











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Bats of the widely distributed genus Myotis are an excellent model for studies of diversification and historical biogeography (Stadelmann et al. 2007; Ruedi et al. 2013). With ca. 134 species (www.mammaldiversity.org), Myotis is one of the more remarkable mammalian radiations worldwide. Although long considered to be a classic example of an adaptive radiation in temperate regions, more detailed studies of Neotropical (Larsen et al. 2012a, b) and Afrotropical species (Patterson et al. 2019) using molecular data are now uncovering substantial cryptic diversity (Bickford et al. 2007) in tropical regions.
Based on previous systematic revisions (La Val 1973; Hernandez-Meza et al. 2005; Wilson 2008; Moratelli and Wilson 2014; Mantilla-Meluck and Muñoz-Garay 2014), the nominal species M. keaysi was partitioned into two subspecies: M. k. keaysi J. A. Allen 1914, distributed in the Andes of Colombia, Colombian Caribbean, Perú, Ecuador, Bolivia, and Argentina above 1,100 m, with most specimens known from above 2,000 m and M. k. pilosatibialis (La Val 1973), occurring in northern Venezuela, the island of Trinidad, Colombian Caribbean, eastern cordillera of the Colombian Andes, and elsewhere from southern México, southeastward through Central America into northwestern Panamá. Both subspecies are known to occur in sympatry in Caribbean Colombia and the eastern cordillera of Colombia. Although the phylogenetic placement for both is still pending, Mantilla-Meluck and Muñoz-Garay (2014) recognized pilosatibialis as distinct at the species level based on morphology. Based on those findings, Moratelli and Wilson (2014) recommended reassigning the specimens previously assigned to M. keaysi by Stadelmann et al. (2007) and Ruedi et al. (2013) from Yucatán (México) to M. pilosatibialis. Furthermore, Moratelli et al. (2016) and Moratelli et al. (2017) were unable to confidently identify these specimens from Yucatán, México and assigned provisionally a “cf .” (Latin, confer), preceding the specific epithet.
Myotis keaysi as envisioned by Stadelmann et al. 2007 and Ruedi et al. 2013 was identified as a monophyletic clade with considerable geographic structure and at least three different lineages (Larsen et al. 2012a; Chaverri et al. 2016), which were named as follows: M. keaysi (Yucatán Peninsula, México), M. cf. keaysi 2 (México, El Salvador, Guatemala), and an unnamed clade from the mountain tops of Cordillera Oriental from Ecuador and Panamá (Clare et al. 2011; Larsen et al. 2012b) that was later suggested to be M. keaysi (Costa Rica) in Chaverri et al. (2016).
In this study, we provided the phylogenetic placement for M. keaysi str. and M. pilosatibialis str. in the Myotis Neotropical radiation and under this phylogenetic framework, we reviewed specimens from the unnamed clade from mountain tops of Chiriquí (Panamá) and Cordillera Oriental (Ecuador). We found that these specimens are distinct from other species of Neotropical Myotis and described this new species based on the General Lineage Concept (GLC), which uses the concept of species as separately evolving metapopulation lineages that can be recognized using diverse secondary recognition criteria (De Queiroz 2007). We apply the criteria used by Florio et al. (2012) as follows: a) identify lineages based on clades from analysis of multiple molecular markers b) employ multivariate procedures such as principal component analysis (PCA) and discriminant function analysis (DFA) to determine morphological variation associated with these phylogenetic groups, and c) use climatic analysis to evaluate the environment and geographic space occupied by the groups supported by covariation of genetic and morphological evidence.
Methods
Specimens in Panamá were collected under an ongoing project entitled “Caracterización de la epidemiología y ecología de enfermedades zoonóticas trasmitidas por vectores (emergentes y reemergentes) en áreas silenciosas y conocidas de Panamá” signed by Instituto Conmemorativo Gorgas and Ministerio del Medio Ambiente de Panamá, with permits SC/A-50-1 and SEX/A-1-19. In Ecuador, specimens were collected under the project “Caracterización de la diversidad biológica y genética de los mamíferos del Ecuador”, signed by Pontificia Universidad Católica del Ecuador and Ministerio del Medio Ambiente (MAE), with permits (MAE-DNB-CM-2016-004 and 70-12-2017- DPAN/MAE). Specimens in both countries were collected with ground-level mist netting and field methods generally followed Galbreath et al. (2019), under guidelines of the American Society of Mammalogists (Sikes et al. 2016) and approved by the University of New Mexico Animal Care and Use Committee.
Specimens examined. Specimens directly examined (n= 72) for this research include representatives of Central and South American Myotis, including type specimens (Appendix I). These comprise fluid-preserved specimens, stuffed skins, and skulls deposited in the following institutions: American Museum of Natural History (AMNH); Field Museum of Natural History, Chicago (FMNH); Natural History Museum, University of Kansas (KU); Natural History Museum of Los Angeles County, Los Angeles (LACM); Louisiana State University Museum of Natural Science (LSUMZ); Museum of Vertebrate Zoology, University of California, Berkeley (MVZ); Museum of Southwestern Biology, University of New Mexico (MSB); National Museum of Natural History, Smithsonian Institution (USNM); Museo de Zoología, División de Mastozoología, Pontificia Universidad Católica del Ecuador (QCAZ); Royal Ontario Museum (ROM); Sam Noble Oklahoma Museum of Natural History, University of Oklahoma (SNOMNH); Texas A&M Biodiversity Research and Teaching Collection, College Station (TCWC); Museum of Texas Tech University (TTU; Dunnum et al. 2018). These specimens were identified according to Wilson (2008), Moratelli and Wilson (2011), Moratelli et al. (2013), Moratelli et al. (2017).
Measurements. All measurements are in millimeters (mm) or grams (body mass) and are from adults. The lengths of head and body (HB), tail, hind foot, ear, and the body weight (mass) were recorded from skin labels and are reported to the nearest millimeter or nearest gram. Forearm length (FA) and third metacarpal length (3ML) were directly measured from specimens. Measurements were taken using digital calipers accurate to 0.02 mm. Craniometric measurements were taken under a binocular microscope at low magnification (usually 6x). These dimensions were recorded and analyzed to the nearest 0.01 mm, but values were rounded off to 0.1 mm throughout the text because this is the smallest unit allowing accurate repeatability with calipers (Voss et al. 2013).
Measurements include four external, 14 cranial, and two mandibular dimensions. The measurements and abbreviations are defined as follows: forearm length (FA), from the elbow to the distal end of the forearm including carpals; third metacarpal length (3ML), from the distal end of the forearm including carpals to the distal end of the third metacarpal; length of dorsal hairs (LDH), from the base to the tip of the hair in the fur between the scapulae; length of ventral hairs (LVH) in fur at mid thorax; greatest length of the skull (GLS), from the apex of the upper internal incisors to the occiput; condylocanine length (CCL), from the anterior surface of the upper canines to a line connecting the occipital condyles; condylobasal length (CBL), from the premaxillae to a line connecting the occipital condyles; incisive length (CIL), from the apex of upper internal incisors to a line connecting the occipital condyles; basal length (BAL), from the apex of upper internal incisors to the ventral margin of the foramen magnum; zygomatic breadth (ZB), greatest breadth across the outer margins of the zygomatic arches; mastoid breadth (MAB) greatest breadth across the mastoid region (MAB); braincase breadth (BCB), greatest breadth of the globular part of the braincase; interorbital breadth (IOB), least breadth between the orbits; postorbital breadth (POB), least breadth across frontal posterior to the postorbital bulges; breadth across canines (BAC), greatest breadth across outer edges of the crowns of upper canines, including cingulae; breadth across molars (BAM), greatest breadth across outer edges of the crowns of upper molars; maxillary toothrow length (MTL), from the upper canine to M3; molariform toothrow length (M1-3), from M1 to M3; mandibular length (MAL), from the mandibular symphysis to the condyloid process; and mandibular toothrow length (MAN), from the lower canine to m3. Measurements and abbreviations are further defined in Moratelli et al. (2013). Descriptive statistics (mean and range) were calculated for all dimensions. The cranial index (CRI= (((IOB + BCB) x GLS)/2)) and a modification of the maxillary index (MXI = (((BAC + BAM) x MTL)/2)) were as used by Baud and Menu (1993) and López-González et al. (2001). We used capitalized colors from (Ridgway 1912).
