











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Al momento de evaluar la eficiencia de una dieta en el animal, existe el concepto de energía metabolizable, también se puede hablar de proteína, grasa, fibra, carbohidratos, y cualquier otro nutriente que es absorbido por el organismo. Estos nutrientes metabolizables pueden expresarse por día, en % o por gramo de alimento consumido. Al respecto, Pirgozliev et al. (2024) mencionan este concepto de nutrientes metabolizables para pollos de engorda. Mientras que, Anwar et al. (2023) y Abella et al. (2024) mencionan valores de calor de combustión, materia seca, fibra cruda, proteína, lípidos y cenizas metabolizables. También se reportan datos de materia seca, calcio y fósforo metabolizables (Adekoya y Adeola 2023). Estos nutrientes son fáciles de analizar químicamente y pueden evaluarse con los métodos biológicos rápidos propuestos para la medición de la energía metabolizable aparente (EMA) o verdadera (EMV).
La energía metabolizable en aves se ha venido estudiando desde hace varios años (Li et al. 2024, Ye et al. 2024, Noblet et al. 2024, Aguirre et al. 2024). Estos autores citan los trabajos pioneros de Hill, Sibbald y Farrell y sus colaboradores, mismos que datan de más de 40 años. Paralelamente se ha estudiado en relación con el metabolismo del nitrógeno, fósforo y calcio (Dias et al. 2023, Adekoya y Adeola 2023). Se han propuesto la energía metabolizable aparente (EMA), verdadera (EMV) y a balance cero de nitrógeno (EMAn o EMVn), las cuales se han evaluado para aves jóvenes como pollitos (Khalil et al. 2023, Abella et al. 2024), o aves adultas como gallos maduros (Li et al. 2023). La energía metabolizable se ha medido haciendo recolección total de heces o usando marcadores inertes como óxido de cromo, dióxido de titanio, cenizas insolubles en ácido u óxido férrico (Macie et al. 2020, Adekoya y Adeola 2023, Eshtejarani et al. 2024, Reid et al. 2024), y se ha medido haciendo alimentación forzada, restringida o ad libitum (Li et al. 2024, Wu et al. 2020).
Hay controversia respecto al uso de EMA o EMAn ésta última subestima el valor de energía del alimento por lo que se sugiere formular con EMA en lugar de EMAn (Abdollahi et al. 2021). Las EMVs requieren estimar por regresión o colectar las excretas de gallos en ayuno, cuyo calor de combustión es la energía metabólica y endógena (Aguirre et al. 2024, Wu et al. 2020). También se pueden usar muestras del contenido ileal para la determinación de la energía digestible (Xavier et al. 2024) o medir la energía retenida en el cuerpo del animal (Barzegar et al. 2020). Estos métodos requieren el sacrificio de animales y el segundo mide más bien la energía neta de crecimiento o de mantenimiento. Sánchez-Peña et al. (2017) mencionan que el valor de EMA de una dieta depende de muchos factores como edad, tipo de ingrediente, genotipo, frecuencia de alimentación, sexo y consumo de alimento. Jiang et al. (2024), encontraron que las aves adultas poseen más habilidad para metabolizar la energía de los ingredientes que las jóvenes.
En el presente trabajo no se utilizó alimentación forzada, los valores obtenidos son aparentes (no verdaderos) y no se utilizaron gallos en ayuno. Por lo anterior, el objetivo fue evaluar la energía y los nutrientes metabolizados de una dieta en cantidad (g día-1 o cal día-1) y en porcentaje usando dos genotipos de gallos (Criollo y Sasso) y dos niveles de consumo (100 y 50 g día-1), asimismo conocer el efecto del tiempo de medición (1 a 7 días) en los nutrientes metabolizados.
