











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
La intensificación de actividades antropogénicas trae como consecuencia la fragmentación del paisaje, pérdida del hábitat y biodiversidad (Laurance et al. 2002). Ejemplos como la construcción y operación de carreteras y autopistas son actividades antropogénicas que generan impactos directos sobre ecosistemas y las especies que lo componen, ya que origina la fragmentación de hábitat, cambios microclimáticos, promueve la dispersión de especies exóticas, produce material contaminante del agua, suelo, aire, además de generar ruido por el flujo vehicular continuo. El efecto más fácil de observar en una autopista es la muerte de individuos de fauna silvestre de diferentes especies por atropellamiento de vehículos (González-Gallina et al. 2013, González-Gallina y Benítez-Badillo 2013, Myczko et al. 2017). La identificación de dichos efectos sobre la comunidad de quirópteros ha sido poco explorada en zonas tropicales y, su estudio, es la base para el diseño de estrategias que eviten, mitiguen o compensen estos impactos (Arroyave et al. 2006).
Los murciélagos son un grupo vulnerable ante los efectos de los desarrollos viales (Altringham 2008, Russell et al. 2009, Myczko et al. 2017). Por ejemplo, las carreteras representan espacios sin protección ante depredadores de murciélagos (Ciechanowski et al. 2007); es decir, los murciélagos perciben a los vehículos como una amenaza, lo que causa que tomen medidas evasivas derivadas de un comportamiento anti depredatorio (Clark et al. 2001). Además, las carreteras son barreras físico-geográficas que dificultan el acceso a sitos de alimentación, descanso, refugio para la crianza o apareamiento (Berthinussen y Altringham 2012a, b) y obliga a los murciélagos a desplazarse hasta encontrar sitios de cruce apropiados, lo que implica un gasto energético mayor (Kerth y Melber 2009). Forrajear cerca de carreteras limita la capacidad de ecolocalización eficaz de los murciélagos debido al ruido generado por los vehículos (Schaub et al. 2008) y, en consecuencia, se manifiestan cambios en el comportamiento forrajero (Waters et al. 1999, Luo et al. 2015). Efectos que se reflejan en la variación de la estructura del ensamblaje de murciélagos, siendo las especies que presentan vuelos altos y especializados en alimentarse en áreas abiertas, las que se adaptan a estas perturbaciones (Jones 1995).
La frecuencia de coalición de murciélagos por vehículos varía por especie, gremio trófico y época del año (Russell et al. 2009), siendo perjudicial para especies de murciélagos con bajas tasas de reclutamiento poblacional (Lesinski 2007) y posiblemente sea causa de la extinción local (Mickleburgh et al. 2002). En otro contexto, ciertas especies de murciélagos se benefician al aprovechar como forraje a los insectos con fototaxis positiva a las luces de los vehículos en autopistas (Rydell 2006, Jung y Kalko 2010). Sin embargo, en selvas tropicales de México se cuenta con poca evidencia del efecto de las autopistas sobre la comunidad de quirópteros (Gaisler et al. 2009, Lesinski et al. 2011), en este sentido, es importante evaluar estos efectos como indicador de disturbios en la comunidad de murciélagos (Slater 2002, Stone et al. 2009).
En México se ha estudiado el efecto negativo que los desarrollos viales provocan sobre la comunidad de vertebrados (González-Gallina y Benítez-Badillo 2013, Cervantes-Huerta et al. 2017). Una manera indirecta de estudiar el efecto que generan las autopistas (principalmente impacto por el ruido e intensidad lumínica emitida por los vehículos) sobre la estructura de la comunidad de murciélagos (Van der Ree et al. 2011), es mediante la evaluación de la riqueza y abundancia de murciélagos que habitan o usan las coberturas del paisaje del área de estudio (Peter et al. 2013, Arroyave et al. 2014). Otra forma es mediante la representatividad de los gremios tróficos. Por ejemplo, murciélagos generalistas de hábitat con amplitud de nicho trófico, estarán mejor representadas en paisajes abiertos y perturbados; en comparación con murciélagos con hábitos alimenticios especializados, las cuales serán más raros en paisajes abiertos y más comunes en paisajes con presencia de masas de bosque denso (Medellín et al. 2000, Adán et al. 2007, Pardini et al. 2009). Por lo anterior, los objetivos del presente estudio fueron: i) describir la estructura de la comunidad de murciélagos y ii) relacionar la riqueza y abundancia de la comunidad de murciélagos con respecto al flujo vehicular, en una localidad con presencia del tipo de vegetación denominada selva mediana subperennifolia en la zona centro del estado de Veracruz, México.
