










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBotanical Sciences
versión On-line ISSN 2007-4476versión impresa ISSN 2007-4298
Bot. sci vol.90 no.3 México sep. 2012
Taxonomía y florística
Sierra de La Libertad: a major transition between two desert regions in Baja California, Mexico
Sierra de La Libertad: una importante transición entre dos regiones de desierto en Baja California, México
Elisabet V. Wehncke1,4, Jon Rebman1, Xavier López-Medellín1,2, and Exequiel Ezcurra3
1Biodiversity Research Center of the Californias. San Diego Natural History Museum, San Diego, CA 92101, USA.
2Center for US-Mexican Studies, University of California, San Diego, USA.
3Department of Botany and Plant Sciences, University of California Riverside, CA 92521, USA.
4Corresponding author: lizwehncke@gmail.com
Received: October 28, 2011
Acepted: March 22, 2012
Abstract
The Sierra de La Libertad, a group of prominent mesas and canyons in the Central Desert region of Baja California, is one of the most isolated and overlooked areas in the peninsula. For centuries no other explorations have entered this area with a scientific interest, and efforts to delimit its biogeographic boundaries are done mostly by extrapolating data from global information systems and from records in adjacent areas. This sierra constitutes one of the most problematic transition areas among specialists who try to delimit the ecoregions of the peninsula. We conducted two biological expeditions to the Sierra de La Libertad in order to document and describe the plant communities associated with the blue fan palm oases in this remote area. A total of 351 plant species were registered of which, 60 constitute new records for the northern state (Baja California) and two are new species to science. Additionally, in this transition area we found 45 regional and seven local endemics. Scientific expeditions to remote places are still significant endeavors for better understanding biogeography, ecology, taxonomy, and ultimately conservation of biodiversity, and these ventures are of particular importance in little known desert ecotone areas such as the Sierra de La Libertad.
Keywords: biodiversity, blue fan palm, Central Desert, desert oases, Vizcaíno Desert.
Resumen
La Sierra de La Libertad, un grupo de prominentes mesas y cañones en la región del Desierto Central de Baja California, es una de las áreas más aisladas y desconocidas en la península. Por siglos ninguna exploración ha entrado en este área con un interés científico y los esfuerzos por delimitar sus límites biogeográficos son realizados en su mayoría extrapolando datos de los sistemas de información geográfica global y de registros en áreas adyacentes. Esta Sierra constituye una de las áreas de transición más problemáticas entre los especialistas que intentan delimitar las ecoregiones de la península. Realizamos dos expediciones biológicas a la Sierra de La Libertad con el objetivo de documentar y describir las comunidades de plantas asociadas a los oasis de palma azul en esta área remota. Un total de 351 especies de plantas fueron registradas, de las cuales 60 constituyen nuevos registros para el estado norte (Baja California) y dos resultaron nuevas especies para la ciencia. Además en este área de transición encontramos 45 endemismos regionales y siete locales. Las expediciones científicas a sitios remotos siguen aún siendo esfuerzos significativos para un mejor entendimiento de la biogeografía, ecología, taxonomía, y finalmente la conservación de la biodiversidad, y estos emprendimientos son de particular importancia en áreas poco conocidas de ecotonos de desierto como la Sierra de La Libertad.
Palabras clave: biodiversidad, Desierto Central, desierto El Vizcaíno, oasis, palma azul.
More than three hundred years have passed since the Jesuit missionaries arrived on the Baja California peninsula and started the first biological and cartographic explorations (Kino et al., 1954; Kino, 1969; 1710; Rudkin, 1952). Since then, different studies have been performed by a large number of explorers and researchers, who provided the biological, geographical, and environmental information that gradually allowed the identification of particular biogeographic patterns in the peninsula. Subsequently, this information was delimited through detailed maps, and for more than a hundred years diverse biogeographic regionalizations for the peninsula have been proposed and developed (see, Garcillán et al., 2010; González-Abraham et al., 2010). Even though there seems to be a general agreement about the recognized regions, it is still difficult to determine and even more problematic to map the specific boundaries between the different communities or ecoregions without "on the ground" field work (Garcillán et al., 2010). After an exhaustive review of the published literature, González-Abraham et al. (2010) identified seven ecoregions showing disagreement between the specialists and proposed a new map of ecological regions and/or large vegetation types for the peninsula of Baja California.
With the combined effects of summer tropical rains, occasional hurricanes from the south, and cold weather and winter rain influences from the north, the mesas and canyons of the Sierra de La Libertad run deep into the Central Desert region and constitute one of the most interesting transition zones and one of the most controversial areas for vegetation experts. Probably due to its rough terrain and inaccessibility, the Sierra de La Libertad remains a very remote, intriguing, and overlooked region. Although many studies recognize (1) the climatic and geologic character of a central peninsular north-south transition (González-Abraham et al., 2010); (2) the existence of tropical and non-tropical components in the peninsular biota between 27 - 28° N latitude (Brandegee, 1892; Eisen, 1900; Nelson, 1921); (3) the difference between two ecoregions: the Central Desert in the north and the Vizcaíno Desert in the south based on climate and vegetation evidence (Aschmann, 1959); and (4) the presence of an ancient seaway associated with this transition zone (Gentry, 1978; Upton and Murphy, 1997; Aguirre-L. et al., 1999; Riddle et al., 2000; Zink, 2002), others do not distinguish it as an explicit transition zone (Wiggins, 1980; Peinado et al., 1993; Zipping and Vanderwier, 1994; Arriaga et al., 1997).
In 1751, the Jesuit missionary and geographer Fernando Consag registered for the first time the vegetation of the region and described the lifestyle and customs of its indigenous inhabitants. Later in 1889, T. S. Brandegee explored the Sierra de La Libertad with a particular botanical interest. Since then, this area seems to have become rather forgotten as a research target. In 2009, we performed two biological expeditions to the southern part of the Sierra de La Libertad and in particular along its major basin: El Paraíso canyon. Our particular objectives were to observe, survey, and document the local flora, as well as to describe the main plant communities of the remote desert oases, dominated by blue fan palms (Brahea armata). The presence of the palm oases supports the colonization and persistence of many other plant and animal species and provides an important structure to these ecosystems by trapping sediments and promoting multiple plant-animal interactions with the present-day fauna (Wehncke et al., 2009, 2010; Minnich et al. 2011).
