 en el CEVAMEX. Para la obtención de semilla de las familias F3, las poblaciones F2 de las dos plantas de cada cruza se inspeccionaron visualmente, y como no hubo discrepancia entre sus progenies, se cosecharon 50 plantas individuales de cada una, para la obtención de 100 familias F3 por cruza.
Evaluación de familias F3 de las cruzas susceptible por resistente se sembraron los progenitores y los diferentes números de familias los cuales variaron de 118 a 120 dependiendo de la cruza así, el número fue de 119 familias en la cruza AOC-YrA x Cortázar S94, 118 en la cruza AOC-YrA x Maya S2007 y 120 en las cruzas AOC-YrA x Urbina S2007 y AOC-YrA x Bárcenas S2002, las cuales se sembraron en surcos dobles de 1 m x 0.80 m de separación, en el campo experimental del CEVAMEX-INIFAP. Alrededor del experimento y entre las calles se sembró semilla del genotipo susceptible Morocco + Lr19, el cual actuó como fuente de inoculó de la roya amarilla y dispersante del mismo.
Evaluación de familias F3 de las cruzas resistente por resistente se sembraron los progenitores y 98 familias F3 de cada cruza, esto con el fin de realizar la prueba de alelismo como se indica en el cuadro 2.
inoculación de familias F3 se estableció una epifitia artificial 22 días después de la siembra, lo que se logró mediante tres inoculaciones de esporas frescas de roya amarilla que fueron suspendidas en aceite mineral (Sotrol 170®), (la primera el 17 de julio, con el fin de asegurar que el hongo se estableciera a tiempo, se hicieron dos inoculaciones más el 22 y 27 de julio de 2009) la inoculación se realizó utilizando atomizadores manuales, asperjando directamente la superficie de las hojas de las plantas, en los bordos sembrados en los surcos laterales y calles del experimento ocupadas con el material susceptible Morocco + Lr 19.
Análisis genético las frecuencias observadas y esperadas se compararon mediante la prueba de Ji-cuadrada (X)2 el valor de tablas y la significancia fue determinada de acuerdo a la Ji-cuadrada (X2) que obtuvieron las proporciones de las familias de cada cruza. Para el valor de tablas se usaron n-1 grados de libertad, donde n es el número de grupos de clasificación de familias F3 (Infante y Zárate de Lara, 1990).
Resultados y discusión
Desarrollo de la enfermedad y toma de notas las condiciones de humedad y temperatura fueron óptimos para el desarrollo de la enfermedad, y antes de la floración se observaron los primeros signos de roya amarilla de tal forma que la primera evaluación del comportamiento de las familias F3 y de los progenitores involucrados en cada cruza se realizó en agosto 29, cuando el progenitor susceptible alcanzó niveles de infección entre 70 y 80% de acuerdo con la escala modificada de Cobb (Peterson et al, 1949).
En esta primera lectura se clasificaron las familias de cada cruza en cuatro grupos como sigue: grupo 1= familias homocigóticas tan resistentes o más (transgresivas) como el progenitor resistente; grupo 2= familias homocigóticas tan susceptibles o más que el progenitor susceptible; en este caso Avocet-YrA; grupo 3= familias heterocigóticas (segregando) con presencia de plantas tan resistentes como el progenitor resistente y plantas con niveles de infección intermedia o menos resistentes que el progenitor resistente y ausencia de plantas completamente susceptibles; y grupo 4= familias heterocigóticas con plantas tan resistentes o más que el progenitor resistente, plantas intermedias y plantas tan susceptibles o más que el progenitor susceptible.
Una segunda clasificación de las familias en campo permitió corroborar que las familias clasificadas en el grupo 1 permanecieron como tal o se reclasificaron en el grupo 3 si los niveles de infección fueron 5% mayor que el progenitor resistente.
Frecuencias esperadas de los diferentes grupos en las cruzas susceptible x resistente las frecuencias esperadas de las familias F3 de los cuatro grupos de clasificación se ilustran en el Cuadro 3, bajo el supuesto de que la resistencia está condicionada por 2, 3, 4, y 5 genes menores de efectos aditivos; el número de genes implicados en la resistencia se determinó con base en las frecuencias observadas de las familias clasificadas en el grupo 2 y comparadas con las frecuencias esperadas del Cuadro 3. Por ejemplo, si la resistencia estuviera controlada por dos genes las frecuencias esperadas de familias homocigóticas susceptibles de 6.25% (1/16); si fuera condicionada por tres genes, la proporción sería 1.6% (1/64), y si estuviera controlada por cuatro genes, la proporción sería equivalente a 0.4% (1/256).

La distribución, frecuencias relativas de las familias F3, el número de genes que condicionan la resistencia y la prueba de Ji- cuadrada (X2) de las cuatro cruzas susceptible x resistente, se muestran en el Cuadro 4.