Materiales y métodos
Aves
Se utilizaron seis gallos Criollos mexicanos y seis gallos de raza Sasso de 29 semanas de edad alojados en jaulas individuales (1×1×1 m, a un m del piso), con comedero y bebedero, y una charola de plástico debajo de cada jaula para la recolección de excretas. El peso promedio de los Criollos fue 2.29 ± 0.172 y el de los Sasso 2.75 ± 0.265 kg, ambos numéricamente parecidos entre sí, aunque estadísticamente diferentes (p < 0.05). Las jaulas están dentro de una caseta que posee cortinas móviles y ambiente natural. Todos los animales se manejaron siguiendo las recomendaciones del Comité de Bienestar Animal del Colegio de Posgraduados (COLPOS 2016).
Recolección de excretas y consumo
Se utilizó el método de recolección total de excretas (Wu et al. 2020, Eshtejarani et al. 2024), cuyas impurezas se retiraron usando pinzas de disección cada 24 h por 7 días. Las excretas recolectadas se colocaron en bolsas de plástico a 0 °C. Previo a su secado (50 °C) y para evitar pérdidas de nitrógeno, se agregó 1 cm3 de ácido sulfúrico 1 N por cada 10 g de excretas para obtener un pH alrededor de 5, y posteriormente se molieron por malla de 1 mm resultando un polvo fino. Después de entrenar y adaptar los gallos a las dietas durante 2 días (Li et al. 2023, Wu et al. 2020), los siguientes 4 se ofrecieron 100 g de alimento a cada gallo y 50 g otros 3 días más. El agua se proporcionó siempre ad libitum.
Nutrientes metabolizables o variables de respuesta
En las excretas y en el alimento, se determinaron proteína cruda, extracto etéreo, fibra cruda y calor de combustión con base a las metodologías del Análisis Proximal y de Van Soest (AOAC 1990, Van-Soest et al. 1991). Estas determinaciones sirvieron para calcular los nutrientes metabolizables inicialmente en g día-1 o en kcal día-1 y después en %: (nutriente en el alimento en %×consumo en g o kcal - nutriente en las excretas en %×excretas en g o kcal)/100%. Los valores anteriores relacionados a la cantidad de nutrientes en el alimento consumido (100%) sirvieron para calcular los nutrientes metabolizables en % y divididos entre el número de días, quedaron en g o kcal día-1. Además, dividiendo la energía metabolizada entre el consumo en g, se calculó la EMA de la dieta en kcal g-1. Todas éstas se consideran las variables de respuesta a los distintos factores en estudio (día de evaluación, genotipo y consumo de alimento).
Sitios experimentales
Las aves estuvieron en la Granja Experimental del Colegio de Postgraduados y las determinaciones químicas se realizaron en el Laboratorio de Nutrición Animal del Departamento de Zootecnia de la Universidad Autónoma Chapingo. Ambas instituciones ubicadas en el Municipio de Texcoco, Estado de México, México.
Dieta
En la Tabla 1 se muestra la dieta utilizada, 50% de harina de trébol blanco y 50% de una dieta basal. La dieta se proporcionó en forma de pellets. Los análisis calculados se realizaron con base a Rattray y Joyce (1974), Berardo (1997), Cammell et al. (1986), Stypiñsk (1993) y NRC (1994).
Tabla 1 Dieta otorgada de la cual se ofrecieron 100 o 50 g por gallo por día.
| Ingredientes | Contenido, % BTO | Nutrientes (calculados) | Contenido, % BS |
|---|---|---|---|
| Trébol | 50.00 | Energía metabolizable | 2.75 |
| Maíz Amarillo, grano | 29.09 | Proteína cruda | 21.80 |
| Pasta de soya | 13.38 | Fibra cruda | 10.02 |
| Olote de maíz | 5.44 | Extracto etéreo | 3.30 |
| Ortofosfato de calcio | 0.76 | Cenizas | 7.94 |
| Carbonato de calcio | 0.58 | Humedad | 12.19 |
| Sal común | 0.18 | Lisina | 0.96 |
| Bicarbonato de sodio | 0.05 | Metionina | 0.35 |
| L-Lisina-HCl | 0.15 | Metionina + Cistina | 0.56 |
| DL-Metionina | 0.12 | ||
| L-Treonina | 0.08 | ||
| Vitaminas y Minerales1 | 0.20 | ||
| Total | 100.00 |
1Por kilogramo de dieta basal (sin trébol): vitamina A, 12.000 UI; vitamina D3, 1.000 UI; vitamina E, 60 UI; vitamina K, 5.0 mg; vitamina B2, 8.0 mg; vitamina B12, 0.030 mg; ácido pantoténico, 15 mg; niacina, 50 mg; ácido fólico, 1.5 mg; colina, 300 mg; biotina, 0.150 mg; tiamina, 3.0 mg. fierro, 50.0 mg; zinc, 110 mg; manganeso, 100 mg; cobre, 12.0 mg; selenio, 0.3 mg; iodo, 1.0 mg.