Based on Risser's (1995) concept of ecotones, which refers to areas of steep gradients between more homogeneous vegetation associations, we should expect a high number of species extending their distributions to both sides of these margins as well as distinctive communities that take advantage of a variety of ecological niches in the transition zone. Commonly at broad spatial scales such as in biomes, biodiversity increases within the ecotonal zone because the ranges of many species correspond with the edges of adjacent biomes. Consequently, many species from two or more regions may be found in these transitional areas. Thus, near the periphery of their ranges the habitat of species is likely to be fragmented with many communities coexisting, therefore leading to a higher biological diversity (e.g., Whitcomb et al., 1981; Grover and Musick, 1990; Hansen and Urban, 1992; Gosz, 1993). It is in these blurry limits or ecotones where most of the energy flux exchange and the maximum interaction between different ecosystems and processes occurs (Risser, 1995, Franco-Vizcaíno et al., 2007; Wehncke et al., 2009, 2010). In this paper, we discuss the relevance of acknowledging this transition zone as an area of intense and varied climatic, geomorphologic, biological, and cultural mixing that plays a critical role not only for the distribution and exchange of numerous species, but also for the existence of endemic species.
Materials and methods
Study area. Two main regions are distinguished in the middle of the Baja California peninsula: the Central Desert towards the north, and the Vizcaíno Desert to the south (Aschmann, 1959). Based on the morphologic and anatomical differences of the genus Agave and its relationship with geological changes in the peninsula, Gentry (1978) mentioned the presence of an ancient sea intrusion that divided this area. This was previously suspected by the Jesuit missionary Miguel del Barco in 1780, when he interpreted the existence of marine shells in the middle of the peninsula and far from the coast as the result of the elevation of a part of the surface that had been submerged (González-Abraham et al., 2010). Once molecular studies were available investigations on the genealogical relationships among mtDNA lineages made great advances to elucidate the historical biogeography of Baja California (Lindell et al., 2006). With an extension of nearly 2,500 km2, the Sierra de La Libertad (SLL from now onwards) is almost entirely located above 28º N (27º 49' - 28º 50' N; 113º 22'-112º 43' W). Phytogeographically, it is located within Shreve's Vizcaíno Region, also known as the Central Desert of Baja California. El Paraíso Canyon (28º 30' N; 113º 4' W), the largest basin of SLL, is located in the southern part of the sierra and flows into the Pacific Ocean (Figure 1). This canyon is fed by many diverse tributaries that run from the highest elevations of the sierra. At their margins we can find true oases dominated by the blue fan palm (Brahea armata), which provide particular microclimatic conditions and habitats for many species. The area corresponds to the Central Desert Ecoregion according to González-Abraham et al. (2010).
The paleobotanical evidence shows that during the Cretaceous, a paratropical flora with Nearctic and Neotropical features, including Coryphoid palms (Arecaceae, Coryphoideae) was widely distributed in the northern hemisphere to approximately 65º N (Wehncke et al., 2009, 2010). Towards the end of the Miocene (12 million years ago), as climate became dryer and warmer, this vegetation became restricted to higher elevations or canyons that still maintained mesic conditions, creating isolated refuges in the middle of the sierra. By reducing and expanding their margins these populations followed the pace imposed by the cold and humid fluctuations of the Pleistocene (1.8 million-10,000 years ago). At present, these ecosystems constitute critical aquifer catchments and biotic communities that contrast with the surrounding, more arid, environments, and provide diverse ecological services that are fundamental for the region (Wehncke et al., 2012).
El Paraíso Canyon. After the first mission was settled in Loreto in 1697, subsequent explorations performed by Fernando Consag and Wenceslao Linck, among others, left significant information that constitute the first reports of the flora, fauna, ethnography, and biogeography of the northern desert region (del Barco, 1780). Since the beginning Kañayiakamán, as El Paraíso was called by the natives, enticed the interest of Consag who named it 'El Paraíso de los Californios' because of the extraordinary descriptions by the natives of plentiful water and abundant resources in the area (Lazcano-Sahagún, 2000). In 1751, Consag entered the canyon passing through a very rugged landscape dominated by large palm groves covering hills and hanging from cliffs (del Barco, 1780; Lazcano-Sahagún, 2000). This constitutes the first reports that mentions blue fan palms (Brahea armata), together with a tree they called milapa, which is the boojum or cirio (Fouquieria columnaris), the presence of an ivy which they called 'yedra maligna' (Toxicodendron sp.), that produces strong allergic reactions when touched, and as an extension of the Vizcaíno Desert into this area, he mentioned the presence of yucca (Yucca valida; del Barco, 1780; Lazcano-Sahagún, 2000, 2001). Even though Consag never started a mission in this place, a ranch called El Paraíso was founded to provide a resting site for caravans moving between the missions. Traces of ancient roads, ranches, refuges, and even rock walled corrals can still be found abandoned along these canyons. Much later, in 1889, T. S. Brandegee led a botanical expedition to this area, but unfortunately the information regarding the specific localities of specimen collections was scarce, sometimes misleading, or even lost (Moran, 1952). Some time later, Nelson (1921) and Ewan (1942) reconstructed and compiled the available information that could be extracted from Brandegee's specimen labels and letters deposited at the University of California Berkeley Herbarium (UC).
Based on this partial and incomplete information we explored these mountain ranges by air in 2006. From the air, we identified the main El Paraíso Canyon, its tributaries, and many remote blue fan palm oases, but we could not see any roads to reach them. In April and October of 2009, we entered the area by mule with the main objective of surveying and documenting the local ecosystems and their vegetation (Figure 1). Voucher specimens of the flora collected in both expeditions are deposited in the herbaria of the San Diego Natural History (SD) and the Universidad Autónoma de Baja California (BCMEX), in Ensenada, Mexico.