El criterio para determinar el número de genes se basó en el número de familias homocigóticas susceptibles, las cuales son más fáciles de identificar en campo, bajo el supuesto de que la virulencia del patógeno es recesiva y que la resistencia en planta es dominante (Roelfs y Groth, 1988). La frecuencia de familias F3 homocigótica similares al progenitor resistente o susceptibles fueron en frecuencia muy baja lo que indica que la resistencia fue compleja, condicionada por más de un gen y no se basa en genes de efectos mayores.
Con base en las frecuencias esperadas y observadas (Cuadro 4), y mediante la prueba de X2, se determinó que de dos a tres es el número mínimo de genes de efecto aditivo que controlan la resistencia en las variedades Cortázar S94, Bárcenas S2007 y Maya S2007. Como en la variedad Urbina S2007 se incrementó la frecuencia de familias de tipo intermedio, grupo 3 con respecto a las familias susceptibles, grupo 4, probablemente en este caso el número de genes sea de tres a cuatro. Sin embargo; la frecuencia de familias susceptibles fue mucho mayor que las familias resistentes, como se puede observar en el Cuadro 4. Lo anterior sugiere la posibilidad de mezclas mecánicas y presencia de plantas voluntarias cuando se sembró la F2 para en la obtención de familias F3. Esto trae como consecuencia que el número de genes que están confiriendo la resistencia a la roya amarilla en las variedades liberadas para su siembra en el Bajío sea subestimado. Sin embargo, si el número de genes se determina por la frecuencia de familias tan resistentes como el progenitor resistente; se podría inferir que la resistencia a la roya amarilla está determinada por 2-3 genes en Cortázar S94, Maya S2007 y Bárcenas S2007, y de 3-4 en Urbina S2007.
En la variedad Bárcenas S2002, se observó la presencia de necrosis en la punta de la hoja (Ltn= Leaf tip necrosis), carácter morfológico manifestado cuando está presente el gen de resistencia a roya de la hoja Lr34, y también ligado al gen Lr46. El gen Lr34 es el mismo que condiciona resistencia a roya amarilla y se denomina como Yr18 (Kratinger et al., 2009); este gen ayuda a reducir la severidad de la enfermedad, pero individualmente es insuficiente por lo que los niveles de desarrollo de la enfermedad se llegan a incrementar hasta 70%.
El gen Lr46 también está asociado a la quemadura de la punta de la hoja (Ltn). Estudios realizados en CIMMYT han mostrado que el gen Lr46 está fuertemente ligado al gen Yr29 que confiere resistencia de enroyamiento lento a la roya amarilla, el cual tiene efecto similar al del Yr18. Otro gen de efectos menores es Yr30 y se considera de naturaleza durable con efecto aditivo al interactuar con otros genes de efectos aditivos.
Villaseñor et al. (2009) encontró resultados similares en variedades de trigo para temporal donde se muestra que los altos niveles de resistencia, están siendo conferidos por la presencia de tres y cuatro genes de efecto aditivo. Singh y Rajaram (1994) determinaron que la resistencia de Puccinia striiformis está controlada por uno a tres genes aditivos. En este estudio se observó que un mayor número de genes confirió mayor resistencia en los progenitores utilizados, los cuales tuvieron bajos niveles de infección.
Los resultados de las familias F3 indican que la variedad Urbina S2007 posee al menos un número mayor de genes que las otras tres variedades, no obstante es necesario analizar si las cuatro variedades poseen genes diferentes.
En el Cuadro 5 se presenta la distribución de familias F3, de las cruzas entre los progenitores resistentes, sembrados en el CEVAMEX durante el verano, 2009.
En trigo y en otros cereales, la similitud de genes de resistencia, que se puede observar al evaluar las progenies resultantes de las cruzas entre progenitores resistentes, se hace evidente cuando no se observa segregación en dichas progenies de familias homocigóticas susceptibles (Mariscal et al, 2010).
En ninguna de las familias de las cruzas resistente por resistente se observaron familias susceptibles (grupo 2), tampoco se observaron familias segregantes del grupo 4, lo que sugiere que los genes que condicionan la resistencia en estas variedades son los mismos; la otra posibilidad es que por lo menos un gen de efectos aditivos sea común en las cuatro variedades.
Adhikari et al. (1999) al hacer estudios genéticos entre genotipos de avena con el progenitor resistente "omega" con otras 15 líneas resistentes, observaron que todas las familias de estas cruzas fueron resistentes homocigóticas, indicando que la falta de segregantes susceptibles se debió a que los progenitores tenían los mismos genes o éstos eran diferentes, probablemente eran genes ligados o alélicos.
