










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.15 no.2 Chapingo jul./dic. 2009
Composición elemental de algunas especies de plantas silvestres mexicanas
Elemental composition of several wild mexican plants
J. C. Raya Pérez; C. L. Aguirre Mancilla
División de Estudios de Posgrado e Investigación, Instituto Tecnológico de Roque, km 8 Carr. Celaya–Juventino Rosas, Celaya, Guanajuato. C. P. 38110. MÉXICO. Correo–e: juancarlos.raya@gmail.com.
Recibido: 15 de enero, 2009
Aceptado: 23 de junio, 2009
RESUMEN
El conocer la composición elemental de las plantas puede ser útil desde varios puntos de vista, como el saber sus necesidades nutricionales y tener una idea del tipo de mantillo que se podría formar en un momento dado. En el caso específico del silicio, las plantas que lo acumulan pueden formar fitolitos, muy útiles en paleoecología y arqueología. Se recolectaron especies de plantas de varias partes del centro–occidente de México y se determinó su composición elemental mediante sonda dispersiva de rayos X acoplada a un microscopio electrónico de barrido. Las plantas semiparásitas (plantas que fotosintetizan pero que toman sus nutrientes de la planta que parasitan) acumulan cantidades considerables de iones como potasio y cloro para usarlos como osmolitos. Tillandsia recurvata, a pesar de ser epífita, acumula una buena cantidad de silicio, lo mismo que Cyperus sp., registrada como acumuladora de este elemento. Otras especies como Tagetes micrantha, de uso en la cocina mexicana, podrían contribuir en alguna medida al aporte de micronutrientes como potasio y magnesio.
Palabras clave: composición elemental, microanálisis, carbono, silicio, semi–parásitas, minerales.
ABSTRACT
Knowledge of plant elemental composition is useful to know their nutritional needs and the kind of litter created around them. Plants can to form silicon phytolites, which are useful in paleontology and archaeology. Plants were collected from central–occident of Mexico, its elemental composition were analyzed by Scanning Electron Microscope equipped with an Energy Dispersive X–ray microanalyzer SEM–EDS. Semi–parasitic plants (Photosynthetic plants than take nutriments from the host plant) accumulated high quantities of potassium and chloride ions to use as osmolites. Tillandsia recurvata, which is epiphyte, accumulates high quantities of Silicon, as well as, Cyperus sp., this one reported as Silicon accumulator. Tagetes micrantha that is used in Mexican food could contribute with mineral nutrients like potassium and magnesium.
Keywords: elemental composition, microanalysis, carbon, silicon, semi–parasitic, minerals.
INTRODUCCIÓN
Las plantas obtienen los minerales que necesitan desde la solución del suelo con la cual se hallan en contacto. Algunos, como el nitrógeno (N) pueden alcanzar entre 0.5 y 6 % del peso seco de la planta, en tanto otros, como el cobalto (Co), hierro (Fe), manganeso (Mn) y zinc (Zn) están presentes en apenas partes por millón (Epstein, 1994). Los requerimientos de muchas de las plantas cultivadas han sido analizados con detalle desde hace tiempo, especialmente a partir de 1860 que se introdujo la hidroponia, lo que ha permitido llevar a cabo, con un mejor control este tipo de estudios (Epstein, 1999). Por ejemplo, la restricción de fosforo en cacahuate (Arachis hypogaea) detiene su crecimiento y provoca defoliación de las hojas basales que se tornan amarillas. La falta de azufre (S) en esta leguminosa impide la nodulación y la falta de calcio (Ca) afecta severamente la producción de frutos (Bledsoe y Harris, 1950). Desde el punto de vista nutricional también resulta interesante saber que en la semilla de frijol (Phaseolus vulgaris) el contenido de molibdeno (Mo) difiere por un orden de magnitud entre suelos ricos y pobres en este elemento. En el grano de sorgo (Sorghumhalepense), las concentraciones de fósforo (P), potasio (K), y magnesio (Mg) decrecen significativamente cuando disminuye la suplementación de estos elementos (Tyler y Zohlen, 1998).