Results
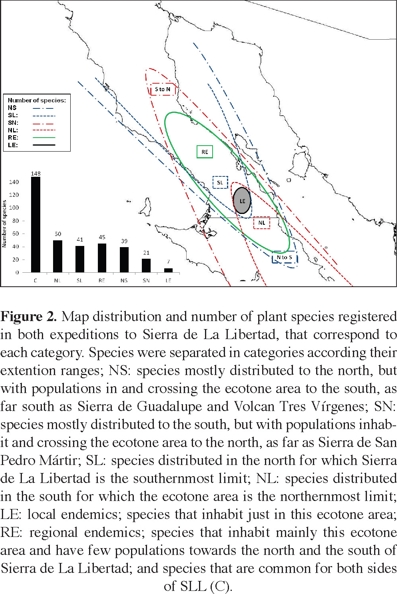
Vegetation description and major plant collections. Our expeditions to SLL contributed with a total record of 351 plant species (Appendix), of which 60 represent records of species that were not previously known to be in the northern Baja California region (state of Baja California). A total of 288 species were collected and other 63 were observed in the field. At a coarse scale, 71 species (20% of the total number of species registered during the two expeditions) are distributed mostly in the southern state (Baja California Sur), 80 (23%) species in the northern state (Baja California), and 148 (42%) species are common to both sides of SLL. On the other hand, 193 of the 351 species registered also inhabit the southern Sierra de San Francisco (SSF; Rebman unpublished flora) and 131 species are also found in the northern Sierra de La Asamblea (SA; Bullock et al., 2008). Altogether, these three sierras share 82 plant species (23%). Nonetheless, 109 species (31%) are present neither in the Sierra de San Francisco nor in the Sierra de La Asamblea; while 39% constitute common species found in both sides of SLL, 21% is formed by species expanding from the south, 30% by species expanding from the north, and 9% is composed by local and regional endemics that were not found in the adjacent sierras.
At a smaller scale of patches or communities, numerous northern species widen their distribution to the south and vice versa. In the first category (NS; Appendix 1, Figure 2), we grouped those species that are mostly distributed to the north. A few species in this group are also found further south, including those endemic to Baja California (BC) that are rarely distributed in Baja California Sur (BCS), but it also includes those species ranging as far south as the Sierra de San Francisco, Volcán Tres Vírgenes, and Sierra de Guadalupe. Thus, we found 39 (11%) northern species that use the SLL transition and extend their distributions to the south, and 21 (6%) southern species that have few populations distributed further north, ranging as far as Sierra de La Asamblea, and eastern Sierra de San Pedro Mártir (SN; Appendix, Figure 2). Furthermore, we identified 41 (12%) plant species commonly distributed in the north for which SLL represents the southernmost limit (SL), and 50 (14%) southern species for which SLL represents the northernmost limit of their distribution (NL; Appendix, Figure 2). Particularly, the southern part of SLL constitutes an area of local endemics and we considered seven main species (2%) into this category: Eysenhardtia peninsularis (Fabaceae), a new species of Stephanomeria (Asteraceae), a new species of cholla cactus, Cylindropuntia sp. nov. (Cactaceae); Holographis virgata ssp. virgata (Acanthaceae), Rhus kearneyi ssp. borjaensis (Anacardiaceae), Agave avellanidens (Agavaceae), and Hedeoma tenuiflora (Lamiaceae). Additionally, the following collected species were also considered an interesting locally endemic group: Astragalus orcuttianum, A. prorifer, Brahea armata, Condalia brandegeei, Cryptantha fastigiata, Chorizanthe rosulenta, Dudleya gatesii, Eriogonum orcuttianum, E. pilosum, Penstemon eximius, Salvia peninsularis, Tragia moranii, Vallesia laciniata, and Xanthisma spinulosum var. gooddingii (Appendix). At a coarser scale of analysis, SLL constitutes an area of regional endemism: 45 species (13%) were identified into this group (RE; Appendix, Figure 2). These species inhabit mainly this ecotonal area, but a few populations can be found towards the north and south of SLL.
A major finding of these expeditions was the discovery and collection of two plant species new to science. As already mentioned, Cylindropuntia sp. nov. (Cactaceae) is a big succulent shrub that is widely distributed in El Paraíso Canyon at nearly 750 m in elevation (Figure 3A, B), and it seems to be a morphological type between C. cholla and C. alcahes. The other new taxon is Stephanomeria sp. nov. (Asteraceae), which was found on a single population along a tributary of El Paraíso Canyon at 870 m. Even though seeds are needed to properly describe the new species, Dr. Les Gottlieb, a taxonomic expert in the genus Stephanomeria, corroborated that it is a new species. This new Stephanomeria is a very rare perennial herb, up to 1 meter tall, with blue green leaves, lavender petals and purple stigmata (Figure 3C).
Three other extremely rare species constituted significant botanical discoveries in the SLL. Sageretia wrightii (Rhamnaceae) and Eriogonum abertianum (Polygonaceae) were each only known in the peninsula from one single historical specimen. Sageretia wrightii was known only from a specimen collected by A. Carter in 1963 in the Sierra de La Giganta, BCS, and during our expeditions we found two different populations in SLL. Similarly, E. abertianum was only known from a specimen collected by T. S. Brandegee in 1889 in the vicinity of San Regis, which is near El Paraíso Canyon. In 2009 we found and collected specimens of this species in two different sites: one at 970 m on volcanic substrate, and the other at 1,200 m growing on basalt substrate on rocky slopes and mesas. The third rare species, Hedeoma tenuiflorum (Lamiaceae), was collected and described by T. S. Brandegee based on solely one specimen from 1889. Later in 1964 and 1966, Reid Moran found this species in two different places, one near Sierra San Borja, at the northern limit of SLL, and the other at SLL's southern limit. This endemic species was recorded and collected in one population during our first expedition at an altitude of 1,260 m, which constitutes the second occasion in which specimens have been collected with flowers. It seems to be endemic only to SLL, where it is also quite rare.