Principal component analysis (PCA) and discriminant function analysis (DFA) were used to examine overall patterns of skull size and shape variation among samples. We selected the following cranial dimensions representing different axes of length and width of the skull, rostrum and mandible: GLS, CCL, CBL, CIL, BAL, MAB, BCB, IOB, POB, BAC, BAM, MTL, M1-3, MAL, MAN. To obtain a more balanced design for multivariate analysis, we selected a minimum of 4 and maximum of 10 adult specimens, totaling 33 individuals (17 females and 16 males). Males and females were pooled together to enhance analysis. PCA was used to summarize trends in size and shape variation (total data set was considered a unique sample). We conducted a varimax rotation of the loading matrix after PCA in order to have more interpretable factors with a simpler structure that can be obtained using orthogonal rotations. We used rotated PCA scores to test for statistical significance of difference among species. This was assessed by multiple analysis of variance MANOVA, with Pillai’s Trace. This previous analysis was followed by one-way ANOVA. A post-hoc multiple comparison Bonferroni corrected approach was used to evaluate pairwise differences among species (Rice1989).
DFA was used to assess craniometric characters that best discriminate samples, with a priori identification of samples (Moratelli et al. 2013). For DFA, probabilities for misclassification rates were also assessed and individuals were resigned using a jackknife procedure. Missing data values were estimated using Amelia R package (0 % of total data set) from raw data set (Honaker et al. 2011). We checked for normality assumptions and measurements were log transformed (Zar 1998). Finally, variance-covariance matrices were computed using all variables.
Statistical analyses were performed in software R (R Core Team 2020). We used the following R packages as follows: MASS (DFA analysis), stats (PCA analysis), ggplot2 and ggord (graphics and visualization), factoextra (eigenvalues extraction), psych (varimax rotation), car (MANOVA analysis), rstatix (summary statistics, T-test, ANOVA, adjust P-values for multiple comparisons, formatting and adding significant symbols; Venables and Venables and Ripley 2002; Wickham 2016; Beck 2017; Fox and Weisberg 2019; Revelle 2020; Kassambara 2020; Kassambara and Mundt 2020.
We contrasted the skull size and shape of M. sp. n. (n= 10) from Chiriquí (Panamá) and Cordillera Oriental (Tungurahua Province, Ecuador); M. pilosatibialis str., including the holotype specimen (n = 8); M. keaysi str. from Puno and Cuzco, Perú (n = 5); M. oxyotus. gardneri, including the holotype specimen (n = 6) from Costa Rica and Panamá; and specimens provisionally labelled as M. cf. pilosatibialis (n = 4) from Quintana Roo, Yucatán Peninsula, México, hereafter called M. sp. We included M. pilosatibialis str. (El Salvador, Guatemala, Honduras and México) and M. sp. (Quintana Roo, Yucatán, México) due to its close morphological similarity and phylogenetic proximity (Larsen et al. 2012a; Moratelli et al. 2016; Moratelli et al. 2017). Similarly, we included specimens that morphologically match M. oxyotus gardneriLa Val 1973, which was recorded in syntopy with the newly identified lineage at Chiriquí Province (Panamá) and Valle del Silencio (Costa Rica). Likewise, we included M. keaysi str. from Perú, due to resemblance in external dimensions, cranial morphology and fur traits. Qualitative traits employed here to characterize and distinguish species follow Moratelli et al. (2013). Capitalized color nomenclature follows Ridgway (1912).
DNA Extraction, Sequencing, Editing, and Assembly of partial cytochrome b and nuclear genes. To obtain DNA sequence data, whole DNA was extracted from liver, muscle or wing tissues, following a guanidinium isothiocyanate extraction protocol, E.Z.N.A. Tissue DNA Kit (Omega), and DNeasy Blood & Tissue Kit (Qiagen). Samples were quantified using Nanodrop® ND-1000 (NanoDrop Technologies, Inc) or Qubit fluorometer kit (Invitrogen), resuspended and diluted to 25 ng/ul in ddH2O prior to amplification.
We relied on different primers and PCR protocols, which are reported as follows: cytochrome b in Hoffmann and Baker (2001); Larsen et al. (2012b); and Naidu et al. (2012); nuclear exon and intron genes in Matthee and Davis (2001); Eick et al. (2005); Larsen et al. (2012b); Lack et al. (2010); Roehrs et al. (2010); and Salicini et al. (2011). We targeted ~ 710 bp of cytochrome-b sequence (n = 11), 1, 038 bp of nuclear exon, (RAG2) recombination activating gene II sequence (n = 13), and intron region of other 3 genes, 402 bp of protein kinase C, iota sequence (PRKCI, n = 14), 414 bp signal transducer and activator of transcription 5A sequence (STAT5A, n = 14), and 475 bp of thyrotropin sequence (THY, n = 14). Sequencing was conducted using Applied Biosystems 3110 Sequencer of the molecular biology facility at the University of New Mexico (UNM). In Ecuador, successful amplified PCR products were sent for sequencing to the commercial laboratory Macrogen Inc in Seoul, South Korea. Editing and assembly of sequences were performed with Geneious Prime (BioMatters Ltd. 2020).
DNA Extraction and Sequencing of cytochrome oxidase c subunit I. We retrieved partial sequences of cytochrome c oxidase subunit I (657 bp) from mitochondrial genomes for the following species: M. oxyotus gardneri (n = 1), M. pilosatibialis str. (n = 2), M. sp. (n = 1, Quintana Roo, Yucatán, México), and M. sp. n. (n = 2). DNA was extracted using standard manual extraction methods such as DNeasy Blood & Tissue Kit (QUIAGEN, Hilden, Germany) following manufacturers guidelines, as well as one open method based on magnetic particle using KingFisherTM Duo (Thermo Fisher Scientific). DNA was incorporated into double stranded DNA short fragment libraries built following BEST protocol (Mak et al. 2017; Carøe et al. 2018) using BGISEQ-500 adapters. Libraries were sequenced using 100 base paired end read chemistry on a BGISEQ-500 sequencer machine (BGI-Copenhagen).
Mitogenomes assembly and annotation. Low quality base reads, missing bases and adapters were trimmed followed by adapters removal using Adapter Removal v2 (Lindgreen 2012; Schubert et al. 2016). We built mitochondrial genomes using NOVOPlasty (Dierckxsens et al. 2017) using as reference the mitochondrial genome of Myotis lucifugus accession number (NC_029849.1). Furthermore, annotation was carried on with MitoZ (Meng et al. 2019).
DNA Alignments and Phylogenetic analysis. Alignments of sequences were performed with Geneious Prime (BioMatters Ltd. 2020), using MUSCLE (Edgar 2004). Phylogenetic analysis of mitochondrial and nuclear sequences was conducted with newly generated sequences and sequences retrieved from GenBank for Neotropical species of Myotis previously known to form a monophyletic clade (Ruedi et al. 2013). We included previously generated sequences from GenBank for cytochrome-b for 91 individuals (Ruedi and Mayer 2001; Kawai et al. 2003; Rodriguez and Ammerman 2004; Stadelmann et al. 2004; Kawai et al. 2006; Stadelmann et al. 2007; Baird et al. 2008; Larsen et al. 2012a, b; Patterson et al. 2019), including unpublished data by Parlos et al. (2008); COXI for 51 individuals used in Chaverri et al. (2016); RAG2 for 14 individuals; PRKCI for 13 individuals; STAT5A for 13 individuals; and THY for 14 individuals (Lack et al. 2010; Roehrs et al. 2010)
Outgroup selection. For cytochrome b phylogeny, we selected three sequences of M. gracilis, one sequence of M. brandtii and one sequence of M. yanbarensis, which are sister species to Neotropical Myotis. For our cytochrome c oxidase subunit I phylogeny, four sequences of M. brandtii, and one sequence of M. lucifugus were used. In our species tree analysis, we included myotine bats from the Nearctic clade as well, therefore we selected the eastern Palearctic lineage M. cf. ikonnikovi as the outgroup.