MATERIALES Y MÉTODOS
Área de estudio y descripción de los sitios de muestreo
El estudio se realizó en el predio del Colegio de Postgraduados, Campus Córdoba, ubicado en el municipio de Amatlán de Los Reyes, Veracruz, México (18◦ 51’ 20.52” LN, 96◦ 51’ 37.59” LO), el cual se encuentra dividido por la autopista Córdoba-Veracruz. En la mayor parte del municipio se cuenta con vegetación denominada selva mediana subperennifolia (Cházaro-Basáñez 1992). En el año de 1962 la autopista fue construida por lo que la vegetación original quedó divida en dos o más fragmentos. La autopista está construida en los extremos transversales con taludes de 3 m de altura en promedio, con presencia de plantas tipo ruderal y especies representativas de la selva. La autopista es la única vía de comunicación terrestre de cuota que conecta el centro con el sur y sureste de México.
En el área de estudio, que correspondió al Km 11 de la autopista Córdoba-Veracruz, tres sitios de muestreo fueron establecidos: sitio 1 (fragmento de selva mediana subperennifolia) a 30 m (18◦ 51’ 47.50” LN, 96◦ 51’ 42.24” LO), sitio 2 (cultivo de café bajo sombra) a 80 m (18◦ 51’ 54.20” LN, 96◦ 51’ 34.03” LO) y sitio 3 (pastizal con presencia de vegetación secundaria) a 120 m (18◦ 51’ 45.06” LN, 96◦ 51’ 35.47” LO) de distancia transversal a la autopista. La distancia entre sitios de monitoreo fue de 250 m en promedio, abarcando las coberturas del paisaje (Ávila-Torresagatón et al. 2012). El área de estudio se eligió debido a que a 3.1 km en línea recta, se encuentra un refugio (cueva o caverna) de murciélagos en la localidad denominada Cerro de Lourdes, municipio de Amatlán de Los Reyes, Veracruz, México (18◦ 50’ 53.50” LN, 96◦ 53’ 10.08” LO).
Caracterización del flujo vehicular
El flujo vehicular de la autopista se solicitó a la Secretaría de Comunicaciones y Transportes de México, de la caseta de peaje “Cuitláhuac”(folio 912000003118 y 912000008518), localizada a 12.5 km del área de estudio (18◦ 50’ 21.98” LN, 96◦44’ 49.01” LO). De los valores mensuales, se obtuvo el 60% de pases vehiculares que ocurrieron entre las 19:00 y 23:00 h (Tseng et al. 2014) y este se dividió entre 30 días y se multiplicó por tres (que correspondieron a los tres días de muestreo), de tal modo que se obtuvo el número de pases de vehículos mensuales para octubre (21 737), noviembre (22 488), diciembre (27 177), enero (24 253), febrero (20 209) y marzo (25 283).
Características del muestreo y captura de murciélagos
El trabajo de campo se realizó de octubre de 2017 a marzo de 2018, efectuando muestreos mensuales con duración de tres noches consecutivas de monitoreo en horario de 18:00 a 00:00 h (Calderón-Patrón et al. 2013). Los muestreos se realizaron en días con distinta fase lunar, pues en otros estudios se ha determinado que la fase lunar no afecta la frecuencia de actividad de los murciélagos, pues no existe evidencia de fobia lunar (Pech-Canché et al. 2018, Musila et al., 2019). De las 17:00 a 17:30 h se realizó un recorrido de 1 km sobre las orillas de los dos carriles de la autopista con el fin de encontrar rastros de murciélagos atropellados.
En cada sitio de monitoreo se colocaron tres (a manera de réplica) redes de niebla, para los sitios 1 y 2 fueron dos redes de 12 x 2.5 m y una red de 9 x 2.5 m, y para el sitio 3 se instaló una red de 12 x 2.5 m y dos redes de 9 x 2.5 m. Las redes se revisaron cada 30-45 min para liberar a los murciélagos y depositarlos en una bolsa de manta hasta su evaluación (Tschapka 1998, Briones et al. 2005).
