











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La radiación gamma es un agente mutagénico que se utiliza para inducir variabilidad genética en programas de mejoramiento vegetal. Antes de iniciar un programa de mejoramiento genético por mutagénesis, es necesario establecer la dosis de radiación con mayor probabilidad de producir mutaciones efectivas (Ramchander et al. 2015, Rajarajan et al. 2016). Diversos investigadores concuerdan que la dosis donde muere el 50% (DL50) de los individuos irradiados, es donde existe mayor probabilidad de producir mutaciones útiles en un programa de mejoramiento genético (Golubinova y Gecheff 2011,Ángeles-Espino 2013). Mientras que otros investigadores señalan que otra dosis con alta probabilidad de producir mutaciones efectivas es donde se reduce el crecimiento en 50% (RG50) (Akgüm y Tosun 2004, Khalil et al. 2014). Lo que se debe a que con dosis bajas de radiación los impactos en el genoma son mínimos, por lo tanto los cambios fenotípicos también serán escasos. Por otro lado, con dosis altas el genoma sufre múltiples impactos que regularmente producen aberraciones o cambios negativos (Songsri et al. 2011, Thole et al. 2011).
El pasto africano (Eragrostis lehmanniana) es una especie invasora que afecta los pastizales del norte de México y se caracteriza por ser de alta prolificidad, capacidad de establecimiento y baja apetencia para el ganado (Flanders et al. 2006). Su hábito de crecimiento y rusticidad proveen al pasto africano de alta capacidad para invadir y dominar la composición de gramíneas en los pastizales donde se encuentra (Hupy et al. 2004). Sin embargo, estas características también convierten al pasto africano en una especie atractiva para rehabilitar pastizales con alto grado de degradación, donde otras especies no logran establecerse (McGlone y Huenneke 2004). Antes de utilizar esta especie es importante que se someta a un programa de mejoramiento genético para aumentar su aceptación por el ganado y disminuir su capacidad invasora (O’Reagain y Mentis 1989). No obstante, por ser una especie introducida en México, las poblaciones de pasto africano pudieran presentar una reducida variabilidad genética; por lo que la mutagénesis es una alternativa que puede generar variabilidad genética para el mejoramiento del pasto africano. Pero se desconoce la dosis con mayor probabilidad para producir mutaciones efectivas. Por lo que, el objetivo fue determinar la dosis letal media y la reducción media del crecimiento para determinar la dosis óptima de irradiación gamma e inducir mutagénesis en pasto africano.
MATERIALES Y MÉTODOS
El experimento se realizó en laboratorio e invernadero. Se evaluaron 10 dosis de irradiación en semilla de pasto africano, las cuales consistieron en la aplicación de 0 (tratamiento testigo), 100, 200, 300, 450, 600, 900, 1400, 2000 y 4000 Gray (Gy). La irradiación se realizó en el Complejo MOSCAMED de SENASICA-SAGARPA en Metapa de Domínguez, Chiapas, México, en colaboración con el Campo Experimental Rosario Izapa, Chiapas del INIFAP. Las dosis se aplicaron en un irradiador panorámico Gamma Beam 127 MDS Nordion, con fuente de almacenamiento de 50 g de cobalto 60 (Co60) en seco.
Prueba de germinación
Se utilizaron cajas Petri de 90 mm de diámetro provistas de algodón y papel filtro. La prueba de germinación se realizó bajo un diseño experimental completamente al azar, donde la unidad experimental fue la caja Petri con 50 semillas, con cuatro repeticiones por dosis de radiación. Cada repetición fue humectada con 25 ml de agua al inicio de la prueba y posteriormente, se suministraron riegos por aspersión de 2.0 ml cada dos días, durante los 17 d que duró la prueba. Las cajas Petri se colocaron a temperatura ambiente de 28 ± 2 °C. Se consideró semilla germinada aquella que alcanzó 0.5 cm de plúmula o de radícula. Las variables evaluadas fueron porcentaje germinación (PG), índice de velocidad de germinación (Vge), longitud de plúmula (LP) y longitud de radícula (LR). Para obtener LR y LP se dejaron crecer tres plántulas por caja Petri durante siete días después de germinadas. La Vge se calculó mediante la formula propuesta por Maguire (1962):
Dónde: Vge = velocidad de germinación, ni = número de semillas germinadas por día, t = día de la germinación.