La disponibilidad de nutrientes también puede afectar el balance competitivo entre las especies de plantas de un ecosistema (Hogh–Jensen et al., 2001) y hay diferencias en cuanto a la composición entre las plantas calcícolas y calcifugas. Las calcícolas tienen la capacidad de exudar ácidos orgánicos para solubilizar elementos como el fósforo, el fierro y el manganeso; aunque en elementos como potasio, magnesio, zinc, fósforo y azufre, permanecen relativamente constantes los contenidos entre unas y otras (Tyler y Zohlen, 1998). En Brassica la concentración de calcio varía entre 1.7–3.3 % y la de magnesio de 0.35–0.8 % y las mismas variedades de plantas muestran cantidades semejantes en condiciones de campo e invernadero, lo que indica que estas características tienen un componente hereditario (Broadley et al., 2008). El silicio (Si) es un elemento sobre el cual hasta hace relativamente poco se llamó la atención, es importante para aminorar o aliviar la toxicidad de otros elementos presentes en exceso como el aluminio (Al) o el manganeso (Mn) y para algunas plantas como el arroz (Oryza sativa) y la caña de azúcar (Saccharum officinarum) resulta esencial (Gunes et al., 2007a; Gunes et al., 2007b). Se han buscado modelos biológicos que sirvan como base para estudiar la formación de estructuras con este elemento, siendo a la fecha las diatomeas los organismos más estudiados al respecto (Ludwig et al., 1996; Vrieling et al., 2007). El microanálisis usando sonda dispersiva de rayos X es ya una técnica muy usada en muestras biológicas (Thibaut et al., 1978; Liu et al., 2007). El objetivo del trabajo fue buscar posibles especies candidatas que acumularan Silicio (Si) para usarse como modelo en la absorción, traslocación y acumulación de este elemento. Esto reviste interés por el papel del Si en contrarrestar los efectos tóxicos de algunos elementos como el aluminio (Al) y el manganeso (Mn); por conferir resistencia contra plagas y enfermedades e incluso por la formación de nanoestructuras utilizando como catalizadores compuestos orgánicos, enzimas u organismos (Epstein, 1999; Aguirre et al., 2007).
MATERIALES Y MÉTODOS
Las plantas Cyperus sp., Tagetes micrantha, Mangifera indica (Mango), Struthantus quercicola (correhuela) y Cuscuta umbellata fueron recolectadas en Uruapan, Michoacán, México; en el caso particular del champiñón (Agaricus campestris), fue adquirido en el mercado local, se decidió incluir un hongo por su papel de ayudador de las plantas a la adquisición de nutrientes. Las plantas fueron llevadas al laboratorio y secadas a 40 °C hasta peso constante. Se molieron usando un molino de bolas marca Retsch MM301 y se hicieron pastillas, mediante el uso de una prensa hidráulica marca Carver. Estas pastillas fueron usadas para los microanálisis, tres por cada pastilla. Las plantas de Almutaster pauciflorus y Aster spinosus fueron recolectadas en Irapuato, México, junto a una corriente de agua. Las de Tillandsia recurvata se recolectaron en San Luis de la Paz, México, creciendo sobre troncos de capulín (Prunus serotina) o acacia (Acacia sp.). El microscopio electrónico de barrido utilizado fue marca JEOL 6480 LV acoplado a una sonda dispersiva de rayos X INCAx sigth Oxford instruments.
RESULTADOS Y DISCUSIÓN
Para la vegetación se considera como referencia general que contienen alrededor de 50 % de carbono; sin embargo, este valor puede variar desde 34.6 % en las hierbas de las praderas a 52.8 % en los árboles (Figueroa–Navarro et al., 2005). En el presente estudio se encontró un alto contenido de carbono en las especies de Almutaster y Aster (51 %) (Figuras 1 y 2), lo cual se podría explicar por que son plantas prácticamente sin hojas y con tallos muy duros. En otras especies, como anís (Tagetes micrantha) (45.6 %) (Figura 3) y Tillandsia (48.5 % cuando crece sobre capulín y 46.81 % cuando crece sobre acacia) (Figuras 4 y 5), muestran la tendencia señalada por otros autores respecto a que las hierbas presentan un menor contenido que los árboles. Los valores aquí mencionados refuerzan la idea de que una estimación de 50 % es adecuada, aunque se pueden encontrar diferencias entre especies y formas de vida: hierbas, árboles y arbustos (Cuadro 1) (Figueroa–Navarro et al., 2005).