Maximum likelihood trees were generated using IQ-TREE with 100 bootstraps and 1000 replicates (Nguyen et al. 2015; Trifinopoulos et al. 2016). We used the program ModelFinder (Kalyaanamoorthy et al. 2017) with Bayesian information criteria (BIC) for selecting a nucleotide substitution model for cytochrome-b, cytochrome oxidase c subunit I, exon and intron regions. Heterozygous nuclear introns alleles were statistically resolved using PHASE 2.1.1 (Stephens et al. 2001) prior to the inclusion in further analyses. We used SEQPHASE web server (Flot, 2010) to generate the input files for PHASE. The Bayesian analysis was conducted in MrBayes v.3.2 for partial cytochrome b and cytochrome c oxidase subunit I (Ronquist et al. 2012). The search started with a random tree and the Markov chain was run for 10 million generations with trees sampled every 1,000 generations in two replications. Default values were kept for the “relburning” and “burninfrac” options in MrBayes, therefore the first 25.000, 00 generations were discarded as burn in, and posterior probability estimates of all model parameters were based on the remaining trees. Tree convergence and stationary was accessed in the Bayesian analysis by plotting the likelihood values in Tracer v1.7.1 (Rambaut et al. 2018).
We follow Moratelli et al. (2017) when designating nodal support in the mitochondrial gene trees; for ML analyses there is a strong support for bootstraps values above ≥ 75 %, moderate support for values > 50 % and < 75 %; and negligible for support for values ≤ 50 %. For the BI analysis, there are two categories, with significant support in cases in which a node posterior probability was ≥ 0.95, and insignificant or negligible support for posterior probability values < 0.95.
Identification of molecular synapomorphies. For the identification of unique molecular synapomorphies for the newly identified lineage, we generated maximum parsimony trees from mitochondrial data sets and intron regions of 3 nuclear genes using PAUP* (Swofford 2003). For these analyses, characters were treated as unordered and equally weighted. We performed a heuristic search with random addition of sequences and tree bisection-reconnection branch swapping. We set maxtree limit to 1000, with the goal of applying a maxtree limit of 100 to each 10 random addition sequence replicate. To measure clade support, 1000 bootstrap replicates were performed on a 50 % majority-rule consensus tree. We used the command describetrees and apolist with delayed transformation (DELTRAN) to obtain a list of molecular synapomorphies. To polarize character state transformation, we used outgroups previously used in maximum likelihood and Bayesian analyses.
Genetic distances. We calculated K2P distances (these were computed for comparisons with previous molecular studies on Neotropical Myotis; e. g, Larsen et al. 2012a, b) expressed as percentages and pairwise genetic distances using uncorrected sequence divergence (p-distances), and modeled in MEGAv10.17 (Kumar et al. 2016) for mitochondrial genes.
Species tree and species delimitation. Under the multispecies coalescent model, we inferred both species tree and species delimitation. For species tree inference we used *BEAST (Heled and Drummond 2010) method using the software suite available in BEAST 2.6.2 (Bouckaert et al. 2019). We used three phased intron and one exon alignments with substitution, clock, and tree models unlinked among all loci. All loci were assigned the lognormal relaxed-clock model using a Yule prior and linear with constant root population size model. We decided not to include mtDNA, due to its potentially strong influence on species tree inference given its higher variability and that assumptions of lower ploidy are not always met (which is modelled by *BEAST; McLean et al. 2016). We ran the analysis for 1 x 108 generations in two replicates, saving the results every 10,000 generations. The first 10 % of each run was discarded as burn-in and assembled using LOGCOMBINER v.1.10.4 to produce a maximum clade credibility tree in TREEANNOTATOR v1.10.4. Likewise, we used Tracer v.1.7.1 to access convergence and stationarity of model parameters based on ESS values and examination of tree files.
We used STACEY v1.2.1 (Species Tree and Estimation classification, Yarely), implemented in BEAST 2.6.2 (Jones 2017; Bouckaert et al. 2019). This method requires no prior assignment of individual to species, and no guided tree. We ran the analysis for 2 x 108 generations in two replicates. After completion, we used Species Delimitation Analyzer (Jones 2014; Jones 2015) to process log files and find the distribution over species assignments under two collapse height priors (collapse 0.0001 and 0.0005). Tracer v.1.7.1 was used to assess convergence and stationarity of model parameters based on ESS values and examination of tree files. All newly generated sequences were deposited in GenBank with accession numbers MW025265-MW025275 and MW041968 - MW042028; (see Appendix 2 for cytochrome b sequences, Appendix 3 for cytochrome c oxidase subunit I and Appendix 4 for nuclear sequences, including previously generated ingroup and outgroup sequences used in this study).
Climatic analysis. Species climatic envelopes have been used for establishing species boundaries in cryptic species (Florio et al. 2012). To evaluate climatic differences among species that are closely related to Myotis sp. n., we only included specimens used in phylogenetic analysis (n = 19). We extracted values for 19 climatic variables in WorldClim at 2.5 arc second resolution (Fick and Hijmans 2017). Prior to conduct PCA analysis, we carried out a Pearson´s correlation analysis to indicate presence of multicollinearity among climatic variables. Based on this analysis, we selected nine bioclimatic variables as follows: BIO1 = Annual Temperature, BIO2 = Mean Diurnal Range, BIO4 = Temperature seasonality, BIO5 = Max Temperature of the Warmest Month, BIO6 = Min Temperature of the Coldest Month seasonality, BIO12 = Annual Precipitation, BIO13 = Precipitation of the Wettest Month, BIO14 = Precipitation of the Driest Month and BIO15 = Precipitation seasonality.
We conducted a principal component analysis to examine the degree of climatic variation among species, then we ran a MANOVA using informative rotated PC´s, followed by a post-hoc multiple comparison approach using a Bonferroni corrected approach, to evaluate pairwise differences among species. Statistical analyses were performed in software R (R Core Team 2020).
Results
Phylogenetic analysis. The partial cytochrome b alignment contained 145 sequences with 1, 140 columns, 620 distinct patterns, 434 informative sites, 66 singleton sites and 640 constant sites. ModelFinder found that the Best-fit model was the Hasegawa-Kishono-Yano, with empirical base frequencies allowing for a proportion of invariable sites plus discrete Gamma model (HKY + F + I + G4), whilst our cytochrome c oxidase subunit I alignment contained 61 sequences with 657 columns, 216 distinct patterns, 188 informative sites, 22 singleton sites and 447 constant sites. ModelFinder found Hasegawa-Kishono-Yano, with empirical base frequencies plus discrete Gamma model (HKY + F + G4) as the Best-Fit model according to BIC.
Both maximum likelihood and Bayesian phylogenetic analysis of mitochondrial genes sequences (Figures 1 and 2) from specimens that morphologically match M. pilosatibialis str. sensuLa Val (1973), were recovered as part of a monophyletic clade containing specimens from Guatemala, El Salvador, and México. This clade was found well-supported in cytochrome b analyses, with negligible support found in the cytochrome c oxidase subunit I phylogenetic tree. A specimen from Guatemala was geographically closest to the type locality in Honduras, therefore, we followed La Val (1973) in referring to this clade as M. pilosatibialis str.
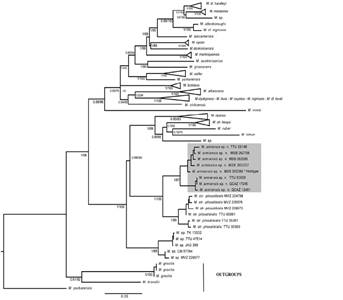
Figure 1 Partial cytochrome b phylogeny resulting from bayesian and maximum likelihood inference, with shaded grey sequences of M. armiensis sp. n., including holotype. The Bayesian analysis was conducted in MrBayes and maximum likelihood trees were generated using IQ-TREE with 100 bootstraps and 1000 replicates. Scores are bootstrap and probabilities values. Nodal support is shown right and left of slashes (“ /” ) respectively.
In our cytochrome b phylogenetic tree, a clade formed by sequences from specimens of M pilosatibialis str. is sister to a well-supported high elevation clade formed by specimens from Chiriquí and Cordillera Oriental (Ecuador), hereafter called M. sp. n., whilst our cytochrome c oxidase subunit I, analysis retrieved a supported clade from Costa Rica, Panamá and Ecuador samples as sister to M. sp. (Quintana Roo, Yucatán, México).
The phylogenetic placement of M. sp. (Quintana Roo, Yucatán, México) differs in our cytochrome b phylogenetic analysis, being paraphyletic to a well-supported monophyletic clade formed by M. sp. n. + M. pilosatibialis str. and the ruber group (simus, keaysi, ruber, riparius, sp.). M. sp. n. is part of well supported clade in both mitochondrial topologies. Within the ruber group, we were able to obtain a cytochrome b sequence from a specimen that morphologically matches M. keaysi sensuLa Val (1973) from Cochabamba, Bolivia which is the specimen geographically nearest to the type locality of Minas at Puno, Perú.