Tratamientos y diseño experimental
Se establecieron dos diseños completamente aleatorizados, uno con arreglo factorial 2×2, con 6 repeticiones, consumo (100 y 50 g) y genotipo (Criollo y Sasso). Otro diseño fue de un solo factor (tiempo de medición de 1 a 7 días). La unidad experimental fue un gallo, con 6 repeticiones de cada genotipo y 7 días en observación. Por tanto, el número total de observaciones fue 6 gallos×2 genotipos×7 días = 84.
Análisis estadísticos
Las medias del factor genotipo se compararon con la prueba F de Fisher (p < 0.05) y así se hizo con las medias del factor consumo. Las medias de los 7 tratamientos del factor tiempo fueron separadas por medio de la prueba de Tukey (p < 0.05). Todos los análisis estadísticos se realizaron empleando el paquete estadístico SPSS® (2011)
Resultados
Efectos del factor tiempo
En la Tabla 2 se presentan los efectos del factor tiempo en todas las variables metabolizables de respuesta evaluadas. Los días 1 a 4 casi todas las variables fueron similares (p > 0.05) pero mayores que sus valores en los días 5 a 7. Es decir, al avanzar el día de evaluación, la mayoría de las variables disminuyeron (p < 0.05). Por ejemplo, de los días 1 a 7, la PC metabolizable disminuyó (p < 0.05) 11.9 g día-1 y la EMA disminuyó (p < 0.05) 197.2 kcal día-1 y varió (p < 0.05) de 12.76 a 13.23 en los días 1 a 4 y de 1.33 a 2.93 g día-1 en los días 5 a 7 (Tabla 2).
Tabla 2 Efectos del tiempo de medición en la energía y los demás nutrientes metabolizables.
| Día | |||||||
|---|---|---|---|---|---|---|---|
| Variable | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
| Proteína cruda (PC) metabolizable, g día-1 | 13.23a* | 13.80a | 12.76a | 12.96a | 1.66b | 2.93b | 1.33b |
| PC metabolizable, % | 56.77a | 59.22a | 56.43a | 57.33a | 14.83b | 25.89ab | 11.73b |
| Extracto etéreo (EE) metabolizable, g día-1 | 1.03b | 1.04b | 1.74a | 1.68a | 0.69c | 0.79c | 0.77c |
| EE metabolizable, % | 80.67 | 80.90 | 86.87 | 83.70 | 69.22 | 78.72 | 76.33 |
| Fibra cruda (FC) metabolizable, g día-1 | 6.62ab | 6.98a | 5.82bc | 5.64c | 0.33e | 1.81d | 1.79d |
| FC metabolizable, % | 61.01a | 64.33a | 58.56a | 56.71a | 6.70c | 36.44b | 35.95b |
| Energía metabolizable (EM), kcal día-1 | 328.0a | 329.5a | 304.7b | 305.6b | 106.16d | 131.61c | 130.80c |
| EM, % | 82.71a | 83.09a | 80.47a | 80.72a | 56.08 c | 69.53b | 69.10b |
| EM, kcal g-1 de dieta | 3.28a | 3.29a | 3.05a | 3.05a | 2.12c | 2.63b | 2.62b |
*Letras distintas en un mismo renglón indican diferencias significativas (Tukey, p < 0.05).