Evaluación en invernadero
Para la prueba en invernadero se utilizaron las mismas dosis de irradiación que en la prueba de germinación, con excepción de la dosis de 4000 Gy. La cual se excluyó del análisis en invernadero por no presentar germinación. La prueba se realizó bajo un diseño experimental en bloques al azar con 10 repeticiones por tratamiento. Se sembraron 20 semillas por maceta para asegurar emergencia y establecimiento y dejar un individuo (planta) por maceta. La siembra se realizó en bolsas de polietileno negro de 26 cm de altura por 18 cm de diámetro, perforadas en la parte inferior. Las bolsas se llenaron con 23 cm de suelo franco-arenoso de origen aluvial. Los riegos se suministraron hasta punto de saturación cada dos o tres días, según el desecamiento del suelo. La prueba se realizó en una sola fase de crecimiento durante 16 semanas de mayo a agosto de 2016, periodo en que la plantas alcanzaron la madurez. Las variables evaluadas fueron rendimiento de forraje (RF), producción de semilla (PS), numero de tallos (NT), altura de planta (AP), diámetro de macollo (DM) e índice de concentración de clorofila (ICC). Para el PS se recolectó la totalidad de la semilla de cada planta y se colocó en bolsas de papel. Para el RF la biomasa se cortó a 5 cm del suelo y se colocó en bolsas de papel. Las muestras extraídas de forraje y semilla se secaron en estufa a 65 °C durante 72 h. Una vez secas las muestras se pesaron en balanza analítica marca Viper BC, Mettler. La AP se midió desde la base hasta el punto vegetal y el NT se contabilizó en la base de la planta. El ICC se midió en la parte media de cinco hojas seleccionadas al azar con medidor marca Opti-Sciences, modelo CCM-200.
Análisis estadístico
Los datos se analizaron mediante análisis de tendencia para cada variable y se seleccionó el modelo de regresión que presentó mayor nivel de significancia. Con la ecuación de regresión resultante se estimó la DL50 con base en el PG. La RG50 se determinó en las variables Vge, LP, LR, RF, PS, NT, AP, DM e ICC. Los datos se analizaron mediante comparación de medias con la prueba de Dunnett a un α=0.01, donde el factor de influencia fue la dosis de radiación. Los análisis se realizaron con el procedimiento GLM del paquete estadístico SAS 9.1.3 (2006). Con los resultados de la DL50 y RG50 obtenidos de las 10 variables evaluadas, por medio de regresión, se obtuvo la media ponderada. La DL50 se ponderó con 30%, mientras que el 70% restante se dividió equitativamente entre los valores obtenidos de RG50 de las nueve variables. Cabe señalar que la mayor ponderación se le dio a la germinación, ya que la muerte del individuo claramente es el máximo daño que puede producir la radiación.
RESULTADOS Y DISCUSIÓN
Al aumentar la dosis de irradiación, el comportamiento del PG mostró una tendencia lineal negativa (p < 0.0001). No obstante, las únicas dosis que presentaron diferencias (p < 0.01) con respecto al tratamiento testigo (T-0) fueron 1400, 2000 y 4000 Gy. En términos generales el PG tendió a disminuir conforme aumentó la dosis de irradiación (Figura 1a). Este resultado concuerda con diversos autores, quienes encontraron que la germinación de diferentes especies de gramíneas tiende a disminuir a medida que aumenta la dosis de radiación (Golubinova y Gecheff 2011, Rajarajan et al. 2016). Esto se debe a que la radiación ionizante puede deprimir o inhibir funciones vitales de las células, lo que provoca la muerte de algunas células e incluso la muerte del embrión. Este fenómeno tiende a aumentar con el incremento de la dosis de irradiación, lo que causa disminución de la germinación (Olasupo et al. 2016). La ecuación de regresión indica que la DL50 en pasto africano se presentó a 2076.1 Gy.