McKenzie et al. (1965), concluyeron que la cruza entre los progenitores 'Rosen's Mutant*'C.I.6829' poseen el mismo gen 'H', que confiere resistencia a la raza 6AF de roya del tallo esto a observar que las 33 familias F3 fueron resistentes.
La presencia de la quemadura de la punta de la hoja podría indicar que este gen posiblemente sea Yr18; que alguna vez se postuló en Salamanca S75 y que está como progenitor en todas estas variedades resistentes (Cuadro 1); sin embargo, el análisis molecular indica que la variedad Salamanca S75 no posee el gene Yr18, el cual confiere resistencia de enroyamiento lento a la roya amarilla y que está ligado al gene Lr34 localizado en el genoma 7D, Kolmer et al. (2008).
La otra posibilidad es que sea el gen Lr46, el cual está ligado al gen Yr29 William et al. (2003); y también ligado a la quemadura de la punta de la hoja Rosewarne et al. (2006). Pruebas moleculares indican la presencia de Yr29 en las cuatro variedades (Huerta-Espino datos sin publicar), siendo Salamanca S75 el progenitor común en las cuatro variedades y en donde se ha determinado la presencia del gene Yr29 Kolmer et al. (2008).
En variedades como Pavón F76 y Huites F95 que no poseen el gen Yrl 8, también es posible reducir los niveles de infección mediante otros genes que igualmente inducen este tipo de resistencia, como Yr29 y Yr3 Singh et al. (2001 y 2003).
Por otro lado, se ha determinado que el gen Yr18 está presente en Urbina S2007 y Cortázar S94, pero no en Bárcenas S2002 y Maya S2007, mientras que Yr30 está presente en Cortázar S94, Bárcenas S2002 y Maya S2007 pero no en Urbina S2007.
Singh y Dubin (1997) y Singh et al. (2000) encontraron que las combinaciones de Yr18 más o tres genes de la misma naturaleza inducen niveles aceptables de resistencia, y que a mayor número de genes de efecto aditivo la resistencia es más estable en cualquier ambiente que presente roya amarilla.
Un gen adicional localizado en el cromosoma 7B, también está presente en Urbina S2007, pero no en las otras variedades. Lo anterior explica la ausencia de familias susceptibles y confirma el efecto aditivo entre los genes antes mencionados y en particular la del gen Yr29. El mayor nivel de resistencia y número de genes en Urbina S2007 indica la presencia de un gen adicional y diferente a los antes mencionados.
Conclusiones
Con base en los resultados de las familias F3 de las cruzas resistente x susceptibles, se concluye que la resistencia de planta adulta a roya amarilla en la variedad Urbina S2007 está determinada por al menos tres genes de efectos aditivos, esta variedad posee mayor número de genes o al menos un gen diferente que le confiere mayor nivel de resistencia. En las variedades Cortázar S94, Bárcenas S2007 y Maya S2007 se identificaron entre dos y tres genes confiriendo la resistencia.
En las cruzas de familias resistentes x resistentes no se observaron familias susceptibles lo que indica que estas variedades tienen al menos un gen en común.
Literatura citada
Adhikari, K. N.; McIntosh, R. A. and Oates, J. D. 1999. Inheritance of the stem rust resistance phenotype. Pg-a in oats. Euphytica. 105:143-154. [ Links ]
García, E. 1981. Modificaciones del sistema de clasificación climática de Köppen (adaptada a condiciones de la república mexicana). 3ª ed. México, D. F. 86 p. [ Links ]
Infante, G. S. y Zárate de Lara, G. P. 1990. Métodos estadísticos: un enfoque interdisciplinario. 2a ed. Trillas. México, D. F. 643 p. [ Links ]
Kolmer, J. A.; Singh, R. P.; Garvin, D.F.; Viccars, L.; William, H. M.; Huerta, E. J.; Ogbonnaya, F. C.; Raman, H.; Orford, S.; Bariana, H. S. and Lagudah, E. S. 2008. Analysis of the Lr34/Yr18 rust resistance region in wheat germplasm. Crop Sci. 48:1841-1852. [ Links ]
Krattinger, S. G.; Lagudah, E. S.; Spielmeyer, W.; Singh, R. P.; Huerta, E. J. McFadden, H.; Bossolini, E.; Selter, L. L. and Keller, B. 2009. A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science 323:1360-1363. [ Links ]