La EMA de la dieta ofrecida los días 1 a 2 produjo el siguiente valor de EMA: 3.29 kcal g-1 y la de los días 3 y 4 resultó ser de 3.05 kcal g-1, valores estadísticamente iguales (p > 0.05). Mientras que la EMA en kcal día-1 y la FC en g día-1 disminuyeron del día 1 al 7, el extracto etéreo metabolizable en g día-1 aumentó del día 1 al 4, y posteriormente disminuyó (p < 0.05).
Efectos del factor genotipo
En la Tabla 3 se muestran los efectos del factor genotipo, los gallos Criollos superaron a los Sasso en proteína cruda metabolizable, tanto en g día-1 como en %. Contrariamente, el genotipo comercial metabolizó mejor el extracto etéreo, o los lípidos en g día-1.
Tabla 3 Efectos del genotipo en la energía y los demás nutrientes metabolizables.
| Genotipo | ||
|---|---|---|
| Variable | Criollo | Sasso |
| Proteína cruda (PC) metabolizable, g día-1 | 9.13a* | 5.84b |
| PC metabolizable, % | 47.06a | 26.99b |
| Extracto etéreo (EE) metabolizable, g día-1 | 0.99b | 1.12a |
| EE metabolizable, % | 78.88 | 79.12 |
| Fibra cruda (FC) metabolizable, g día-1 | 3.95 | 3.66 |
| FC metabolizable, % | 44.34 | 42.32 |
| Energía metabolizable (EM), kcal día-1 | 222.48 | 217.76 |
| EM, % | 73.38 | 73.28 |
| EM, kcal g-1 de dieta | 2.83 | 2.80 |
*Letras distintas en un mismo renglón indican diferencias significativas (Tukey, p < 0.05).
Efectos del factor consumo
En la Tabla 4 se muestran los efectos del factor consumo de alimento. Todas las variables metabolizables de respuesta fueron mayores (p < 0.05) cuando el consumo fue de 100 g día-1 que cuando fue de 50 g día-1.
Tabla 4 Efectos del consumo en la energía y los demás nutrientes metabolizables.
| Consumo, g día-1 | ||
|---|---|---|
| Variable | 100 | 50 |
| Proteína cruda (PC) metabolizable, g día-1 | 13.00a* | 1.98b |
| PC metabolizable, % | 56.57a | 17.49b |
| Extracto etéreo (EE) metabolizable, g día-1 | 1.36a | 0.75b |
| EE metabolizable, % | 83.25a | 74.76b |
| Fibra cruda (FC) metabolizable, g día-1 | 6.30a | 1.31b |
| FC metabolizable, % | 60.29a | 26.36b |
| Energía metabolizable (EM), kcal día-1 | 317.38a | 122.86b |
| EM, % | 81.75a | 64.90b |
| EM, kcal g-1 de dieta | 3.17a | 2.46b |
*Letras distintas en un mismo renglón indican diferencias significativas (Tukey, p < 0.05).