An interesting botanical note of our expeditions to SLL was the rediscovery of Eysenhardtia peninsularis (Fabaceae). First collected in the vicinity of the El Paraíso Canyon in 1889 by T. S. Brandegee, he later named this rare and endemic legume as E. peninsularis in 1911 based on his one specimen. For more than a century this species was never collected again and it was thought to be possibly extinct. In May 1992, J. Rebman, H. Cota, and L. Slauson rediscovered this rare species in the Sierra de San Francisco, BCS. Only a few shrubs severely impacted by cattle and goat browsing were observed on that occasion and one specimen was sent to R. Barneby at the New York Botanical Garden who confirmed the identification. Other taxonomic descriptions of E. peninsularis include: F.W. Pennell (1919); F. Shreve (1951); and I. Wiggins (1963); and one monograph of the genus Eysenhardtia by J. M. Lang (1972). Wiggins (1980) briefly describes E. peninsularis illustrating and comparing it with E. polystachya, a species from Durango and Tamaulipas to Oaxaca. All these descriptions were based on the only specimen collected by Brandegee in 1889. During our 2009 expeditions, we discovered four populations of E. peninsularis along a range of approximately 9.3 km. Two were small populations distributed along the canyon drainages between 720–870 m. The other two populations were found over the mesas at nearly 1,200 m, and constitute the largest populations known of this rare endemic species. These populations were found on a whitish volcanic tuff substrate with other species like Bernardia myricifolia, Calliandra californica, Croton magdalanae, Cylindropuntia alcahes, Dalea bicolor var. orcuttiana, Dodonaea viscosa, Fouquieria splendens, Krameria erecta, Prosopis articulata, Senegalia greggii, Solanum hindsianum, and in many cases with Brahea armata as well. Even though E. peninsularis had been rediscovered in 1992, large populations such as the ones found during our expeditions had not been documented. The high grazing pressure exerted by cattle and goats could be an important factor explaining its rarity and scarcity in the central peninsula. We suggest that efforts should be directed to manage cattle and goat activities in some select areas in order to preserve this unique and locally endemic shrub species.
During the 2009 expeditions, thirteen other species documented were particularly significant collections: (1) Muhlenbergia minutissima (Poaceae), was previously only known from a small number of voucher specimens mainly from Sierra de San Pedro Mártir; (2) Nolina palmeri var. palmeri (Ruscaceae) shows here its southernmost specimens; (3) Oenothera brandegeei (Onagraceae) is rare on the peninsula and only known from only a few voucher specimens; (4) Panicum capillare (Poaceae) is represented here in its southernmost distribution; (5) Mimosa aculeaticarpa var. biuncifera (Fabaceae), also very rare in the peninsula, shows here what is probably the southernmost specimen ever found; (6) M. appressa (Poaceae) is rare, not regularly documented in the peninsula, and usually distributed in small "sky islands"; (7) Ipomoea cristulata (Convolvulaceae) and (8) Bidens leptocephala (Asteraceae) are both mainly from BCS with only a pair of collections known from BC; (9) Pseudognaphalium canescens (Asteraceae) also shows here its southernmost specimen; (10) Abutilon incanum (Malvaceae) grows here with striking white flowers while the usual collections have yellow and/or orange flowers; (11) Chenopodium fremontii (Chenopodiaceae) has here its southernmost specimen; (12) M. emersleyi (Poaceae) is a rarely-collected species from the SLL; and, finally, (13) Aesculus parryi (Sapindaceae) shows here its southernmost specimen, collected far from the Mediterranean climate where it usually prospers.
Another interesting natural history observation of these expeditions was the presence of the mistletoe Phoradendron brachystachyum (synonym = P. diguetianum) parasitizing several host plants such as, Celtis pallida (Cannabaceae), Condalia brandegeei (Rhamnaceae), C. globosa var. pubescens (Rhamnaceae), Fouquieria splendens ssp. splendens (Fouquieriaceae), Prunus ilicifolia, P. fremontii (Rosaceae), Schoepfia californica (Schoepfiaceae), Vachellia farnesiana (Fabaceae), and Vallesia laciniata (Apocynaceae). The interaction of Phoradendron brachsytachyum with F. splendens has, to our knowledge, never been registered before on the peninsula. Other two species of mistletoe have been found in SLL, one is the widely distributed P. californicum that was parasitizing different shrubs and trees from the family Fabaceae; while P. villosum was found in trees and shrubs of the genus Quercus (Fagaceae).
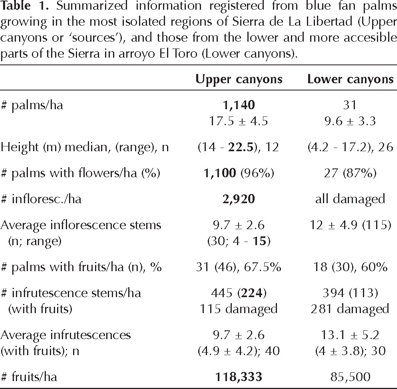
Along the El Paraíso Canyon, their tributaries and on the mesas, we found what probably are the most remote and dense blue fan palm populations of the peninsula (Figure 4). In Table 1, we compared some biological information of these populations growing in what we called the "upper parts", or "the sources", with those distributed in the more accesible canyons, or what we called the "lower parts" of the Sierra. We registered more than 300 plant species associated with Brahea armata in the upper parts and along remote canyons, while in the lower parts we found more than 100 plant species. Three communities were of particular interest around blue fan palms: (1) the group of orchids, ferns, oaks, and climbing vines along canyons of El Paraíso in the upper parts or sources, and their tributaries (Figure 5A), (2) the diverse and rare species of grasses at higher altitudes on the mesas (Figure 5D), and (3) the association of ocotillo, Fouqueria splendens, with blue fan palms on a whitish-volcanic tuff substrate up on the mesas (Figure 5E).