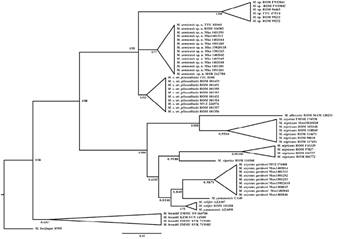
Figure 2 Partial cytochrome oxidase c subunit I phylogeny resulting from bayesian inference and maximum likelihood inference. The Bayesian analysis was conducted in MrBayes and maximum likelihood trees were generated using IQ-TREE with 100 bootstraps and 1000 replicates. Scores are bootstrap and probabilities values. Nodal support is shown right and left of slashes (“ /” ) respectively.
M. sp. n. is genetically distinct using both mitochondrial markers, the genetic pairwise distances between M. sp. n. and other species studied range from 6.1 to 12 %, p-distance 0.06 - 0.11 in partial cytochrome b, and from 3.6 to 9.8 %, p-distance 0.03 - 0.08 in partial cytochrome c oxidase subunit I. In partial cytochrome b, we retrieved the lowest value for conspecific populations from Chiriquí and eastern Cordillera Oriental (Ecuador; 3.1 %, p-distance 0.03). For cytochrome oxidase c oxidase subunit I, the lowest value was retrieved between conspecific populations from Valle del Silencio (Costa Rica) and Chiriquí Province (Panamá; 0.0 %, p-distance 0.00). Distance values are of the same order of magnitude as other interspecific and intraspecific comparisons within the taxa compared (Table 1).
Our species tree analysis readily identified M. sp. n. as genetically distinct from other species in the Neotropical and Nearctic Myotis radiation. Although we recovered relatively low support values for the newly identified clade, this topology also depicts a sister species relationship between conspecific populations from Chiriquí (Panamá) and Cordillera Oriental (Ecuador) as previously shown in our mitochondrial phylogenetic analyses (Figure 3). Results from two STACEY replicates analyses using nuclear data set suggested 20 putative species, under two collapse heights (0.0001 and 0.0005). Among the delimitated species, we found M. sp. n. as a candidate to be evaluated under an integrative taxonomy approach using independent data.
Table 1 Matrix of genetic distances (partial cytochrome b and cytochrome c oxidase subunit I) within and among three species of Myotis, where armiensis sp. n. in divided in two clades in cytochrome b analysis and three clades in cytochrome c oxidase subunit I. Below the diagonal:pairwise genetic distance using Kimura 2-parameter model (percentage). On the diagonal within clade distance using the Kimura 2-parameter model (percentage). Above the diagonal:pairwise p-distance values. Number of specimens sequenced in parenthesis.
| Partial Cytochrome-b | |||||
|---|---|---|---|---|---|
| Species/clades | 1 | 2 | 3 | 4 | 5 |
| 1 M. armiensis sp. n. Panamá (5) | 1 | 0.03 | 0.06 | 0.11 | 0.09 |
| 2 M. armiensis sp. n. Cordillera Oriental (Ecuador) (3) | 3.1 | 0 | 0.06 | 0.11 | 0.09 |
| 3 M. pilosatibialis (6) | 6.2 | 6.1 | 2 | 0.11 | 0.11 |
| 4 M. keaysi (1) | 12 | 11.7 | 11.7 | n.a | 0.11 |
| 5 M. sp. (5) | 9.8 | 10.1 | 10 | 12.1 | 2 |
| Partial Cyto-chrome c oxidase subunit I | |||||
| Species/clades | 1 | 2 | 3 | 4 | 5 |
| 1 M. armiensis sp. n. Panamá (2) | 0 | 0.01 | 0.01 | 0.03 | 0.08 |
| 2 M. armiensis sp. n. Cordillera Oriental (Ecuador) (1) | 1.8 | n.a | 0.01 | 0.03 | 0.08 |
| 3 M. armiensis sp. n. Costa Rica (11) | 0 | 1.8 | 0 | 0.03 | 0.08 |
| 4 M. pilosatibialis (6) | 3.6 | 3.8 | 3.6 | 0 | 0.07 |
| 5 M. sp. (5) | 9.5 | 8.7 | 9.8 | 8.1 | 1.3 |
Morphological analysis. The principal component analysis comparing all five species extracted two major components that accounted for 53.0 and 30.3 % of the variation (Figure 4; Table 2). The PCA plot shows that M. sp. n. overlaps partially with M. pilosatibialis str., which is due to size and shape similarities, while all of the other species were distinct. M. str. keaysi and M. oxyotus gardneri plotted at the lower left end of PC1 reflecting their larger size, whilst M. sp. (Quintana Roo, Yucatán, México) plotted at the right lower end, which is explained by its small size. PC1 shows variation in size and is influenced most by variables related to length of the skull, having MAL, GLS, CCL, CBL, CIL and M1-M3 with loadings above 0.8. PC2 shows shape variation, and it is most influenced by measurements associated with the width of the skull, with BCB and POB having loadings above 0.8.
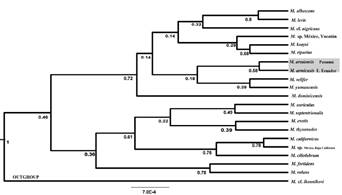
Figure 3 Species tree inferred in *BEAST using multilocus sequence data for New World Myotis. Number under branches represent bayesian posterior probability values with conspecific populations from Ecuador and Panama shaded grey.
A one-way multivariate analysis of variance suggested there was a statistically significant difference between all species on both PC1 and PC2, as suggested by Pillai’s Trace test, F(8,58) = 15.411, P < 0.001. Follow-up ANOVAs showed that this dissimilarity occurred at both PC´s (PC1, F(4,29) = 15.9, P < 0.05, PC2, F(4,29) = 15.0, P < 0.05). Post-hoc multiple comparison analyses revealed that M. sp. n. is different from M. oxyotus gardneri (P < 0.01), and M. str. keaysi (P < 0.05) in PC1, and different from M. sp.(Quintana Roo, Yucatán, México; P < 0.001) and M. str. pilosatibialis (P < 0.05) in PC2. For other comparison, see Appendix 5.
Table 2 Factor loading after PCA and varimax rotation, with Kaiser Normalization for two principal components from a principal component analysis (PCA). The analysis is based on 15 craniometric measurements of five species, including 33 individuals. See Methods for variable abbreviations.
| Measurements | PC I | PC II |
|---|---|---|
| MAL | -.939 | .231 |
| MAN | -.695 | .585 |
| GLS | -.914 | .318 |
| CCL | -.862 | .444 |
| CBL | -.866 | .423 |
| CIL | -.944 | .245 |
| BAL | -.754 | .574 |
| MAB | -.751 | .323 |
| BCB | ---- | .921 |
| IOB | -.645 | .564 |
| POB | -.374 | .812 |
| BAC | -.337 | .599 |
| BAM | -.513 | .761 |
| MTL | -.765 | .575 |
| M1-M3 | -.840 | .299 |
| eigenvalues | 11.14 | 1.34 |
| % of variance explained | 53.0 | 30.3 |
| cumulative % | 53.0 | 83.3 |
Although M. sp. n. is overlapping partially with M. pilosatibialis, our DFA analysis readily distinguishes M. sp. n. from other species along the first axis (83.5 %) and to a lesser extent the second axis (9.9 %, Figure 5; Tabla 3). The jackknifed classification matrix showed that the analysis correctly classified 81.8 % of the specimens, with M. sp. n. 70 % correctly classified, M. str. pilosatibialis 75 % correctly classified, M. sp. (Quintana Roo, Yucatán, México) 100 % correctly classified, M. str. keaysi 75 % correctly classified, and M. oxyotus gardneri 100 % classified.
Table 3 Vector coefficient correlations between original variables and discriminant functions (DF1 and DF2) for selected samples of Myotis.
| Measurements | DF1 (83.5%) | DF2 (9.9%) |
|---|---|---|
| MAL | -32.71 | -179.89 |
| MAN | 135.04 | 62.82 |
| GLS | 215.37 | -84.56 |
| CCL | -120.75 | -83.33 |
| CBL | 42.31 | -75.27 |
| CIL | -136.83 | -52.95 |
| BAL | 70.81 | -144.96 |
| MAB | 190.98 | 142.86 |
| BCB | 21.57 | 49.32 |
| IOB | .67 | -23.05 |
| POB | 18.80 | -36.78 |
| BAC | -97.68 | 106.43 |
| BAM | 38.63 | -76.04 |
| MTL | -89.95 | -23.80 |
| M1-M3 | 78.38 | 85.67 |
Cranial index of M. sp. n. (CRI: 50.6) is larger than M. str. pilosatibialis (CRI: 47.3), M. sp. (Quintana, Roo, Yucatán, México, CRI: 42.1), but smaller than M. str. keaysi (CRI: 51.5) and M. oxyotus gardneri (CRI: 53.8).