Cada murciélago capturado se pesó (g), determinó el sexo, edad y condición reproductiva; además de que se identificó a nivel de especie con la clave taxonómica de Medellín et al. (2008) y se clasificó con base en su gremio trófico (Giannini y Kalko 2004); para marcarlo con barniz de uñas para identificar recapturas y evitar dobles conteos, y se liberó.
Evaluación del muestreo y cobertura de la muestra
Para evaluar el esfuerzo de muestreo (Ĉm) en cada uno de los sitios de monitoreo, se aplicó la ecuación: Ĉm = (A) (h) (d) (m), donde A = área (m2) total de las redes; h = promedio de horas de muestreo; d = total de días muestreados y m = total de muestreos realizados (Briones et al. 2005). Para evaluar la cobertura de la muestra (Ĉn) en cada sitio, con los datos de la abundancia observada se construyeron las curvas de acumulación de especies bajo el modelo completitud basado en la muestra, se calcularon sus respectivos intervalos de confianza al 95% y se usaron para identificar diferencias cuando estos no se traslaparon entre las unidades muestrales (Cumming et al. 2007). Este análisis se realizó en el software iNext (Chao et al. 2016), con el fin de comparar la riqueza y abundancia de especies entre las unidades muestrales con una Ĉn similar (Jost 2006, Jost 2010, Chao et al. 2014).
Análisis de la estructura de la comunidad de murciélagos
La estructura de la comunidad de murciélagos fue descrita mediante los números de Hill (1973) del orden 0 (q0 = riqueza de especies), orden 1 (q1 = exponencial del índice de Shannon); orden 2 (q2 = inverso del índice de Simpson) para el periodo del monitoreo y por tipo de cobertura muestreada (Jost 2006). Comparamos estas medidas entre los 3 sitios de monitoreo a partir de la cobertura de la muestra y sus respectivos intervalos de confianza calculados al 95%, en el cual se asumen diferencias significativas cuando estos no se superponen (Cumming et al. 2007). Estos análisis se realizaron en el software iNext (Chao et al. 2016). Con el propósito de determinar el efecto del tráfico vehicular con respecto a la variación de la riqueza y abundancia mensual de la comunidad de murciélagos, se aplicó un análisis de regresión entre la riqueza y la abundancia mensuales con respecto al flujo vehicular mensual (Zar 2010).
RESULTADOS
Con un total de 17 noches de monitoreo (una noche no se monitoreó por que las condiciones extremas del clima no lo permitieron), se obtuvo una Ĉm = 6480, 4860 y 7830 h/red para el sitio 1, 2 y 3 respectivamente, que sumaron en conjunto una Ĉm = 19 170 h/red para el muestreo global.
Los valores de la Ĉn fueron superiores al 80% de completitud del muestreo (Sitio1 Ĉn = 0.87, Sitio 2 Ĉn = 0.91, Sitio 3 Ĉn = 0.97), sin diferencias significativas entre sitios de monitoreo. Las medidas de diversidad del número efectivo de especies del orden 0, 1 y 2 (q0, q1 y q2) no fueron significativamente diferentes entre sitios de monitoreo, ya que los intervalos de confianza se superponen (Figura 1). Esto significa que la muestra de la comunidad de murciélagos fue homogénea en cuanto a riqueza y abundancia de especies (Figura 2). Los recorridos realizados en las orillas de la autopista no evidenciaron la presencia de quirópteros atropellados.
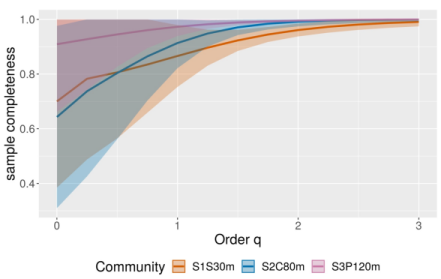
Figura 1: Evaluación de la completitud de la muestra (Ĉn) con base en las medidas de diversidad del orden 0, 1 y 2 (q0, q1 y q2). Las líneas sombreadas asociadas a la Ĉn representan los intervalos de confianza calculados al 95% de confiabilidad, los cuales se traslapan. S1S30m = Sitio 1 con selva a 30 m, S2C80m = Sitio 2 con cafetal a 80 m, S3P120m = Sitio 3 Pastizal a 120 m.