Communities living along the canyons. Before reaching El Paraíso Canyon we surveyed the northern part of arroyo El Toro (635 m), which has many patches of blue fan palms imbedded on an arid matrix composed of Aesculus parryi, Agave shawii ssp. goldmaniana, Cylindropuntia alcahes, C. cholla, Fouquieria columnaris, and Pachycereus pringlei. Along the canyon and associated with Brahea armata we recorded Ambrosia ambrosioides, Baccharis salicifolia, B. sarothroides, Chloracantha spinosa, Dodonaea viscosa, Proso-pis articulata, and Vachellia farnesiana. A rare collection was the climbing Macroptilium atropurpureum (Fabaceae) with dark purple flowers, which constituted a new record for BC. By walking down several mesas through a volcanic substrate covered by populations of C. cholla, Encelia farinosa, F. columnaris, and P. pringlei we reached the El Paraíso main stream where we found the abandoned San Bartolo ranch at an altitude of 500 m. Here the substrate becomes granitic and the species guild associated with B. armata was composed of particular riparian populations of A. ambrosioides, B. salicifolia, C. spinosa, D. viscosa, P. articulata, and V. farnesiana. Around the ranch we also found date palms, Phoenix dactylifera, introduced by the former inhabitants of the area and the perennial grass Phragmites australis reaching 2 m high. Continuing along the El Paraíso Canyon and after crossing an area with ancient cave paintings we reached the Santa Marta area at 750 m, where once a ranch was settled. Date palms, P. dactylifera of several age-size categories congregate here in high densities. We continued to El Paraíso Viejo, another abandoned ranch where we found 12 new plant records for the northern BC state including the new species of Cylindropuntia. This new and undescribed species reproduces very well vegetatively and interestingly, the fleshy fruits that fall to the ground and were not eaten by animals can develop roots and establish as new clonal individuals. Further along El Paraíso Canyon, blue fan palm concentrations increased as well as those of wild figs, Ficus palmeri. In April, several plant species were flowering including blue fan palms and the cochal cactus Myrtillocactus cochal, the scent of which was very strong along the way. Along a tributary, arroyo El Descanso (870 m), blue fan palms were particularly tall and dense, and the community around them included A. ambrosioides, B. salicifolia, C. spinosa, D. viscosa, P. articulata, Toxicodendron diversilobum, and oaks, Quercus oblongifolia (Figure 6). This spot has an important water hole; the canyon is very humid and deep reducing the number of light hours in it, probably determining the height of these palms (Figure 6). On the floor, we found several blue fan palm seeds oviposited by the larvae of the beetle Coryobruchus veseyi Horn (Bruchidae; see Wehncke et al., 2009).
Another wide tributary of the El Paraíso Canyon is El Olvido, into which four canyons with abundant blue fan palm populations merge (Figure 4A). We selected the most accessible of these canyons (La Misericordia) to survey the vegetation and estimated a density of 114 reproductive blue fan palm trees per /0.1 ha, of which 96% were in flower in April (Figure 5A). Contrary to what was expected, instead of finding 'yedra maligna', Toxicodendron radicans (Anacardiaceae), that is normally found in the Sierra de San Francisco and BCS, most blue fan palm trees were covered by the vine 'roble venenoso', T. diversilobum, which was out of its normal distribution range on the Pacific slopes of the Sierra de San Pedro Mártir and Sierra de Juárez (Figure 5A). Around blue fan palms the community was composed of Ambrosia ambrosioides, Baccharis salicifolia, Dodonaea viscosa, Prosopis articulata, T. diversilobum, the ferns Adiantum capillus-veneris, Pellaea wrightiana, Pentagramma triangularis, and Thelypteris puberula, and the orchid Epipactis gigantea (Figure 5B, C).
Communities living on the mesas. By using the old missionary road we reached a locality called Las Cuevitas in the higher parts of the mesas at 1,040 m. Bordering the mesas and cliffs and around blue fan palm individuals the community was composed of Agave cerulata, Cylindropuntia alcahes, Dodonaea viscosa, Echinocereus engelmannii, Ficus palmeri, Fouquieria splendens, Gambelia juncea, Malosma laurina, Prunus ilicifolia, Quercus turbinella, and Vauquelinia californica. We also found a diverse community of grasslands in a very good state of conservation with native perennial grasses such as Melica frutescens. This community is distributed over a remarkable landscape of volcanic rocks forming a very fractured substrate covered with high densities of blue fan palms (Figure 4B). Compared with the populations growing along the El Paraíso tributaries, these palms were lower in size (from 1.5 to 6 m tall, average 3.5 ± 1.3 m, n = 20). In April, these populations were flowering (average 6.9 ± 2.4 inflorescence per palm, n = 20).
In the higher parts of the mesas, between 1,050 and 1,100 m, Brahea armata was surrounded by Bernardia myrcifolia, Cordia curassavica, Croton magdalenae, Dodonaea viscosa, Prosopis articulata, and Vachellia greggii. Between them we also found two new records for the state of BC, a Rubiaceae, Diodia teres, and a Verbenaceae, Lantana hispida. Still on the mesas and along a deep canyon, palms produced inflorescence stems, but only 67.5% of them had green immature fruits later in October (estimated from 40 palms in 0.87 ha). Palms produced a total of 387 inflorescence stems in this patch (average: 9.7 ± 2.6; range 3–18), from which only 195 had immature fruits (average: 5 ± 4.2). Calculating the percentage of fruits produced per palm tree and considering that each inflorescence may approximately produce 1,000 fruits, we estimated that 40 palm trees produced 118,333 fruits/ha. This total represents only 27% of the fruit production that could be achieved if all the inflorescence stems that developed would have produced fruits. Furthermore, if the inflorescences would have developed 100% of fruits, the number of fruits per ha would approximately have reached 445,000. In reality, however, numerous (26%), of the infrutescence stems were damaged at the date of sampling. We registered a loss of 32.5% of the fruits due to damage in this patch, and numerous aborted fruits were found lying on the soil. The same kind of damage was also documented in the populations of the lower parts of the Sierra (E.Wehncke, unpublished data), and is related with the larva of an endemic moth, Litoprosopus bajaensis. Deep in Las Cuevitas canyon, the landscape presented many red and black volcanic foldings (Figure 7), and blue fan palms increase in height; the tallest size we measured was of 23 meters and represents the highest record for a blue fan palm tree (see estimation of Bullock and Heath, 2006). Here we found Baccharis sarathroides, Dodonaea viscosa, Ficus palmeri, Quercus turbinella, and Vachellia greggii, plus five new records for BC were found; three rare species of perennial grasses (Poaceae), one Pontedericaceae, and one Fabaceae (Appendix, Figure 7).