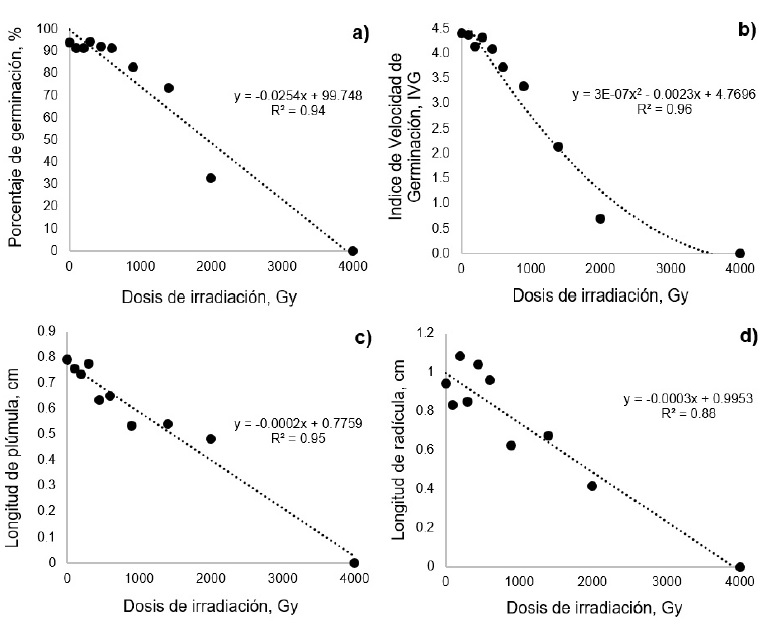
Figura 1: Efecto de diferentes dosis de irradiación con CO60 sobre el porcentaje de germinación (a), índice de velocidad de germinación (b), longitud de plúmula (c) y longitud de radícula (d) en pasto africano (Eragrostis lehmanniana).
Una de las gramíneas más estudias en cuestión de radiosensibilidad es el arroz (Oryza sativa). En esta especie Harding et al. (2012) al exponer a 13 variedades a radiación gamma encontraron que la DL50 varió entre 345 y 423 Gy. Otra especie gramínea donde se ha estudiado la radiosensibilidad es mijo perla (Pennisetum glaucum), donde la DL50 fue determinada en 669 Gy (Ousmane et al. 2013). Con respecto a gramíneas forrajeras la DL50 en pasto sudan (Sorghum sudanense) varia entre 307 y 342 Gy para diferentes variedades (Golubinova y Gecheff 2011). No obstante, los resultados anteriores son inferiores a la DL50 obtenida en este estudio para el pasto africano. Esto probablemente se debe a que la semilla de pasto guarda bajo contenido de humedad, ya que la radiación ionizante puede provocar ruptura de enlaces covalentes y descomposición de moléculas de agua. Lo cual forma radicales libres, que de manera indirecta dañan diferentes organelos de la célula e incluso las moléculas de ADN (Alegre 2001).
La Vge mostró una tendencia cuadrática (p < 0.0001). Los tratamientos 100, 200, 300 y 450 Gy no presentaron diferencias (p > 0.01) con el tratamiento testigo. De los 600 Gy en adelante, la semilla irradiada presentó menor Vge (p < 0.01) con respecto al testigo (Figura 1b). Este retraso en la germinación de las semillas irradiadas probablemente se debe a que la radiación inhibe funciones vitales para la célula como la síntesis de proteínas y la actividad enzimática, lo cual retrasa la división celular y por tanto su germinación (Chandrashekar 2014, Olasupo et al. 2016). De acuerdo a la ecuación, en esta variable la RG50 se encontró a 1357.8 Gy.