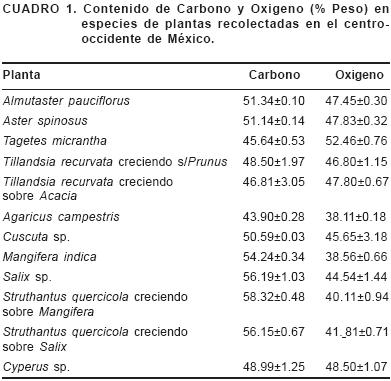
En plantas, el porcentaje de calcio y magnesio oscila entre 0.02–13.1 % para el primero y de 0.03–3.2 % para el segundo (Broadley et al., 2008). La cantidad de fósforo conocida varía de entre 0.15–0.5 %; vale la pena mencionar que el champiñón (Figura 6), un hongo, presenta una alta acumulación de este elemento. Si la alta concentración de fósforo se debe al tipo de sustrato usado para el cultivo es algo que se ignora todavía, y que es importante considerar por ser también una especie comestible; en su forma de micorriza facilita a las plantas la adquisición de algunos nutrimentos como el fósforo y el potasio. De acuerdo con los resultados obtenidos en el presente estudio, estos hongos son capaces de almacenar cierta cantidad de nutrimentos, como el fósforo, en su cuerpo fructífero. Los valores que encontramos para las plantas caen dentro de los valores citados (Epstein, 1994; Broadley et al., 2008). La concentración de azufre mencionada en otros estudios (Epstein, 1994) oscila entre 0.1–1.5 %, intervalo dentro del cual caen los valores obtenidos en este trabajo. En el caso del potasio, de importancia como osmolito para las plantas y también desde el punto de vista de nutrición humana, se indican valores de 0.8–8 % (Epstein, 1994). Los resultados muestran que Struthantus quercicola (correhuela) alcanza un valor de 1.1 %, el champiñón de 4.85 % y Cuscuta (Figura 7) de 0.32 %; la correhuela y la Cuscuta se sabe que usan el potasio como osmolito, a fin de atraer el flujo de savia de las plantas que parasitan, hacia sus propios tejidos, por ello presentan concentraciones considerables de este ión.


Para la Cuscuta se mencionan valores de acumulación de silicio de entre 0 y 0.63 % (Hodson et al., 2005), intervalo dentro del cual se encuentran los hallados en el presente estudio (0.09 %, Figura 7). Para Cyperus se encontró un valor de 0.26 %; se han registrado valores desde 3.42 % para Cyperus alternifolius hasta 0.79 % para C. alopecuroides (Hodson et al., 2005) El valor determinado fue menor y amplía el intervalo en cuanto al contenido de silicio en este género.
En el caso del anís (Tagetes micrantha) el contenido de magnesio es alto (0.28 %), dado que se añade a atoles o en la preparación de tizanas, puede considerarse una fuente importante de este elemento, que podría ser un ión cuya carencia es crónica en un alto porcentaje de la población humana (Killilea y Ames, 2008).
El silicio se presenta de manera importante en Tillandsia recurvata, con más de 1 %. Hodson et al. (2005) mencionan valores de 0.256 % para Tillandsia usneoides. Sería interesante hacer análisis elemental de más especies dentro del género y abarcar diferentes regiones geográficas, de ampliar y ahondar en este género respecto a su contenido de silicio. Las Figuras 4 y 5 muestran que la composición de la planta se mantiene constante tanto al crecer sobre acacia como al crecer sobre capulín. En la calabaza (Cucurbita sp.) y el arroz, se sabe que hay un loci que interviene en la acumulación de silicio (Piperno et al., 2002; Wu et al., 2006). Entre las plantas acumuladoras de silicio se halla el arroz, que alcanza 6 % y se ha convertido en uno de los modelos para estudiar la acumulación y el papel del silicio en los vegetales; además los fitosilicatos se encuentran entre los principales componentes del suelo que interaccionan con la materia orgánica (Hogh–Jensen et al., 2001; Ma y Yamaji, 2006). Es probable que Tillandsia pueda servir como un modelo alternativo para estudiar la absorción y transporte de silicio.