On the way to El Rodeo, we bordered the northern slopes of the Siete Cerritos Mountains, and found Brahea armata with Celtis pallida, Dodonaea viscosa, Cylindropuntia cholla, and C. alcahes growing on a basaltic substrate. Here, we found another new record for BC, the Amaranthaceae, Amaranthus torreyi. Once at El Rodeo at 1,220 m, and around a canyon with abundant water, we found blue fan palms of up to half a meter tall, most of which were browsed by cattle (Figure 8). Here, on a substrate formed by a whitish volcanic tuff, blue fan palms were associated with C. pallida, Condalia brandegeei, C. cholla, C. alcahes D. viscosa, and Prosopis articulata (Figure 8). At this site, we also documented 15 species that represent new records for BC (Appendix).
Discussion
Far from trying to exactly delimit new boundaries between the different communities of organisms inhabiting the central region of the Baja California peninsula, our intention is to provide evidence that shows the ecotone character of this area and with this to stress the special attention it deserves. Above 28° N, the SLL forms a true transition between the Central Desert and the Vizcaíno Desert, where the overlapping distribution ranges of many species create particular and unique communities. This transition and the climatic contrast between the north and the south, the topographical change, and the change in the flora have been noted by many explorers and researchers starting with Jesuit missionaries like Consag in 1773, Longinos 1792, and subsequently Eisen 1893, Nelson 1921, Shreve 1951, Wiggins 1980, etc. (see Garcillán et al., 2010, for a thorough historical review), and more recently by Turner et al. 1995, González-Abraham et al., 2010. However, it was Brandegee who between 1889 and 1902 provided the first botanical data and suggested the marked change in the flora that occurs at about latitude 28° N. Later on, Shreve's (1951) observations of the Sonoran Desert suggested that life forms are distributed according to their strategies in order to survive the aridity of the desert and to conserve humidity. The combined latitudinal effects of the northern and southern climates (summer tropical rains with occasional hurricanes, and cold temperatures combined with winter rains), as well as the altitudinal effects that determine the western and eastern oceanic influences in the local weather of SLL (Reyes-Coca et al., 1990; Cavazos-Pérez, 2008) create a melting pot area of ca. 2,500 km2 with numerous ecological niches where species with more northern and southern affinities coexist. This region has consecutive years with little rainfall interrupted by the sporadic passing of a hurricane and two to three winter storms. Seventy-nine percent of the annual rainfall almost exclusively depends upon extreme events (Cavazos-Pérez, 2008). Growth tolerance of Brahea armata can explain its more widespread distribution and higher elevation limits in the region when compared to the other palm species found on the peninsula (Minnich et al., 2011), and match with the high variability in the amount of rainfall over the region that is different from other areas of the Baja California Peninsula (Cavazos-Pérez, 2008). Records of new endemics and extended distributions of species documented in the brief surveys of the 2009 expeditions evidence the transition role of these canyons and mesas as well as the potential for future research in this area.
The two new species of Cactaceae and Asteraceae found along the humid stream sides of El Paraíso Canyon illustrate the scarce scientific reconnaissance that this particular sierra has had. The collection of locally endemic plant species, as well as the rare and uncommon plants that comprise 68% of the total species recorded in the area; the bulk of southern species that extend their distribution ranges to the north; and the numerous northern species that broaden their distribution areas further south from their normal distributions highlights the transitional nature of the Sierra de La Libertad. Besides filling information gaps in herbaria and other specimen collections, these expeditions identified particular and unknown plant communities associated with blue fan palm populations, the dominant vegetation of desert oases at these latitudes. Thus, orchids, ferns, and climbing vines growing with palms close to water holes form a different type of tropical forest; small-sized palms growing within a matrix of diverse species of grasses that sway in the breeze of the high mesas; and the ocotillos interspersed with the palms covering the rocky landscape of whithish volcanic tuffs are some of the most noteworthy and unique communities registered in the area.
The transition between the two Brahea species of palms on the peninsula (B. armata and B. brandegeei) is still unclear because of taxonomic uncertainties, but seems to coincide with the 28° N parallel (Felger and Joyal, 1999; Felger et al., 2001). Based on Google-Earth images Minnich et al. (2011), indicated that blue fan palms concentrate in the southern part of SLL. They proposed that palms occur in Miocene basalt tablelands overlying granitic basement, on volcanic caprock, and exposed granitic rock. The surveys we performed in-site indicated that blue fan palm populations on the mesas were distributed over a peculiar substrate related to a layer of peralkaline ignimbrites (L. Delgado pers. comm.). These ignimbrite units, correspond to processes of simple cooling, and are composed by different lithographic layers including a black layer of volcanic glass over which there is a whiter level formed by fractures that evidence the presence of water in these valleys (Vidal-Solano et al., 2007). A peculiar volcanic episode represented by ignimbrites (pyroclastic peralkaline fluxes) has been recognized among the Tertiary volcanic sequences of central Sonora and part of Baja California up to SLL, as a drastic change in the Tertiary magmatism during the Middle Miocene (12 My; Vidal-Solano et al., 2007). Blue fan palm roots can reach the water trapped in these lithographic layers, a factor which may explain their abundance on the mesas.