La LP presentó una tendencia lineal negativa (p < 0.0001). Sin embargo, la LP disminuyó de forma significativa (p < 0.01) a partir de los 450 Gy (Figura 1c). Según la ecuación de regresión, en esta variable, la RG50 se ubicó en 1900.5 Gy. De manera similar, la LR mostró una tendencia lineal (p < 0.0001). Para esta variable solamente los tratamientos con 900, 1400, 2000 y 4000 Gy tuvieron menor (p < 0.01) LR que el tratamiento testigo (Figura 1d). La ecuación indica que en esta variable la RG50 se presentó a 1748.3 Gy. Los resultados de las variables anteriores, concuerdan con lo reportado por Ambavane et al. (2014), quienes encontraron que en finger millet (Eleusine coracana) la LP y la LR se vieron afectadas por el incremento en la dosis de radiación. En este estudio se determinó la RG50 en 500 Gy. Este mismo efecto fue encontrado en arroz, donde la RG50 se registró en 250 y 450 Gy para ambas variables, respectivamente (Talebi y Talebi 2012). La disminución en la elongación de la plúmula y radícula puede deberse a que la radiación ionizante comúnmente provoca retraso en la actividad metabólica y la división celular, lo cual retrasa el crecimiento de las plántulas (Chandrashekar 2014, Olasupo et al. 2016).
El comportamiento del RF mostró tendencia cuadrática (p = 0.03), no obstante, las únicas dosis que presentaron diferencias (p < 0.01) con el tratamiento testigo fueron 1400 y 2000 Gy (Figura 2a). Cabe señalar que todas las plántulas que emergieron de la dosis de 2000 Gy murieron antes de 21 d de edad. Debido a esto, la variable RF en este tratamiento fue cero y lo mismo ocurrió para las otras variables evaluadas. De acuerdo con la ecuación de regresión, en esta variable la RG50 se presentó a 1528.3 Gy. De manera similar, la PS presentó tendencia cuadrática (p < 0.01) y las únicas dosis que presentaron diferencias (p < 0.01) con el tratamiento testigo fueron 1400 y 2000 Gy (Figura 2b). De acuerdo con la ecuación de regresión y la PS la RG50 se presentó a 1536.2 Gy.
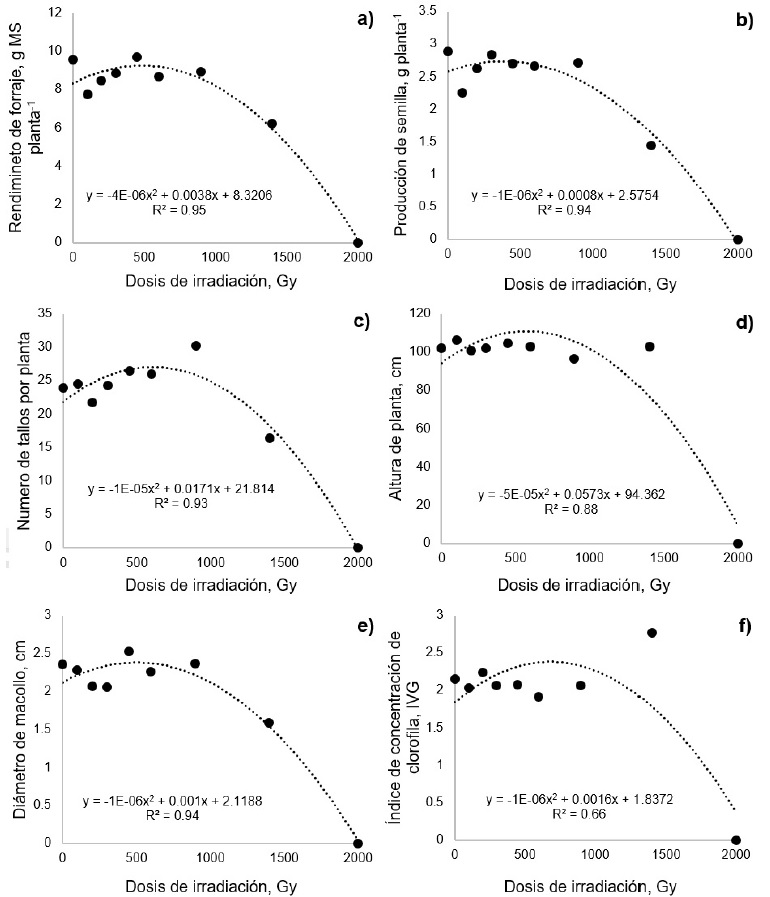
Figura 2: Efecto de diferentes dosis de irradiación con CO60 sobre el rendimiento de forraje (a), producción de semilla (b), número de tallos (c), altura de planta (d), diámetro de macollo (e) e índice de concentración de clorofila (f) en pasto africano (Eragrostis lehmanniana).