Climatic analysis. Our PCA for climatic variables resulted in three PCA´s with eigenvalues > 1, explaining 85.6 % of the total variation. PCA1 was primarily informative for Min Temperature of the Coldest Month, whilst Mean Diurnal range was the principal driver along PCA2. In contrast Annual Temperature was the principal driver of PC3. One-way multivariate analysis of variance suggested there was a statistically significant difference on PC1, PC2, and PC3 as suggested by Pillai’s Trace test, F(6, 30) = 2.283, P < 0.1. Follow-up ANOVAs showed that this dissimilarity occurred at PC3 F(2, 16) = 4.32, P < 0.5, but not for PC1 F(2, 16) = 1.50 ns and PC2 F(2, 16 ) = 1.50 ns. Post-hoc multiple comparison analyses did not reveal statistically significant differences in the climatic envelopes of all species, as indicated by extensive overlap (Figure 6; Table 4).

Figure 4 Principal components (PC´s) from a PCA based on 15 cranial measurements from 33 individuals. Samples:M. armiensis sp. n (circles), M. sp. (triangles), M. oxyotus gardneri ( + symbols), M. keaysi str. ( X symbols), and M. pilosatibialis str. (diamonds).

Figure 5 Vector correlation coefficients (loadings) between original variables and discriminant functions (DF1, DF2), with jackknifed percentage of correctly classified specimens for each group. Samples: M. armiensis sp. n (circles), M. sp (triangles), M. oxyotus gardneri ( + symbol), M. keaysi str. ( x symbol), and M. pilosatibialis str.(diamonds).
Discussion
In this report we were able to determine the phylogenetic placement of M. pilosatibialis str. based on specimens from Alta Verapaz, Guatemala (San Pedro Carcha, Finca Bethel, 15° 0.61 ′ N, -90° 0.27 ′ W), the collecting locality for specimens included in the subspecies description (La Val 1973). This locality is ca. 370 km from the type locality at Francisco Morazán, Honduras (14° 0.24 ′ N, -85° 0.5 ′ W), therefore we recommend assigning the name pilosatibialis to this species complex, which has a disjunct distribution (primarily low and high elevation species) from southern México, southeastward through Central America into northwestern Panamá, and to the eastern cordillera of the Colombian and Cordillera Oriental (Ecuador). Elsewhere, this complex occurs from the island of Trinidad, to Venezuela and the Colombian Caribbean. This group is characterized by small to medium size, with short woolly hair, and the dorsal surface of tibia partially or entirely covered by fur, which might extend to the hindfoot and across the tibia and onto the plagiopatagium. A flattened occipital region and moderate to high sagittal crest characterize the skull. In agreement with Moratelli et al. (2016) and Moratelli et al. (2017), we included pilosatibialis as part of the ruber group based on shared morphological traits (woolly hair, moderate to high sagittal crest, and flat occipital crest) and close molecular evolutionary proximity. Likewise, we determined the phylogenetic placement of M. keaysi str., from voucher specimens collected in Cochabamba, Bolivia (-17° 0.21 ′ S, -65° 0.6 ′ W), located ~ 600 km distant from the type locality of M. keaysi at Inca Mines, Cuzco, Perú (-13° 0.30 ′ S ,-70 ° 0.0 W). Myotis keaysi is another member of the ruber group, with larger external and cranial dimensions, and longer woolly fur, now suggested to be distributed in high elevation sites from the Andes of Colombia, southward to Bolivia and Argentina (Moratelli et al. 2013). According to Mantilla-Meluck and Muñoz-Garay (2014) both species are living in sympatry in the Colombian Caribbean and eastern cordillera of the Andes of Colombia.
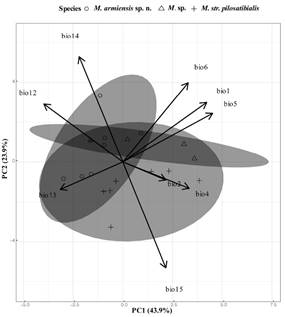
Figure 6 Principal component (PC´s) from a PCA based on 9 bioclimatic variables extracted from 19 distribution localities of pilosatibialis species complex, with confidence ellipses and corresponding vectors correlations of climatic variables with the first two eigenvectors. Samples:M. armiensis sp. n. (circles), M. sp. (triangles), and M. pilosatibialis str.(+ symbols).
Larsen et al.’s (2012 a, b) molecular study of Neotropical Myotis identified an unnamed clade formed by specimens misidentified as M. nigricans (TTU 39146 from Santa Clara, Chiriquí, Panamá) and M. riparius (TTU 85060 from Azuay, Tungurahua, Ecuador) collected by Robert J. Baker and collaborators in 1983 (Panamá) and 2001 (Sowell-Expedition, Ecuador). Chaverri et al. (2016) referred to this clade as M. keaysi based on cytochrome c oxidase subunit I sequences of unvouchered specimens from Valle del Silencio, Costa Rica and one specimen (ROM 104302), collected by the Royal Ontario Museum in Santa Clara, Ojo de Agua, 2 km N of Santa Clara, Panamá.
Table 4 Factor loading after PCA and varimax rotation, with Kaiser Normalization for three principal components from a principal component analysis (PCA). The analysis is based on 9 bioclimatic variables, including 19 localities.
| Climatic variable | PC I | PC II | PCIII |
|---|---|---|---|
| Annual Mean Temperature | .972 | .209 | .086 |
| Mean Diurnal Range | -.013 | .697 | .027 |
| Temperature Seasonality | .242 | .633 | .287 |
| Max Temperature of the Warmest Month | .923 | .337 | .128 |
| Min Temperature of the Coldest Month | .988 | -.037 | -.036 |
| Annual Precipitation | -.157 | -.820 | -.480 |
| Precipitation of the Wettest Month | -.241 | -.883 | .243 |
| Precipitation of the Driest Month | -.048 | -.086 | -.965 |
| Precipitation Seasonality | .047 | .073 | .973 |
| eigenvalues | 2.9 | 2.5 | 2.2 |
| % of variance explained | 32.4 | 27.8 | 25.3 |
| cumulative % | 32.4 | 60.2 | 85.6 |
Prior to those studies, voucher specimen series of Myotis collected from the highlands of Chiriquí Province (USNM, TCWC) had been identified as M. k. pilosatibialis. Our bat surveys (in Ecuador in 2017 and Panamá in 2012) provided additional specimens for morphological and genetic studies. Herein, we have examined specimens assigned to the unnamed clade sensuLarsen et al. (2012 a, b), and found morphometric and genetic differences that support recognition of a new species. We propose that populations from Chiriquí Province (Panamá) and Cordillera Oriental (Ecuador) represent a new species, which is described as follows:
Myotis armiensis, species novum
Armien’s Myotis, Myotis de Armién
Figure 7, 8, and 9; Tables 5 and 6
Myotis keaysi pilosatibialis, La Val, 1973, part
Myotis keaysi pilosatibialis, Hernandez-Meza et al. 2005, part.
Myotis keaysi pilosatibialis, Wilson, 2008, part.
Myotis nigricans, Larsen et al. 2012a, part.
Myotis riparius, Larsen et al. 2012b, part.
Myotis keaysi, Chaverri et al. 2016, part.
Myotis cf. pilosatibialis, Moratelli et al. 2016, part.
Myotis cf. pilosatibialis, Moratelli et al. 2017, part.
Holotype and type locality. Voucher MSB 262089; adult male; preserved as skin, skull and skeleton (Figures 8 and 9) at the Museum of Southwestern Biology (MSB), University of New Mexico collected on 20 March 2012 by Joseph A. Cook and collaborators (Tropical Biology Class 2012 and Instituto Conmemorativo Gorgas de Estudios de la Salud) at La Amistad International Park Ranger Station (8° 89 ′ N, -82° 61 ′ W, elevation 2, 214 m), Bugaba District, Chiriquí Province, Panamá. Tissues deposited at the same museum (NK 209314, LN2 preservation), with additional tissues at Instituto Gorgas, Panama City (-80°C preservation). The specimen is well preserved. The measurement for the forearm (44 mm) recorded on the specimen tag is incorrect and should be FA = 39 mm as now recorded in the Arctos Museum Database.