Following the intermittent flow of occasional rains or bearing the intense flooding after sporadic hurricanes, the plant communities dominated by blue fan plams provide a somewhat predictable pulse of resources and create diverse ecological niches for myriad other organisms throughout this mountain range. Massive production of nectar, flowers, fruits, seeds, and leaves are the focus of a highly diverse insect community that live in the area and also use canyons as corridors to connect isolated populations. New species of insects have been found in the inflorescences, infrutescences, and leaves of blue fan palms, and astonishing interactions are starting to be noted as the area is getting more explored (E. Wehncke, unpublished data).
Numerous explorations and descriptions of the Baja California fauna, flora, geology, and local inhabitants have occurred since the arrival of the Jesuit missionaries to the peninsula. By the end of the XIX century, successive scientific explorations noted the contrasting change in climate and flora around the 28° N latitude (Brandegee, 1889, 1891, 1892; Nelson, 1921; Ewan, 1942; Wiggins, 1963). Throughout the years and with the development of new technology, several distribution ranges of organisms were incorporated with climatic and geologic data to produce the first biogeographic regionalizations for the peninsula (see Garcillán et al., 2010). Nonetheless, we feel that apart from trying to delimit the exact boundaries of the transitions between ecoregions, a huge and significant gap will be filled when the SLL is better understood biologically, and more importantly, recognized as a true ecotone between the northern and the southern desert regions of the Baja California Peninsula. Today, in an era of incessant scientific and technological advances, when communications go beyond distances and roads, the value of scientific expeditions to access and explore remote places is still huge.
Acknowledgments
These expeditions were financed by a fellowship of the Packard Foundation to E.Ezcurra, and by the JiJi Foundation, CA. We want to thank the San Diego Natural History Museum and the Valle de los Cirios Protected Area for support and logistics. We are grateful to Víctor Sánchez Sotomayor, director of the APFF, Valle de los Cirios, Baja California, Michael Wall, Sandy Lanham, Carolina Espinoza, and Gonzalo Rodríguez for all their help. Photographs were taken by Miguel Ángel de la Cueva from the International League of Conservation Photographers, Planeta Peninsula, AC., J. Rebman, and by E.Wehncke. We are grateful to Hexiquio and Carlota Mendoza and Francisca and Ignacio Flores for showing us the way to enter this area and welcoming us into their lands and houses, and Matilde, Andrea, Pancho, Nacho and Tomás Murillo who were our guides during the expeditions. We express our gratitud to Dr. R. Felger for his helpful review and pertinent comments on this manuscript.
Literature cited
Aguirre-L. G., Morafka D.J. and Murphy R.W. 1999. The peninsular archipelago of Baja California: a thousand kilometers of tree lizard genetics. Herpetologica 55:369-381. [ Links ]
Arriaga L., Aguilar C., Espinosa D., y Jiménez R. Eds. 1997. Regionalización Ecológica y Biogeográfica de México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México, D.F. [ Links ]
Aschmann H. 1959. The Central Desert of Baja California: Demography and Ecology. University of California Press, Berkeley. [ Links ]
Brandegee T.S. 1889. A collection of plants of Baja California. Proceedings of the California Academy of Sciences 2nd ser. 2:117-232. [ Links ]
Brandegee T.S. 1891. Flora of the Cape Region of Baja California. Proceedings of the California Academy of Sciences 2nd ser. 3:108-182. [ Links ]
Brandegee T.S. 1892. The distribution of the flora of the Cape Region of Baja California. Zoe 3:223-231. [ Links ]
Bullock S.H. and Heath D. 2006. Growth rates and age of native palms in the Baja California desert. Journal of Arid Environments 67:39-402. [ Links ]
Bullock S.H., Salazar-Ceseña J.M., Rebman J.P. and Riemann H. 2008. Flora and vegetation of an isolated mountain range in the desert of Baja California. The Southwestern Naturalist 53:61-73. [ Links ]
Cavazos T. 2008. Clima. In: Danemann G.D. and Ezcurra E. Eds. Bahía de los Angeles: Recursos Naturales y Comunidad. Línea Base 2007, pp. 67-90. Secretaría de Medio Ambiente y Recursos Naturales. México, D.F. [ Links ]
del Barco M. ca. 1780. Correcciones y adiciones a la historia o noticia de la California en su primera edición de Madrid, año de 1757. En León-Portilla M. Re-Ed.(1988) Historia Natural y Crónica de la Antigua California: Adiciones y Correcciones a la Noticia de Miguel Venegas, pp. 379. Universidad Nacional Autónoma de México, México, D.F. [ Links ]
Eisen G. 1900. Explorations in central Baja California. Journal of the American Geographical Society of New York 32:397-429. [ Links ]
Ewan J. 1942. Bibliographical miscellany. IV. A bibliographical guide to the Brandegee botanical collections. American Midland Naturalist 27:772-789. [ Links ]
Felger R.S. and Joyal E. 1999. The palms (Arecaceae) of Sonora, Mexico. Aliso 18:1-18. [ Links ]
Felger R.S., Johnson M.B. and Wilson M.F. 2001. The Trees of Sonora, Mexico. Oxford University Press, New York. [ Links ]
Franco-Vizcaíno E., López-Beltrán A.C. and Salazar-Ceseña M. 2007. Water relations and community composition in three blue fan palm oases across the Californian-Sonoran biome transition. The Southwestern Naturalist 52:191-200. [ Links ]
Garcillán P.P., González C.E. and Ezcurra E. 2010. The cartographers of life: two centuries of mapping the natural history of Baja California. Journal of the Southwest 52:1-40. [ Links ]
Gentry H.S. 1978. The Agaves of Baja California. California Academy of Sciences, San Francisco. [ Links ]