La variable NT mostró tendencia cuadrática (p = 0.0008). El único tratamiento diferente (p < 0.05) al testigo fue el de 2000 Gy (Figura 2c). En esta variable, la RG50 se tuvo en 1648.7 Gy. Del mismo modo la AP mostró tendencia cuadrática (p < 0.0001) y el único tratamiento diferente (p < 0.05) al testigo fue el de 2000 Gy (Figura 2d). De acuerdo con esta variable, la RG50 se presentó a 1666.9 Gy. El DM presentó tendencia cuadrática (p = 0.01), en esta variable los tratamientos que presentaron diferencias (p < 0.01) con respecto al testigo fueron 1400 y 2000 Gy (Figura 2e). De acuerdo a la ecuación de regresión y el DM, la RG50 se presentó a 1592.6 Gy. El ICC presentó comportamiento cuadrático (p = 0.0008). Al igual que en la variable anterior, los tratamiento con diferente ICC (p < 0.01) con respecto al testigo fueron 1400 y 2000 Gy; sin embargo, en la dosis de 1400 Gy, el ICC presentó incremento (p < 0.01, Figura 2f). De acuerdo con el ICC la RG50 se presentó a 1983.5 Gy. La media ponderada de la DL50 y la RG50 resultantes de las nueve variables fue de 1533 Gy.
Las variables RF, PS, NT, AP, DM disminuyeron al incrementar la dosis de radiación. Esto probablemente se debe a que la exposición a radiación gamma con frecuencia causa la destrucción de compuestos de importancia para el desarrollo vegetal, como auxinas y ácido ascórbico (Shah et al. 2008). Además, puede deberse a que la radiación puede causar daños en la estructura del ADN que en la mayor parte de las ocasiones afecta de forma negativa el desarrollo de las plantas (Kiong et al. 2008). Por otro lado, el ICC presentó un incremento con la dosis de 1400 Gy. Lo que concuerda con Jan et al. (2013), quienes encontraron que la radiación puede activar el sistema de los pigmentos fotosintéticos. Lo anterior, confirma que el comportamiento del pasto africano pudo haber sido influenciado por una radio estimulación, por la modificación ocurrida en el ICC. En este sentido, determinar la dosis óptima de radiación ayudará para realizar futuros trabajos en mutagénesis.
En la semilla irradiada a 2000 Gy hubo emergencia de plántulas, no obstante, ninguna logró sobrevivir más de dos semanas. Este fenómeno pudo deberse a que la radiación provocó un desorden metabólico en el pasto africano. Este mismo efecto se reportó por Marcu et al. (2013), quienes encontraron que las plántulas de maíz que emergieron de semilla irradiada con dosis mayores a 500 Gy, no fueron capaces de sobrevivir más de 10 d. La DL50 y las RG50 en este estudio variaron de 1357 a 2076 Gy. Para inducir variabilidad genética que posibilite la selección y mejoramiento genético de esta especie, se recomienda utilizar la media ponderada de ambos parámetros, la cual se obtuvo a 1533 Gy. Estos resultados permitirán la inducción de nuevo material genético de interés agronómico y ecológico.