Paratypes. Nineteen additional specimens were collected from Chiriquí Province, Panamá and Cordillera Oriental (Ecuador). Two specimens were designated as paratypes based on genetic identification, (partial cytochrome b), morphometrics and qualitative data: skin of one adult female (MSB 262085), collected by Joseph A. Cook and collaborators, 20 March 2012; skin and skull of one female, adult (TTU 85060) collected by Robert J. Baker and collaborators (Sowell Expedition-Ecuador, 2001), 24 July 2001.
Six paratypes identified solely by genetic identification (partial cytochrome b) were collected at Bugaba District, Chiriquí Province, Panamá ;one adult male (MSB 262237) collected by Joseph A. Cook and collaborators, 20 March 2012), Jurutungo, Río Sereno, Renacimiento District, Chiriquí Province, Panamá ;sex unknown (MSB 262788), collected by Gorgas Institute field researchers, 6 May 2011, Santa Clara, Renacimiento District, Chiriquí Province, Panamá ;one female, age unknown (TTU 39146), collected by Robert J. Baker, 19 January 1983, Cabañas del Aliso, Cosanga, Napo Province, Ecuador ;one female, age adult, (QCAZ 17245) collected by Carlos A. Carrión Bonilla, 13 December 2017), Yantzaza, Campo Minero Fruta del Norte, Zamora Chinchipe, Ecuador ;male, adult (QCAZ 12461), collected by Paula Iturralde, 3 March 2011. Another specimen from Ojo de Agua, 2 km N of Santa Clara, Renacimiento District, Chiriquí Province, Panamá (ROM 104302), collected by Burton Lim and Eamon O’Toole, on 8 March 1995 was identified based on barcode cytochrome c oxidase subunit I.
Table 6 Selected measurements, cranial index, body mass (adults males and females;N). Mean, (Minimum-Maxima) and sample size (n). See methods for variable abbreviations.
| Characters | M. armiensis sp. n. Chiriquí (Panamá) and Cordillera Oriental (Ecuador) | M. keaysi str. Perú | M. pilosatibialis str. El Salvador Honduras México | M. sp. Quintana Roo, Yucatán, México | M. oxyotus gardneri Costa Rica and Panamá |
|---|---|---|---|---|---|
| Body mass | 5.0 (4.5-5.6) n = 10 | 6.0 n = 1 | 6.1 n = 1 | 8 n = 1 | |
| HB | 86 (77-92) n = 12 | 94.0 n = 1 | 93.0 (87-98) n = 3 | ||
| Tail | 39 (32-48) n = 13 | 42.0 n = 1 | 40.3 (39-43) n = 3 | ||
| Foot | 8 (7-10) n = 13 | 8.0 n = 1 | 8.3 (8-9) n = 3 | ||
| Ear | 13 (11-14) n = 13 | 14.0 n = 1 | 15.7 (15-16) n = 3 | ||
| LDF | 6.2 (5.3-7.4) n = 10 | 7.7 (7.4-7.9) n = 3 | |||
| LVF | 6.3 (4.5-8.3) n = 10 | 6.6 (5.6-7.2) n = 3 | |||
| FA | 38.0 (36.3-39.4) n = 13 | 40.2 n = 1 | 35.9 (34.3-37.1) n = 5 | 32.6 (31.8-34.4) n = 4 | 40.4 (38.0-42.8) n = 6 |
| 3MC | 34.3 (32.9-35.9) n = 13 | 36.8 n = 1 | 33.2 (32.1-34.4) n = 5 | 29.3 (27.7-30.4) n = 4 | 36.3 (34.5-38.6) n = 6 |
| GLS | 13.5 (13.0-14.0) n = 10 | 14.5 (14.1-15.0) n = 5 | 13.6 (13.5-13.9) n = 8 | 12.7 (12.6-13.0) n = 4 | 14.5 (14.1-15.0) n = 6 |
| CCL | 12.1 (11.7-12.5) n = 10 | 12.6 (12.0-13.0) n= 5 | 12.1 (11.8-12.3) n = 8 | 11.4 (11.2-11.7) n = 4 | 12.8 (12.3-13.3) n = 6 |
| CBL | 12.7 (12.3-13.0) n = 10 | 13.3 (12.6-13.8) n = 5 | 12.7 (12.4-12.9) n = 8 | 12.1 (11.7-12.4) n = 4 | 13.5 (13.1-14.1) n = 6 |
| CIL | 12.7 (11.9-13.2) n = 10 | 13.5 (12.9 -14.0) n = 5 | 12.9 (12.7-13.1) n = 8 | 12.1 (11.8-12.5) n = 4 | 13.7 (13.2-14.2) n = 6 |
| BAL | 11.7 (11.3-12.1) n = 10 | 12.1 (11.5-12.5) n = 5 | 11.6 (11.3-11.9) n = 8 | 11.0 (10.7-11.3) n = 4 | 12.4 (12.0-12.8) n = 6 |
| ZB | 8.5 (7.9-8.8) n = 8 | 8.6 (8.5-8.8) n = 4 | 8.2 (8.1-8.4) n = 6 | 8.9 n = 1 | |
| MAB | 7.0 (6.6-7.4) n = 10 | 7.0 (6.9-7.2) n = 5 | 7.0 (6.9-7.2) n =8 | 6.7 (6.6-6.7) n = 4 | 7.3 (7.1-7.4) n = 6 |
| BCB | 6.8 (6.2-7.3) n = 10 | 6.5 (6.4 -6.5) n = 5 | 6.3 (6.1-6.5) n =8 | 5.9 (5.8-6.0) n = 4 | 6.8 (6.6-7.0) n = 6 |
| IOB | 4.5 (4.2-4.8) n = 10 | 4.7 (4.5-4.9) n =5 | 4.3 (4.2-4.5) n = 8 | 4.0 (3.9-4.1) n = 4 | 4.7 (4.5-4.9) n = 6 |
| POB | 3.6 (3.2-4.2) n = 10 | 3.6 (3.4-3.8) n = 4 | 3.4 (3.3-3.5) n = 8 | 3.1(3.1-3.2) n = 4 | 3.9 (3.8-4-0) n = 6 |
| BAC | 3.6 (3.4-4.0) n = 10 | 3.6 (3.5-3.7) n = 5 | 3.6 (3.5-3.8) n = 8 | 3.3 (3.2-3.4) n = 4 | 3.6 (3.4-3.8) n = 6 |
| BAM | 5.5 (5.3-6.0) n = 10 | 5.6 (5.4-5.8) n = 5 | 5.3 (5.2-5.5) n = 8 | 4.9 (4.7-5.0) n = 4 | 5.7 (5.5-5.8) n = 6 |
| MTL | 5.2( 5.1-5.4) n = 10 | 5.5 (5.2-5.6) n = 5 | 5.1(5.0-5.2) n = 8 | 4.8(4.6-4.9) n = 4 | 5.5 (5.4-5.6) n = 6 |
| M1-3 | 2.9 (2.5-3.0) n = 10 | 3.1 (3.0-3.3) n =5 | 2.9 (2.8-3.0) n = 8 | 2.6 (2.5-2.6) n = 4 | 3.1 (3.1-3.2) n = 6 |
| MAL | 9.6 (8.9-10.3) n = 10 | 10.2 (9.9-10.5) n =5 | 9.6 (9.5-9.7) n = 8 | 9.1 (8.8-9.3) n = 4 | 10.4 (10.0-10.9) n = 6 |
| MAN | 5.5 (5.3-5.7) n = 10 | 5.8 (5.6-5.9) n = 5 | 5.5 (5.4-5.7) n =8 | 5.1 (4.9-5.2) n = 4 | 5.7 (5.7-5.9) n = 6 |
| CRI | 50.6 (47.0-53.7) n = 10 | 51.5 (49.7-53.9) n = 5 | 47.3 (45.8-49.1) n = 8 | 42.1 (41.1-43.3) n = 4 | 53.8 (50.7-56.0) n = 6 |
| MAX | 18.1 (17.2-19.6) n = 10 | 18.8 (17.6-19.9) n = 5 | 17.3 (16.7-17.9) n = 8 | 15.0 (14.0-15.5) n = 4 | 19.2 (18.4-19.8) n = 6 |
Ten paratypes identified with morphometric analysis of craniometric measurement analysis and qualitative morphological of skins and skulls were collected from La Amistad International Park Ranger Station, Bugaba District, Chiriquí Province, Panamá ;(two females, adults (MSB 262217-18, only skins were studied, no skulls available) collected by Joseph A. Cook and collaborators, 20 March 2012); Cerro Punta, Casa Tiley, Tierras Altas District, Chiriquí Province, Panamá ;female, adult (USNM 323599, by Handley, C. and Greenwell, F.M., 6 March, 1962); El Volcán 2 min S. W, Tierras Altas District, Chiriquí Province, Panamá ;two females, adults, (USNM 331942, USNM 331943, by Tyson E, collected 21 March 1962); Cuesta de Piedra, Tierras Altas District Chiriquí Province, Panamá ;female, adult (USNM 331953, by Tyson E, 28 March 1962); 36 km, north of Concepción, Bugaba District, Chiriquí Province, Panamá ;five females, adults (TCWC 12655-59, by Patten, D.R., 8 June 1964). We did not include any specimens from Costa Rica in the type series, because the Chaverri et al. (2016) study was unvouchered, and therefore morphological confirmation is not possible.