Mariscal, A. L.A.; Huerta E. J.; Villaseñor, M. H. E.; Leyva, M. S. G.; Sandoval, I. S. y Benítez, R. I. 2010. Prueba de similitud de genes con resistencia a roya del tallo en genotipos de avena. Rev. Mex. Cien. Agríc. 1(4):541-554. [ Links ]
McKenzie, R. I. H.; Fleischmann, G. and Green, G. J. 1965. A close association of stem and crown rust resistance in 'Ukraine' and 'Rosen's Mutant' oats. Crop Sci. 5:551-552. [ Links ]
Reporte Estadístico de la Cadena Agroindustrial 2010. CANIMOLT. Medio de comunicación anual. Ediciones CANIMOLT. 25 p. [ Links ]
Roelfs, A. P and Groth, J. V. 1988. Puccinia graminis f. sp. tritici black stem rust of Triticum spp. Genet. Plant Pathol. 6:345-361. [ Links ]
Rodríguez, G. M. F.; Huerta, E. J. Villaseñor, M. H. E. y Solís, M. E. 2009. Virulencia de la roya amarilla del trigo en las principales zonas productoras de riego en México. Agric. Téc. Méx. 35(2):179-187. [ Links ]
Rosewarne, G. M.; Singh, R. P.; Huerta, E. J.; William, H. M.; Bouchet, S.; Cloutier, H.; McFadden and Lagudah, E. S. 2006. Leaf tip necrosis, molecular Markers and 1 -proteasome subunits associated with the slow rusting resistance genes Lr46/Yr29. Theor Appl. Genet. 112:500-508. [ Links ]
Secretaría de Agricultura, Ganadería Desarrollo Rural, Pesca y Alimentación (SAGARPA)- Comisión Estatal del Agua (CEA). Anuario estadístico 2007. Base de datos, México, D. F. [ Links ]
Servicio de Información Agroalimentaria y Pesquera (SIAP). 2011. Secretaría de Agricultura Ganadería, Desarrollo Rural, Pesca y Alimentación. (SAGARPA). México, D. F. http://www.siap.sagarpa.gob.mx. [ Links ]
Singh, R. P. and Rajaram, S. 1994. Genetics of adult plant resistance to stripe rust in ten spring bread wheats. Euphytica 72:1-7. [ Links ]
Singh, R. P. and Dubin, J. H. 1997. Sustainable control of wheat diseases in México. In: Mem. First International Wheat Symposium. Cd. Obregón. Sonora, México. 39-103 pp. [ Links ]
Singh, R. P.; Huerta E. J. and Rajaram. S. 2000. Achieving near-immunity to leaf and stripe rust in wheat by combining slow rusting resistance genes. Acta Phytopathol. Entomol. Hungarica. 35:133-139. [ Links ]
Singh, R. P.; Huerta, E. J. and William, H. M. 2001. Resistencia durable a roya de la hoja y roya amarilla del trigo. In: estrategias y metodologías utilizadas en el mejoramiento de trigo un enfoque multidiciplinario. Man, M. K.; Díaz, A. M. and Castro, M. (Eds.). La estanzuela Uruguay 8-11 de Octubre. 109-117 pp. [ Links ]
Singh, R. P.; Huerta, E. J y William, H. M. 2003. Resistencia durable a roya de la hoja y roya amarilla del trigo: Genética y mejoramiento del trigo en el CIMMYT In: estrategias y metodologias utilizadas en el mejoramiento de trigo. [ Links ]
Solís, M. E.; Salazar, Z. A.; Huerta, E. J.; Ramírez, J. R. A.; Villaseñor, M. H. E. y Espitia, R. E. 2003. Bárcenas S2002: nueva variedad de trigo harinero para el Bajio. Rev. Fitotec. Mex. [ Links ]
Solís, M. E.; Huerta, E. J.; Villaseñor, M. H. E. y Aguado, S. C. 2007. Roya lineal, fenología rendimiento y sus componentes en trigo harinero (Triticum aestivum L). [ Links ]
Solís, M. E.; Huerta, E. J.; Pérez, H. P.; Ramírez, R. A.; Villaseñor, M. H. E.; Espitia, R. E. y Borodanenko, A. 2008. Urbina S2007: nueva variedad de trigo harinero para la región del Bajio. Agric. Téc. Méx. 34(1):113-118. [ Links ]
Solís, M. E. Huerta, E. J.; Villaseñor, M. H. E.; Ramírez, R. A. y Pérez, H. P. 2009. Maya S2007: Nueva variedad de trigo harinero para la región el Bajío, México. Agric. Téc. Méx. 34(3):365-370. [ Links ]
Villaseñor, E. O. M.; Huerta, E. J.; Leyva, M. S. G.; Villaseñor, M. H. E.; Singh, R. P.; Sandoval, I. J. S. y Espitia, R. E. 2009. Genética de la resistencia a roya amarilla en plantas adultas de trigo harinero. Rev. Fitotec. Mex. 32(3): 217-233. [ Links ]
William, M. R.; Singh, R. P.; Huerta, E. J.; Ortiz, I. J. and Hoisington. 2003. Molecular marker mapping of leaf rust resistance gene Lr46 ant its association with stripe rust resistance gene Yr29 in wheat. Phytopathology. 93:153-159. [ Links ]