Interacciones entre los factores genotipo×consumo
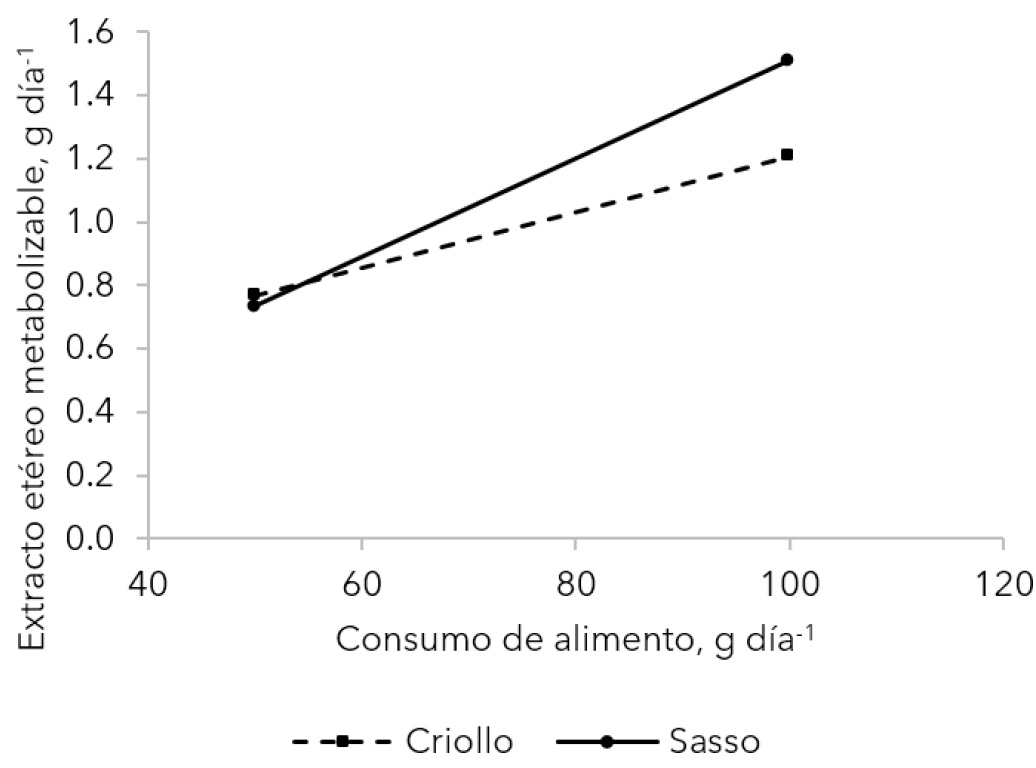
La interacción de estos dos factores solo fue significativa (p < 0.05) en la variable extracto etéreo metabolizable (g día-1). Esta variable fue similar entre genotipos cuando el consumo de alimento fue 50 g día-1, pero cuando fue de 100 g día-1, el genotipo Sasso retuvo o metabolizó más gramos de extracto etéreo por día (Figura 1). En esta figura también se observa que cuando los gallos consumieron 50 g día-1 el extracto etéreo metabolizable fue menor (p < 0.05) que cuando consumieron 100 g día-1 de alimento.
Discusión
Efectos del factor tiempo
Al medir EMA y EMAn, Macie et al. (2020) reportaron que las aves de mayor edad son más eficientes en la utilización de la energía de la dieta. En el presente estudio, todas las aves fueron adultas por lo que la disminución de los nutrientes metabolizables no se debió a la edad. Respecto a los tiempos para la recolección de excretas, Schang y Hamilton (1982) recomiendan 3 días para ensayos de EMV y 2 días para ensayos de EMA, y sugieren un periodo de recolección de excretas de 2 días para obtener mediciones precisas. En el presente estudio, la EMA en kcal día-1 disminuyó (p < 0.05) a partir del día 3 de medición (Tabla 2). Esto probablemente se debió a que la dieta ofrecida los días 1 y 2 resultó con menos cenizas (7.77%) que la ofrecida los días 3 y 4 (8.01%) y como es sabido, las cenizas no contienen energía (Usman et al. 2020).