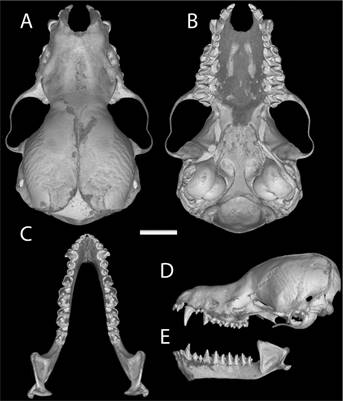
Figure 8 Dorsal (A) and ventral view (B) of the cranium, dorsal view of the mandible (C), and lateral view of the cranium (D) and mandible of the holotype of M. armiensis sp. n. (MSB 262089). Scale bar = 5 mm. Photograph taken by John Korbin (Sandia National Laboratory-New Mexico, USA).
Distribution. Myotis armiensis sp. n. is known from the premontane and montane forest of Chiriquí Province, Panamá, extending its distribution into La Amistad International Park in Panamá (Las Nubes Rangers Station) and Costa Rica (Valle del Silencio). Elevation in Panamá and Costa Rica varies from 975 m to 2,500 m: Concepción ( ≈ 8°.51 ′ N, -82°.62 ′ W, 2,011m), Cerro Punta (8°.88 ′ N, -82°.73 ′ W, 1,280 m ), Cuesta de la Piedra (8°.88 ′ N, -82°.73 ′ W, 975 m), El Volcán (8°.88 ′ N, -82°.73 ′ W, 1,280 m), Parque Internacional la Amistad Ranger Station: (8°.89 ′ N, -82°.61 W, 2, 214 m), Jurutungo, Río Sereno (8°.9 ′ N, -82°.73 W, 2,219 m), Santa Clara ( ≈ 8°.83 ′ N, -82°.75 ′ W, ≈ 1,178 m), Ojo de Agua, Santa Clara (8° 42 ′ N, -82°.45 W, 1,500 m), Valle del Silencio, Costa Rica (9° 11 ′ N, -82°.96 W, 2,500 m). In Ecuador, M. armiensis sp. n. is known from Cordillera Oriental. Elevation in Ecuador varies from 1,200 m to 2,249 m: Yantzaza, Campo Minero Fruta del Norte, Zamora Chinchipe (-3°.75 ′ S, -78°.53 ′ W, 1, 200 m), Colonia Azuay, Tungurahua Province, Ecuador (-1°.34 ′ S, -78.20°, 1,660 m ), Cabañas el Aliso, Sector Las Caucheras, Quijos, Napo Province (-3°.75 ′ S, -78°.53 ′ W, 2,249 m). See Figure 10 for species distribution in Costa Rica, Panamá and Ecuador.
Table 5 Selected measurements (mm) and body weight (g) of the holotype (MSB 262089) and paratype material of Myotis armiensis. See methods for variable abbreviations.
| Characters | MSB 262089 ♂ | Paratypes |
|---|---|---|
| Body mass | 5 | 5 (4.5-5.6) n = 9 |
| HB | 90 | 85 (77-92) n = 11 |
| Tail | 38 | 39 (32-48) n = 12 |
| Foot | 8 | 8 (7-10) n = 12 |
| Ear | 13 | 14 (11-14) n = 12 |
| LDF | 6.4 | 6.1 (5.3-7.4) n = 9 |
| LVF | 7.2 | 6.2 (4.5-7.4) n = 9 |
| FA | 37.1 | 38.1 (36.3-39.4) n = 12 |
| 3MC | 34 | 34.3 (32.9-35.9) n = 12 |
| GLS | 13.3 | 13.5 (13.0-14.0) n = 9 |
| CCL | 12.2 | 12.1 (11.7-12.5) n = 9 |
| CBL | 12.3 | 12.8 (12.5-13.0) n = 9 |
| CIL | 11.9 | 12.8 (12.2-13.2) n = 9 |
| BAL | 12.1 | 11.6 (11.3-11.9) n = 9 |
| ZB | 7.9 | 8.6 (8.0-8.8) n = 7 |
| MAB | 6.6 | 7.1 (6.6-7.4) n = 9 |
| BCB | 7.3 | 6.8 (6.2-7.2) n = 9 |
| IOB | 4.4 | 4.6 (4.2-4.8) n = 9 |
| POB | 4.2 | 3.5 (3.2-3.7) n = 9 |
| BAC | 3.6 | 3.7 (3.4-4.0) n = 9 |
| BAM | 5.4 | 5.6 (5.3-6.0) n = 9 |
| MTL | 5.2 | 5.2 (5.0-5.4) n = 9 |
| M1-3 | 2.7 | 2.9 (2.5-3.0) n = 9 |
| MAL | 9.5 | 9.6 (8.9-10.3) n = 9 |
| MAN | 5.7 | 5.5 (5.3-5.7) n = 9 |
Etimology. Myotis armiensis honors Dr. Blas Armién in recognition of his outstanding contributions to research in zoonotic emergent diseases, public health, and mammalogy in Panamá. Over two decades, he has supported the systematic development of holistic museum collections, including associated cryogenic biorepositories and parasites. This infrastructure (>11,000 specimens) is now the basis for new insights into temporal and spatial aspects of the biology of Panama’s mammals and associated parasites and pathogens.
Diagnosis. Due to the dearth of availability of substantial specimen material and despite the lack of a single morphological character that consistently distinguishes M. armiensis from the rest of Neotropical congeners, Myotis armiensis can be distinguished by a combination of the following morphological traits: pelage is short and woolly, dorsal and ventral fur bicolored; insertion of the plagiopatagium occurs on the foot at the level of the base of toes by a wide membrane; lack or relatively low presence of fur on dorsal surface of tibia, foot and plagiopatagium, and border of the uropatagium without a fringe; skull is moderately large; forehead is steeply sloping; rostrum is long; lambdoidal crest is present and high; sagittal crest present, with height from low to medium; occipital crest is absent; occipital region flattened and the shape of the braincase is globular.
Myotis armiensis can be readily distinguished from Myotis congeners from Central and South America based on gene trees; partial sequence of cytochrome-b ( ~710 pb) and partial cytochrome c oxidase subunit I ( ~ 657 pb) and species tree phylogenetic analysis of one exon: recombination activating gene II (RAG2), and 3 intron regions: protein kinase C, iota (PRKCI), signal transducer and activator of transcription 5A (STAT5A), and thyrotropin (THY). Finally, molecular synapomorphies in mitochondrial and intron genes regions sequences support the species level recognition of M. armiensis sp. n. compared to other species in the New World Myotis radiation (Appendix 6 - 10 ). These served as diagnostic characters for the species.
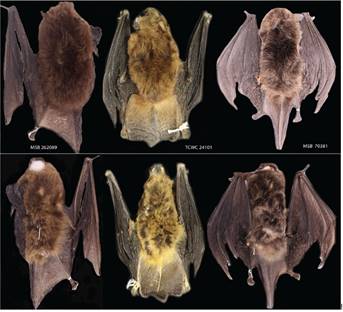
Figure 9 Dorsal and ventral views of the skins of the holotype of M. armiensis sp. n. (MSB 262089), M. pilosatibialis str.(TCWC 24101, paratype), and M. keaysi str. (MSB 70381). Scale bar = 10 mm.