Figura 2: Comparación de la riqueza y abundancia de la comunidad de murciélagos en tres sitios de monitoreo en el Km 11 de la autopista Córdoba-Veracruz, México. Líneas sombreadas representan los intervalos de confianza al 95%. S1S30m = Sitio 1 con selva a 30 m, S2C80m = Sitio 2 con cafetal a 80 m, S3P120m = Sitio 3 Pastizal a 120 m.
Se capturaron 127 individuos (y una recaptura de C. sowelli) pertenecientes a dos familias, cuatro subfamilias, nueve géneros y 12 especies. Las especies de murciélagos presentaron abundancias mensuales que van desde un individuo como Dermanura azteca, Mormoops megalophylla y Pteronotus personatus hasta 36 individuos como Artibeus jamaicensis (Tabla 1). De las especies de murciélagos capturadas, ocho se clasificaron en el gremio frugívoro (75%), tres pertenecen al gremio insectívoro (17%) y solo una al gremio nectarívoro (8%) (Tabla 1).
Tabla 1 Listado taxonómico, gremios tróficos, riqueza y abundancia mensual de la comunidad de murciélagos en un ambiente tropical antropizado. Abd = abundancia; O = octubre, N = noviembre, D = diciembre, E = enero, F = febrero, M = marzo.
| Familias | Subfamilias | Géneros y especies | Gremio | O | N | D | E | F | M | Abd Absoluta | Abd Relativa |
| Phyllostomidae | Stenodermatinae | Artibeus jamaicensis (Leach, 1821) | Frugívoro | 1 | 6 | 14 | 1 | 6 | 8 | 36 | 28.3 |
| Artibeus lituratus (Olfers, 1818 | Frugívoro | 0 | 8 | 1 | 1 | 0 | 0 | 10 | 7.9 | ||
| Sturnira lilium (Geoffroy, 1810) | Frugívoro | 0 | 9 | 4 | 1 | 1 | 2 | 17 | 13.4 | ||
| Sturnira ludovici (Anthony, 1924) | Frugívoro | 2 | 5 | 1 | 3 | 0 | 0 | 11 | 8.6 | ||
| Platyrrhinus helleri (Peters, 1866) | Frugívoro | 0 | 0 | 0 | 2 | 0 | 0 | 2 | 1.6 | ||
| Dermanura azteca (Andersen, 1906) | Frugívoro | 0 | 3 | 1 | 0 | 0 | 0 | 1 | 0.8 | ||
| Carolliinae | Carollia perspercillata (Linnaeus, 1758) | Frugívoro | 4 | 11 | 0 | 0 | 0 | 0 | 7 | 5.5 | |
| Carollia sowelli (Baker et al., 2002) | Frugívoro | 1 | 2 | 12 | 4 | 2 | 4 | 34 | 26.8 | ||
| Glossophaginae | Glossophoga soricina (Pallas, 1766) | Nectarívoro | 0 | 2 | 1 | 0 | 0 | 1 | 4 | 3.1 | |
| Mormoopidae | Micronycterinae | Lampronycteris brachyotis (Dobson, 1879) | Insectivoro | 0 | 0 | 1 | 0 | 0 | 0 | 3 | 2.4 |
| Mormoops megalophylla (Peters, 1864) | Insectivoro | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0.8 | ||
| Pteronotus personatus (Wagner, 1843) | Insectivoro | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0.8 | ||
| Riqueza mensual (S) | 5 | 8 | 8 | 7 | 3 | 4 | 127 | 100% | |||
| Abundancia mensual (Abd) | 9 | 46 | 35 | 13 | 9 | 15 |
La riqueza y abundancia de especies de murciélagos por sitio de monitoreo fue variable, pues el 58 % (siete especies) se registraron en el sitio 1 con vegetación de selva, en el sitio 2 con presencia de cultivo de cafetal estuvieron representadas el 75% de las especies de murciélagos (nueve especies), mientras que en el sitio 3 con pastizal y vegetación secundaria se registró la mayor riqueza (10 especies) y abundancia (72 individuos) de murciélagos (Tabla 2). Los resultados del análisis de regresión lineal indican una baja correlación no significativa entre la riqueza mensual (R2 = 0.27, T = 1.23, P = 0.28) y la abundancia mensual (R2 = 0.13, T = 0.76, P = 0.48) con respecto al tráfico vehicular; sin embargo, la curva de regresión muestra una relación que indica que a mayor distancia de la autopista se registró el mayor número de especies de murciélagos (Figura 3).