Los mayores valores de los nutrientes metabolizables los días 1 a 4 con respecto a los de los días 5 a 7, se deben a que durante este último periodo experimental los gallos tenían más manejo que en el primero y como es sabido el manejo causa estrés (Adhikari et al. 2020) que a su vez disminuyó la EMA y otros nutrientes metabolizables. El aumento de extracto etéreo metabolizable (g día-1), el día 3, probablemente se debe a que el animal tiende a compensar el estrés de la falta de energía (kcal día-1) aumentando su capacidad de metabolizar lípidos e incrementando su habilidad para retenerlos. Bartz et al. (2018) mencionan que el estrés calórico disminuye el desarrollo productivo y Li et al. (2024) mencionan que aumenta la deposición de lípidos. En este estudio no se produjo un estrés calórico si no un estrés de manejo. Los lípidos producen colesterol, que es el precursor de corticosterona en la corteza adrenal (Ieko et al. 2019) y cuyos niveles aumentan en pollos estresados durante más de 60 segundos (Bedanova et al. 2007, Scanes et al. 2020). No obstante, este primer aumento de lípidos metabolizables entre los días 1 a 4, de los días 5 al 7 se perdió mucho de la capacidad para metabolizar los nutrientes evaluados (Tabla 2). Al respecto, Wu et al. (2020) recomiendan usar el mismo tipo de aves cuando se miden EMA de ingredientes. En este trabajo se usó el mismo gallo en días consecutivos. Mientras que Sibbald y Morse (1982) recomiendan un gallo por jaula. A partir de los resultados del presente estudio y en concordancia con Schang y Hamilton (1982) y Li et al. (2024), quienes recomiendan un periodo de 2 días, se recomienda realizar el ensayo de EMA y de los demás nutrientes metabolizables, entre el primero y el cuarto día consecutivos después de la adaptación de los gallos a la dieta, de preferencia los días 1 y 2, para minimizar el estrés del manejo.
Efectos del factor genotipo
La mayor proteína cruda (en g día-1 y %) y el menor extracto etéreo metabolizable (en g día-1) de los Criollos respecto a los gallos Sasso (Tabla 3), coinciden con Paredes y Vásquez (2020), quienes indicaron que la genética del ave criolla incrementa el rendimiento productivo, y que la grasa abdominal fue mayor en el genotipo comercial que en el Criollo. Por su cercanía genética con el ave roja de la jungla, ancestro de las aves domésticas, los gallos criollos utilizaron con mayor eficiencia la proteína que la grasa del alimento. Comparadas con sus correspondientes genotipos comerciales, se ha demostrado que el ave roja de Malasia posee mayor concentración de espermatozoides (Malik et al. 2013), que las aves nativas de Tailandia poseen menos grasa, colesterol y triglicéridos (Chaiwang et al. 2023), y que las de México producen mayor peso de yema y espesor de cascarón (Sosa-Montes et al. 2022).
Debido a su rusticidad bien conocida (Adamu et al. 2019), los gallos Criollos mostraron alta disponibilidad de proteína para formar anticuerpos y poder sobrevivir (Solanki y Devi 2020), y así superar el estrés del manejo durante todos los días de evaluación. Por lo que, el genotipo Criollo mostró mayor capacidad para utilizar la proteína que el Sasso (Tabla 3).
Efectos del factor consumo
El consumo de 100 g día-1 produjo valores más cercanos a los de la literatura consultada. Por ejemplo, usando pollos de engorda, Lasek et al. (2020) reportaron valores de proteína cruda metabolizable para grano de maíz que fueron más altos (76% contra 56.57% con 100 g día-1 de consumo del presente estudio), esto es debido probablemente a que la dieta (Tabla 1) contenía 50% de trébol. Estos mismos autores reportaron, 57.5% de extracto etéreo metabolizable, valor más bajo que el del presente estudio que fue 83.25% con 100 g día-1 de consumo (Tabla 4). Santos et al. (2015) reportaron valores de 90.4% como proteína y 93.3% como extracto etéreo metabolizables del maíz. El mayor valor de lípidos metabolizables (en cal día-1 y en %, Tabla 4) cuando los gallos consumieron 100 g día-1 de alimento (con respecto a 50 g día-1) probablemente incrementó la EMA, que a su vez aumentó los valores de los demás nutrientes metabolizables (p < 0.05). De igual forma, Alvarenga et al. (2013) mencionan que el efecto extra calórico de los lípidos consumidos aumenta la disponibilidad de los nutrientes del alimento.
Ante un consumo tan reducido de 50 g, la utilización de la energía y de todos los demás nutrientes se vio comprometida. Esto concuerda con Sibbald (1980) quien recomienda para los bioensayos, que los gallos adultos no consuman menos de 30 g, ya que ello disminuiría la energía metabolizable aparente en kcal g-1. Sibbald y Morse (1982) usaron niveles crecientes de consumo de alimento 10 a 70 g por animal por día.