González-Abraham C.E., Garcillán P.P. and Ezcurra, E. 2010. Ecorregiones de la península de Baja California: una síntesis. Boletín de la Sociedad Botánica de México 87:69-82. [ Links ]
Gosz J.R. 1993. Ecotone hierarchies. Ecological Applications 3:369-376. [ Links ]
Grover H.D. and Musick H.B. 1990. Shrubland encroachment in southern New Mexico, USA: An analysis of desertification processes in the American Southwest. Climate Change 17:305-330. [ Links ]
Hansen A.J. and Urban D.L. 1992. Avian response to landscape pattern: The role of species' life histories. Landscape Ecology 7:163-180. [ Links ]
Kino E.F. 1710. Favores celestials de Jesús y de María SSa y del gloriosíssimo apóstol de las indias Francisco Xavier. En Gobierno del Estado de Sonora, Hermosillo Ed. (1985) Crónica de la Pimería Alta. [ Links ]
Kino E.F. 1969. First from the Gulf to the Pacific: The diary of the Kino-Atondo peninsular expedition, December 14, 1684 – January 13, 1685. Dawson's Book Shop, Los Angeles. [ Links ]
Kino E.F. and Jesuits 1954. Kino reports to headquarters; Correspondence of Eusebio F. Kino, S.J., from New Spain with Rome. Institutum Historicum Societatis Jesu, Rome. [ Links ]
Lang J.M. 1972. Eysenhardtia (Leguminosae): Taxonomic revision and relationships. Doctoral dissertation, Iowa State University, Ames, 4692 pp. [ Links ]
Lazcano-Sahagún C. 2000. La Primera Entrada. Descubrimiento del Interior de la Antigua California. Colección de documentos sobre la historia y la geografía del municipio de Ensenada. No. 3. Fundación Barca/Museo de Historia de Ensenada, Ensenada. [ Links ]
Lazcano-Sahagún C. 2001. Fernando Consag. Textos y Testimonios. Colección de documentos sobre la historia y la geografía del municipio de Ensenada. No. 4. Fundación Barca/Museo de Historia de Ensenada, Ensenada. [ Links ]
Lindell J., Ngo A. and Murphy R.W. 2006. Deep genealogies and the mid-peninsular seaway of Baja California. Journal of Biogeography 33:1327-1331. [ Links ]
Minnich R.A., Franco-Vizcaíno E. and Salazar-Ceseña M. 2011. Distribution and regional ecology of Californian palm oases interpreted from Google Earth images. Aliso 29:1-12. [ Links ]
Moran R. 1952. The Mexican itineraries of T. S. Brandegee. Madroño 11:253-262. [ Links ]
Nelson E.W. 1921. Lower California and its Natural Resources. National Academy of Sciences, Washington, D.C. [ Links ]
Peinado M., Alcaraz F., Delgadillo J. y Aguado I. 1993. Fitogeografía de la península de Baja California, México. Anales Jardín Botánico de Madrid 51:255-277. [ Links ]
Pennell F.W. 1919. Eysenhardtia North American Flora 24:34-40. [ Links ]
Riddle B.R., Hafner D.J., Alexander L.F. and Jaeger J.R. 2000. Cryptic vicariance in the historical assembly of a Baja California peninsular desert biota. Proceedings of the National Academy of Sciences of the United States of America 97:14438-14443. [ Links ]
Risser P.G. 1995. The status of the science examining ecotones. BioScience 45:318-325. [ Links ]
Reyes-Coca S., Miranda-Reyes F. and García-López J. 1990. Climatología de la región noroeste de México. Precipitación: series de tiempo del valor total mensual y estadísticas del año climatológico. Reporte Técnico CIOFIT9001. CICESE, Ensenada, Baja California. [ Links ]
Rudkin C.N. 1952. Father Kino at La Paz, April 1683. Dawson's Bookshop, Los Angeles. [ Links ]
Shreve F. 1951. Vegetation of the Sonoran Desert. Carnegie Institution of Washington, Washinton, D.C. [ Links ]
Turner R.M., Bowers J.E. and Burgess T.L. 1995. Sonoran Desert Plants: An Ecological Atlas. University of Arizona Press, Tucson. [ Links ]
Upton D.E. and Murphy R.W. 1997. Phylogeny of the side-blotched lizards (Phrynosomatidae: Uta) based on mtDNA sequences: support for a midpeninsular seaway in Baja California. Molecular Phylogenetics and Evolution 8:104-113. [ Links ]
Vidal-Solano J.R., Paz-Moreno F.A., Demant A. and López-Martínez M. 2007. Ignimbritas hiperalcalinas del Mioceno medio en Sonora Central: revaluación de la estratigrafía y significado del volcanismo terciario. Revista Mexicana de Ciencias Geológicas 24:47-67. [ Links ]
Wehncke E.V., López-Medellín X. and Ezcurra E. 2009. Patterns of frugivory, seed dispersal and predation of blue fan palms (Brahea armata) in oases of northern Baja California. Journal of Arid Environments 73:773-783. [ Links ]
Wehncke E.V., López-Medellín X. and Ezcurra E. 2010. Blue fan palm distribution and seed removal patterns in three desert oases of northern Baja California, Mexico. Plant Ecology 208:1-20. [ Links ]
Wehncke E.V., López-Medellín X. and Ezcurra E. 2012. Blue fan palm oasis and sustainability: developing initial anticipated knowledge on a key desert geographic area of northern Baja California. In: International Hydrological Programme Division of Water Sciences Ed. Integrated Water Resource Management and Challenges of the Sustainable Development, pp. 327-341, UNESCO, Paris. [ Links ]
Whitcomb R.F., Robbins C.S., Lynch J.F., Whitcomb B.L., Klimikiewicz M.K. and Bystrak, D. 1981. Effects of forest fragmentation on avifauna of the eastern deciduous forest. In: Burgess R.L. and Sharpe D.M. Eds. Forest Island Dynamics in Man-Dominated Landscapes, pp. 125-205, Springer Verlag, New York. [ Links ]
Wiggins I. 1963. Botanical investigations in Baja California, Mexico. Plant Science Bulletin 9:1-6. [ Links ]
Wiggins I. 1980. Flora of Baja California. Stanford University Press, Stanford. [ Links ]
Zink R.M. 2002. Methods in comparative phylogeography, and their application to studying evolution in the North American aridlands. Integrative and Comparative Biology 42:953-959. [ Links ]
Zippin D.B. and Vanderwier J.M. 1994. Scrub community descriptions of the Baja California peninsula, Mexico. Madroño 41:85-119. [ Links ]