Tabla 2 Características de la riqueza y abundancia de la comunidad de murciélagos en los tres sitios de monitoreo. Abd = Abundancia, S = riqueza de especies.
| Sitio 1 Selva 30 m | Abd | Sitio 2 Cafetal 80m | Abd | Sitio 3 Pastizal a 120m | Abd |
| A. jamaicencis | 6 | A. jamaicencis | 14 | A. jamaicencis | 16 |
| A. lituratus | 1 | A. lituratus | 3 | A. lituratus | 6 |
| C. perspicillata | 2 | C. perspicillata | 3 | C. perspicillata | 2 |
| C. sowelli | 8 | C. sowelli | 6 | C. sowelli | 17 |
| - | 0 | - | 0 | G. soricina | 4 |
| - | 0 | L. brachyotis | 2 | L. brachyotis | 2 |
| P. personatus | 1 | - | 0 | - | 0 |
| - | - | P. helleri | 1 | P. helleri | 1 |
| S. lilium | 1 | S. lilium | 1 | S. lilium | 16 |
| S. ludovici | 2 | S. ludovici | 3 | S. ludovici | 7 |
| - | D. aztecus | 1 | - | 0 | |
| - | 0 | - | 0 | M. megalophylla | 1 |
| S = 7 | Abd = 21 | S = 9 | Abd = 34 | S = 10 | Abd = 72 |

DISCUSIÓN
El presente trabajo contribuye al conocimiento de la diversidad local de 12 especies de murciélagos, además de aportar información de la zona centro del estado de Veracruz. La mayor diversidad de murcielagos capturados pertenece a la subfamilia Stenodermatinae, lo cual fue reportado para regiones con cultivos de café del centro del estado de Veracruz (Saldaña-Vázquez et al. 2010). De acuerdo con las curvas de acumulación de especies, no se completó la riqueza total esperada por la falta de un mayor esfuerzo de muestreo, pues se requiere seguir haciendo muestreos para alcanzar la completitud cercana al 100% de la riqueza esperada. Por lo que el uso de los detectores acústicos en combinación con un mayor número de redes de niebla y sitios de monitoreo podría ayudar a registrar un mayor número especies de murciélagos de otros gremios como los insectívoros, los cuales vuelan a mayores alturas (García-García y Santos-Moreno 2009).
Se identificaron 12 especies de murciélagos de dos familias: Phyllostomidae y Mormoopidae. Las cuales representan el 13.5% de murciélagos reportados para el estado de Veracruz de 89 especies (González-Christen y Delfín-Alfonso 2016). Las especies de murciélagos registradas en este estudio ya han sido reportadas para áreas con cultivo de café (Saldaña-Vázquez et al. 2010), bosque mesófilo de montaña (Pineda et al. 2005), pero para vegetación de selva mediana subperennifolia en el centro de Veracruz, no han sido reportadas. Dos de los sitios de monitoreo completaron más del 90% de la representatividad de la riqueza de especies esperadas, excepto el sitio 1, donde solo se cubrió aproximadamente el 80% de la riqueza esperada; lo que indica que la riqueza, el número de especies comunes y de especies poco comunes fue mayor en el sitio de muestreo más alejado de la autopista (Figura 3).
La mayor abundancia mensual la presentaron A. jamaicensis y C. sowelli. Especies como D. azteca, M. megalophylla y P. personatus fueron especies representadas por un individuo. Por tipo de cobertura, G. soricina y M. megalophylla sólo se capturaron en el pastizal, P. personatus en la selva y D. aztecus en el cafetal. Mientras que especies como L. branchyotis y P. helleri se capturaron en dos tipos de cobertura (cafetal y pastizal), el resto de las especies estuvieron representadas en los tres tipos de cobertura. Este patrón de abundancia puede ser explicado por la adaptación de los murciélagos a la constante interferencia humana y al distinto grado de perturbación de los fragmentos de los tipos de vegetación del paisaje en estudio (Lesinski 2007, Jung y Kalko 2010).