La AME depende de la edad, la especie y el consumo de alimento (Barzegar et al. (2020). Sibbald (1975) informó que la EMA se aproxima a la EMV a medida que aumenta el consumo de los gallos adultos. Esta aproximación de la EMA a la EMV se demuestra fácilmente tomando límites a la ecuación de Sibbald (1976) y mencionada por Wu et al. (2020):
Donde EMV = EMV del alimento, GEex = calor de combustión de la excreta, GEen = calor de combustión de las pérdidas endógenas, CA = consumo de alimento.
Al tomar el límite, cuando CA se hace muy grande el término GEen/CA se aproxima a cero por lo que EMV = (GEal − GEex)/CA = EMA, es decir, la EMV resulta igual a la EMA. Contrariamente, cuando el consumo tiende a cero, predomina el valor -GEex que hace negativa a la EMA. Lo primero fue demostrado experimentalmente por Sibbald (1975) y por Wolynetz y Sibbald (1984), lo segundo se prueba experimentalmente en el presente estudio. Esto se puede generalizar a cualquier tipo de ingrediente metabolizable, no solo a la energía, si el consumo es bajo o se aproxima a cero, los nutrientes metabolizados de una dieta tienden a ser cero o negativos. Por ejemplo, es común hablar de balance negativo de nitrógeno (Von-Bobrutzki et al. 2013) en que la retención de nitrógeno es negativa cuando el consumo de alimento se hace cero. En el presente estudio, algunos valores de nutrientes metabolizables produjeron valores negativos en el día 5, cuando inició el consumo de 50 g día-1. Posteriormente los valores de todas las variables fueron bajos pero positivos, probablemente debido a un proceso de adaptación de los animales al bajo consumo de alimento.
Interacciones entre los factores genotipo×consumo
La interacción de estos dos factores solo fue significativa (p < 0.05) en la variable extracto etéreo metabolizable (g día-1). Esta interacción significa que el genotipo Sasso retiene o metaboliza más gramos de extracto etéreo cuando el consumo de alimento es de 100 g día-1, esta variable fue similar entre genotipos cuando el consumo de alimento fue 50 g día-1 (Figura 1). Probablemente, el ave comercial tiende a depositar lípidos (Paredes y Vásquez 2020) para adaptarse al estrés del manejo, mientras que el ave criolla, por su capacidad de sobrevivencia o rusticidad (Adamu et al. 2019) tiende a depositar proteínas (Tabla 3).
El consumo de cada ave fue 100 o 50 g, todas fueron del mismo sexo, consumieron la misma dieta (Tabla 1), tenían edades y pesos similares. Por tanto, solo las diferenciaba el genotipo, el día de medición y el consumo. La energía metabolizable de los ingredientes o del alimento es muy importante y existen muchos trabajos en la literatura que la evalúan. Sin embargo, el porcentaje de utilización de los nutrientes que se encuentran en los ingredientes (energía o nutrientes metabolizados) ha sido menos estudiado.
Conclusiones
Los gallos Criollos mexicanos mostraron mejor capacidad que los Sasso para metabolizar la proteína cruda tanto en g día-1 como en %, contrariamente, los gallos Sasso mostraron mayor capacidad para metabolizar la grasa en g día-1. En cuanto al consumo de alimento de 100 g día-1 se obtuvieron mayores valores de nutrientes metabolizables que con el consumo de 50 g día-1, por lo que consumos aún menores de alimento, pueden producir valores muy bajos o negativos. Al avanzar los días de evaluación, todas las variables disminuyeron, ya que de los días 1 a 4 se produjeron mayores valores de casi todas las variables de respuesta en comparación con el tiempo de medición de 5 a 7 días. Por tanto, se recomienda hacer la medición de energía y nutrientes metabolizables de las dietas, los dos primeros días después del proceso de adaptación y entrenamiento, y que los gallos consuman por lo menos 100 g de alimento por día.