La única especie, de las analizadas, que presentó sodio (Na) en su composición fue Tillandsia, esto resulta sorprendente dado que la mayoría de las plantas excretan este ión, aunque se ha encontrado que las que tienen fotosíntesis tipo C4 requieren de sodio (Ohta et al., 1987).
Los resultados de este trabajo refuerzan la idea que hay un gran componente hereditario en cuanto a la acumulación de elementos minerales por parte de las plantas. Así se podría buscar dentro de las familias o géneros, plantas que acumulen algunos iones en específicos dependiendo de sus relaciones filogenéticas; en este caso se observa que las especies de Almutaster y Aster analizadas muestran una composición elemental muy semejante, acorde con lo postulado por otros autores (Hodson et al., 2005). Lo mismo se observa con Tillandsia, sea que crezca sobre el tronco de Prunus o de Acacia, su contenido mineral no cambia. El conocimiento de la composición elemental de las plantas puede ayudar a tratar de dilucidar las relaciones intra e interespecíficas en las comunidades (Hogh–Jensen et al., 2001), a reconstruir, con ayuda de los fitolitos, las especies presentes que existían o aquellas cultivadas por pueblos antiguos (Piperno et al., 2002; Fernández–Honaine et al., 2006). También puede ayudar a definir la variación natural que existe entre especies, e incluso entre variedades, en cuanto a la acumulación de algunos macro y micro elementos, para usarla convenientemente, incluso para hacer una mejor estimación del contenido de elementos en las plantas, por ejemplo de carbono (Tyler y Zohen, 1988; Fang, 2008).
CONCLUSIONES
Se presentan datos sobre la composición de varias especies de plantas comunes en México, algunas de uso tradicional, se muestra que el microanálisis con sonda de rayos X es un método disponible y válido para conocer la composición elemental de material biológico y se propone a Tillandsia recurvata como modelo de planta acumuladora de silicio. Con este método obtuvimos un promedio en el contenido de carbono para las plantas analizadas de 50.98 %, valor acorde con la estimación de 50 %.
De acuerdo a lo mencionado por otros autores, encontramos que Tillandsia sin importar la especie de árbol sobre la que crece muestra una composición elemental muy parecida; y las especies de los géneros Almutaster y Aster también muestran una composición semejante, confirmando que la relación filogenética cercana influye en el contenido mineral de las plantas.
AGRADECIMIENTOS
Expresamos nuestro agradecimiento a José García por su apoyo en determinar las especies de Aster.
LITERATURA CITADA
AGUIRRE–MANCILLA, C.; CHÁVEZ–BARCENAS, T.; GARCÍA–SAUCEDO P.; RAYA–PÉREZ, J. C. 2007. El silicio en los organismos vivos. Interciencia 3: 504–509. [ Links ]
ROGER W. BLEDSOE; HENRY C. HARRIS. 1950. The influence of mineral deficiency on vegetative growth, flower and fruit production, and mineral composition of the peanut plant. Plant Physiology 25: 63–77 [ Links ]
BROADLEY, M. R.; HAMMOND, J. P.; KING, G. J.; ASTLEY, D.; BOWEN, H. C.; MEACHAM, M. C.; MEAD,A.; PINK, D. A. C.; TEAKLE, G. R.; HAYDEN, R. M.; SPRACKLEN, W. P.; WHITE P. J. 2008. Shoot calcium and magnesium concentrations differ between subtaxa, are highly heritable, and associate with potentially pleiotropic loci in Brassica oleraceae. Plant Physiology 146: 1707–1720. [ Links ]
EPSTEIN, E. 1999. Silicon. Annual Review of Plant Physiology and Plant Molecular Biology 50: 641–664. [ Links ]
EPSTEIN, E. 1994. The anomaly of silicon in plant biology. Proceedings of the National Academy of Sciences of the United States of America 91: 11–17. [ Links ]