La variabilidad en la abundancia de la comunidad de murciélagos puede deberse al tipo de forrajeo. Las especies de murciélagos con mayor abundancia (A. jamaicensis y C. sowelli ) son indicadoras de un forrajeo en hábitats abiertos, mientras que las menos abundantes (A. lituratus, D. aztecus, M. megalophylla, P. personatus, P. helleri y S. lilium) forrajearon en los sitios con mayor diversidad vegetal (Fenton et al. 1992, Mena 2010). Es decir, independientemente del grado de perturbación, los murciélagos oportunistas estuvieron presentes en los tres tipos de vegetación (Medellín et al. 2000), mientras que, los murciélagos con bajas abundancias se capturaron en los sitios donde la vegetación cuenta con mayor diversidad florística como la selva y el cafetal (Jung y Kalko 2010, Russo y Ancillotto 2015).
La comunidad de murciélagos bajo estudio estuvo compuesta por ocho especies del gremio frugívoro, tres del insectívoro y una especie nectarívora. Evidentemente, la presencia de las especies que componen estos gremios tróficos se adaptan a cambios en los ecosistemas producidos por el hombre; sin embargo, la ausencia de especies de murciélagos del gremio omnívoro y hematófago en el muestreo, pueden ser un indicador de recursos disponibles escasos para estas especies (Kraker-Castañeda y Echeverría-Tello 2012). La colisión con vehículos es la principal causa que afecta a la fauna silvestre debido a que aumentan las tasas de mortalidad, fungen como barreras geográficas artificiales y pueden ser un impedimento físico para la migración o dispersión de las especies (Monge-Nájera 1996). El efecto de las carreteras sobre los murciélagos parece variar dependiendo el grupo de murciélagos, hábitos forrajeros y época del año (Russell et al. 2009) y se ha demostrado que un número considerable de muertes de murciélagos son causadas por colisiones con vehículos (Lesinski 2007, Gaisler et al. 2009, Russell et al. 2009, Lesinski et al. 2011), pero este estudio no generó tal evidencia. El efecto del flujo vehicular sobre la comunidad de murciélagos puede deberse al efecto del periodo invernal en el que se realizó el monitoreo (Berthinussen y Altringham 2012a), al comportamiento forrajero de los murciélagos (Myczko et al. 2017), y a la construcción de la autopista como los puentes vehiculares superiores e inferiores, obras de drenaje y taludes de 3 m con vegetación nativa (Cervantes-Huerta et al. 2017), pueden ser factores que influyen en el mantenimiento de la estructura de la comunidad. En estudios futuros se pueden considerar el tipo de carretera, horario de mayor flujo vehicular y relacionarlos con la actividad de los murciélagos para conocer los efectos a menor escala (Myczko et al. 2017).
Se detectó un posible efecto de borde que debe tomarse con cautela, que sugiere un efecto de barrera (Russell et al. 2009), lo cual debe determinarse en estudios futuros. Pero los valores de riqueza y abundancia de especies de murciélagos se concentraron en el sitio de muestreo más alejado de la autopista con pastizal y vegetación secundaria, donde posiblemente es un espacio con menor perturbación antrópica (Van der Ree et al. 2011). En contraste con el sitio de muestreo 1 ubicado a 30 m de la autopista, un área con un remanente de selva mediana subperennifolia perturbada con mayor intensidad de ruido y luz vehicular, se tuvo una menor riqueza y abundancia de especies de murciélagos (Abbott et al. 2012). Estos efectos de borde no fueron reportados por Abbott et al. (2012), ya que algunas a especies de murciélagos optan por realizar vuelos bajos y cruzan en sitios donde los árboles maduros no han sido talados. Pero debido al número de murciélagos que cruzan, se tiene mayor posibilidad de colisión, debido a que la mayoría de las especies de murciélagos vuelan a velocidades bajas (< 20 km/h) y entre 0 y 4 m del suelo (Russell et al. 2009, Berthinussen y Altringham 2012b).
Algunos murciélagos parecen tolerar las urbanizaciones y aprovechan la presencia de luces artificiales, que atraen insectos, por lo que los usan como sitios de forrajeo (Jung y Kalko 2010). También algunos faros de luz atraen mayor cantidad de insectos, de modo que especies insectívoras como Pipistrellus pipistrellus utilizan con mayor intensidad el espacio aéreo próximo (Adán et al. 2007). Mientras que las especies Barbastella barbastellus (Myczko et al. 2017) y Nyctalus noctula (Bogdanowicz et al. 1999) del gremio insectívoro utilizan caminos asfaltados como sitios de forrajeo y se alimentan a alturas donde el ruido es menor (Bogdanowicz et al. 1999). Por lo que el desarrollo y la intensificación urbana como las autopistas, puede tener efectos positivos o negativos sobre las poblaciones de murciélagos, principalmente del gremio insectívoro (Sánchez 2011). La luz artificial de los vehículos puede ejercer cambios en la estructura de la comunidad de murciélagos (Stone et al. 2009). Sin embargo, especies del gremio insectívoro que son atraídos por insectos con fototaxis positiva (Jung y Kalko 2010), pueden usar estos espacios como sitios de forrajeo (Van der Reed et al. 2011), a pesar de estar cercanos a construcciones vehiculares (Bogdanowicz et al. 1999, Adán et al. 2007).
El ruido es otro factor importante que modifica la distribución y abundancia de murciélagos en zonas perturbadas, afectando a especies del género Myotis (Jones y Rayner 1988, Bogdanowicz et al. 1999, Luo et al. 2015). Especies de murciélagos insectívoros con preferencia de hábitat como M. myotis, son conocidos por su dependencia de la escucha pasiva de los sonidos de sus presas al cazar y por lo tanto evita la contaminación acústica (Schaub et al. 2008, Siemers y Schaub 2011). En el presente estudio, se puede deducir que los murciélagos de esta zona eviten forrajear cerca de las carreteras y se alejen a zonas donde el ruido es menor y les permitan ecolocalizar de forma eficaz (Waters y Jones 1995).
No se encontraron evidencias sobre quirópteros atropellados, probablemente a la dificultad de encontrar sus restos, ya que, en los recorridos de la autopista, no se encontraron evidencias. Factores asociados a la dificultad de encontrar sus restos por su rápida descomposición (Grodsky et al. 2011, Medinas et al. 2013) o por el consumo por especies carroñeras u oportunistas (Slater 2002, Stone et al. 2009) como mapaches (Procyon lotor), zorras (Urocyon cinereoargenteus) y zopilotes (Coragyps atratus), especies observadas en campo, limitan la evidencia de restos (Lesinski 2007, Gaisler et al. 2009, Russell et al. 2009, Lesinski et al. 2011).
Los murciélagos representan un grupo vulnerable ante los desarrollos viales, debido a su baja fecundidad y la necesidad de grandes áreas con hábitat conservado (Altringham 2008), pero también es posible que la comunidad de murciélagos registrados, usen el espacio independientemente de la perturbación de la luz artificial y el ruido vehicular (Myczko et al. 2017). Los efectos que las carreteras pueden ejercer sobre la comunidad de murciélagos son difícilmente cuantificables (Sánchez 2011, Abbott et al. 2012). Por lo que se recomienda en estudios futuros medir la intensidad lumínica y el ruido en los sitios de monitoreo para correlacionar estas variables con la estructura de la comunidad de murciélagos (Medinas et al. 2013).
CONCLUSIONES
El flujo vehicular no afecta de manera significativa la estructura de la quiropterofauna presente en la región centro del estado de Veracruz, ya que la estructura de la comunidad de murciélagos no presentó diferencias entre el número efectivo de especies. En tres gremios tróficos se clasificaron las especies de murciélagos, consideradas especies generalistas de hábitat y tolerantes a la perturbación. La mayor riqueza y abundancia de murciélagos se concentró en el sitio más alejado de la autopista, lo que refleja un efecto borde debido a la respuesta de la comunidad de murciélagos ante el ruido e intensidad lumínica. Aunque no se encontraron evidencias del impacto de las autopistas sobre la comunidad de murciélagos, no debe minimizarse que este es un grupo vulnerable ante los desarrollos viales en ecosistemas selváticos, ya que un gran número de especies requieren de grandes áreas de hábitat o masas densas de bosque.