FANG, Y.; WANG, L.; XIN, Z.; ZHAO, L.; AN, X.; HU, Q. 2008. Effect of foliar application of zinc, selenium, and iron fertilizers in nutrients concentration and yield of rice grain in China. Journal of Agricultural and Food Chemistry 56: 2079–2084. [ Links ]
FERNANDEZ–HONAINE, M.; ZUCOL, A. F.; OSTERRIETH, M. L. 2006. Phytolit assemblages and systematic associations in grassland species of south–eastern pampean plains, Argentina. Annals of Botany 98: 1155–1165. [ Links ]
FIGUEROA–NAVARRO, C.; ETCHEVERS–BARRA, J. D.; VELAZQUEZ–MARTÍNEZ, A.; ACOSTA–MIRELES, M. 2005. Concentración de carbono en diferentes tipos de vegetación de la Sierra Norte de Oaxaca. Terra Latinoamericana 23: 57–64. [ Links ]
GUNES, A.; INAL, A.; BAGCI, E. G.; COBAN, S.; SAHIN, O. 2007a. Silicon increases boron tolerance and reduce oxidative damage of wheat growth in soil with excess boron. Biologia Plantarum 51: 571–574. [ Links ]
GUNES,A.; INAL, A.; BAGCI, E. G.; PILBEAM, D. J. 2007b. Silicon–mediated changes of some physiological and enzymatic parameters symptomatic for oxidative stress in spinach and tomato grown in sodic–B toxic soil. Plant and Soil 290: 103–114. [ Links ]
HODSON, M. J.; WHITE, P. J.; MEAD, A.; BROADLEY, M. R. 2005. Phylogenetic variation in the silicon composition of plants. Annals of Botany 96: 1027–1046. [ Links ]
HOGH–JENSEN, H.; FABRICIUS, V.; SCHJOERRING, J. K. 2001. Regrowth and nutrient composition of different plant organs in grass–clover canopies as affected by phosphorus and potassium availability. Annals of Botany 88: 153–162. [ Links ]
KILLILEA, D. W.; AMES, B. N. 2008. Magnesium deficiency accelerates cellular senescence in cultured human fibroblasts. Proceedings of the National Academy of Sciences of the United States of America. 105: 5768–5773. [ Links ]
LIU, D.; KOTTKE, I.; ADAM, D. 2007. Localization of cadmium in the root cells of Allium cepa by energy dispersive X–ray analysis. Biologia Plantarum 51: 363–366. [ Links ]
LUDWIG, M.; LIND, J. L.; MILLER, E. A.; WETHERBEE, R. 1996. High molecular mass glycoproteins associated with the siliceous scales and bristles of Mallomonas splendens (Synurophyceae) may be involved in cell and maintenance. Planta 199: 219–228. [ Links ]
MA, J. F.; YAMAJI, N. 2006. Silicon uptake and accumulation in higher plants. Trends in Plant Science 11: 392–397. [ Links ]
OHTA, D.; MATOH, T.; TAKAHASHI, E. 1987. Early responses of sodium–deficient Amaranthus tricolor L. plants to sodium application. Plant Physiology 84: 112–117. [ Links ]
PIPERNO, D. R.; HOLST, I.; WESSEL–BEAVER, L.; ANDRES, T. C. 2002. Evidence for the control of phytolith formation in Cucurbita fruits by the hard ring (Hr) genetic locus: archaeological and ecological implications. Proceedings of the National Academy of Sciences of the United States of America 99: 10923–10928. [ Links ]
THIBAUT, M.; ANSEL, M.; DE AZEVEDO–CARNEIRO, J. 1978. A microanalysis approach to investigate problems encountered in mycology. The American Journal of Pathology 90: 23–32. [ Links ]
TYLER, G.; ZOHLEN, A. 1998. Plant seeds as a mineral nutrient resource for seedlings A comparison of plants from calcareous and silicate soils. Annals of Botany 81: 455–459. [ Links ]
VRIELING, E. G; SUN, Q.; TIAN, M.; KOOYMAN, P. J.; GIESKES, W. W. C.; VAN SANTEN, R.A.; SOMMERDIJK, N. A. J. M. 2007. Salinity–dependent diatom biosilification implies an important role of external ionic strength Proceedings of the National Academy of Sciences of the United States of America 104: 10441–10446. [ Links ]
WU, Q. S.; WAN, X. Y.; SU, N.; CHENG, Z. J.; WANG, J. K.; LEI, C. L.; ZHANG, X.; JINAG, L.; MA, J. F.; WAN, J. M. 2006. Genetic dissection of silicon uptake ability in rice (Oryza sativa L.). Plant Science 171: 441–448. [ Links ]