Description. A medium to large species of Myotis (FA 36.3-39.4 mm, n = 13 and weight 4.5-5.6 gr, n = 10); other measurements (Tables 5 and 6), with external size larger than M. pilosatibialis str., M. sp. (Quintana Roo, México) and smaller than M. oxyotus gardneri, and M. keaysi str. Ears are brown in color, comparatively small to medium-sized (EL 11-14 mm). Dorsal and ventral fur is woolly and short (LDF 5.3 -7.4 mm, LVH 4.5-8.3 mm). Dorsal pelage is bicolor, with brown to dark brown at the base and from brown to Mummy Brown at the tips. Ventral pelage is bicolor, with Buckthorn Brown to buff at the tips and dark brown to black at the base. Abdomen is bicolor, from Buckthorn Brown to black with buff tips. Sides and wing color are dark brown or Cinnamon Brown to Mummy Brown. Uropatagium and plagiopatagium are Mummy Brown or Cinnamon Brown. Insertion of the plagiopatagium occurs at the foot at the level of the base of toes by a wide membrane. The uropatagium lacks fringing hairs along the trailing edge. Fur presence on tibia, foot and plagiopatagium, with fur extending across a quarter or less than this at the base of the dorsal and ventral side of uropatagium. Skull and mandible are medium-size (GLS 13.0 -14.0 mm, MAL 8.3-10.3). The dental formula is: 2/3, 1/1, 3/3, 3/3 = 38. In the holotype, the second upper premolar (p3) is aligned and visible in lateral view. Rostrum is long and frontals are steeply sloping; lambdoidal crest is well developed and occipital region is flattened.
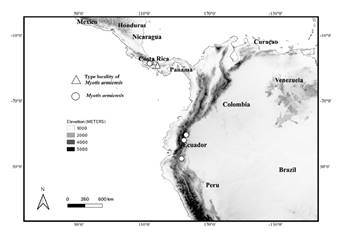
Figure 10 Map of part of Central and South America showing localities examined for M. armiensis, with triangle (type locality) and distribution in Costa Rica, Panamá, and Ecuador with circles. See Appendix I for localities of examined in Panamá and Ecuador.
Comparisons. In comparison with species in the ruber group, M. armiensis sp. n. differs from simus and riparius by having a less contrasting dorsal and ventral coloration (dorsal fur with dark brown to Mummy Brown and ventral fur with Buckthorn Brown to dark brown), with more contrasting (Orange-Brown or Chocolate) in simus, (golden -yellow) in midastactus, (ventral hairs with dark brown based and yellowish tips/reddish-brown or cinnamon brown dorsal color) in riparius, (Bister Brown color of dorsal hairs at tips and buff to orange of ventral color at tips) in pilosatibialis ;type material. M. armiensis sp. n. can be distinguished from keaysi by having shorter woolly hair on dorsal and ventral side, with longer woolly hair, larger proportion of fur on dorsal and ventral side of the uropatagium, and furrier tibia in keaysi. It differs from simus by having the plagiopatagium attached broadly to the side of the foot at the level of the toes; with a narrow band of membrane (< 1.5 mm) attached to foot or ankles and with extremely short and woolly fur in simus. M. armiensis sp. n. shares the flattened occipital region and a moderate to high sagittal crest with members of the ruber group. It can be distinguished from M. ruber by presence of woolly hair, with silky pelage in ruber. Cranial index of M. armiensis sp. n. is only larger that M. pilosatibialis str. but smaller than other congeners in the ruber group (CRI: armiensis = 47.0-53.7 , pilosatibialis = 45.8-49.1, keaysi = 49.7-53.9; simus = 73.0-88.1; ruber = 78.4-85.0; riparius = 64.6-76.1), reflecting a narrower skull configuration.
Myotis armiensis sp. n. differs from the albescens group (albescens, handleyi, nesopolus, nigricans, oxyotus oxyotus, oxyotus gardneri, izecksohni, lavali and levis) by having short woolly hair. It only shares the woolly hair trait with chiloensis. In addition, it can be distinguished from albescens group by the presence of a moderate sagittal crest ;absent in albescens, very low or absent in oxyotus, absent in nigricans or very low, absent in lavali, very low in chiloensis. Myotis armiensis sp. n. differs from levis and albescens by the absence of fringe of hairs along the edge of the uropatagium. Occipital region is flattened in M. armiensis sp. n. ;rounded in albescens, oxyotus, lavali, nigricans. The cranial index for albescens group is larger than M. armiensis sp. n, having the narrowest skull configuration in comparison with this species group.
Reproductive data. One pregnant female with one embryo (MSB 262085) collected by Joseph A. Cook and collaborators at Las Nubes Ranger Station, La Amistad International Park, 20 March 2012. Another female (USNM 323599) with embryo (crown-rump = 3 mm) collected at Cerro Punta, Casa Tiley, Chiriquí Province, collected by Greenwell, F. M, 6 March 1962. In Ecuador, one pregnant female (TTU 85060) with embryo (crown-rump = 3 mm) collected by Robert J. Baker and collaborators (Sowell-Expedition, 2001) at Colonia Azuay, Tungurahua Province, 24 July, 2001.
Habitat and ecological notes. Las Nubes Ranger Station, the type locality for this new species, is part of a natural corridor (401, 000 ha) of relatively undisturbed montane habitats of the eastern Talamanca Mountains that rise between the Pacific and Caribbean coastlines of Panamá and Costa Rica (Morrone 2017). At La Amistad International Park, Las Nubes Ranger Station, M. armiensis sp. n. was found to occur sympatrically with Desmodus rotundus, Anoura geoffroyi, Sturnira cf. burtonlimi, Sturnira mordax, Enchistenes hartii (Phyllostomidae), and Tadarida brasiliensis (Molossidae). All captures in 2011, 2012 were with ground mist nets located on the edge of secondary growth forest near Las Nubes Rangers Station. Specimens were collected at a clearing on the edge of the forest (Cerro Punta, Casa Tiley) and in montane forest near a lake shore and moist montane secondary growth forest (2 miles S. W of El Volcán). We did not record any other species of Myotis living in sympatry at La Amistad International Park, but based on historic samples, the nominal subspecies M. oxyotus gardneri was found in sympatry with the new species at Cerro Punta and El Volcán.
At Valle del Silencio Costa Rica, M. armiensis sp. n. was found with Sturnira burtonlimi, Dermanura tolteca, Hylonycteris underwoodi, Anoura cultrata (Phyllostomidae), and Lasiurus blossevillii, Myotis cf. nigricans, and M. oxyotus gardneri (Vespertilionidae). The vegetation there was characterized by the presence of oak trees (Quercus spp) and bamboo (Chusquea spp.), although some sites at Valle del Silencio were dominated by swampy bogs (Chaverri et al. 2016).
In Ecuador, M. armiensis sp. n. is known from premontane and montane cloud forest of Cordillera Oriental. These forests correspond to Evergreen Lower Montane Forest and Cloud Montane Forest (Bosque Siempreverde Montano Bajo and Bosque de Neblina Montano; Valencia et al. 1999). One specimen of M. armiensis sp. n. was netted across a trail leading to secondary growth forest in Cabañas del Aliso, Cosanga, Quijos Valle, Napo Province in December, 2017. Other species captured in sympatry at that location were: Sturnira bogotensis, Carollia brevicauda (Phyllostomidae), Tadarida brasiliensis (Molossidae), and M. oxyotus oxyotus and Histiotus montanus (Vespertilionidae). The vegetation there was characterized by the presence of epiphytes (moss, ferns, orchids, bromeliads) , bamboo (Chusquea spp.), and pepper plants (Piper spp. and Peperomia spp.). Another specimen of M. armiensis sp. n. was captured in Colonia Azuay, Tungurahua Province at the north side of the Río Pastaza. This locality is bisected by the Río Topo, a tributary of the Pastaza river and comprises secondary forest and fruit orchards (Haynie et al. 2006). Other bat species captured in sympatry at that location were: Anoura caudifer, Artibeus lituratus, Carollia perspicillata, and Sturnira erythromus. (Phyllostomidae).
Remarks. It was not possible to examine a set of fluid preserved specimens (n = 43) collected at La Amistad International Park Ranger Station, Bugaba District, Chiriquí Province, collected in 2018 by Joseph A. Cook and collaborators (Tropical Biology Class 2018 and Gorgas Institute field workers). Those specimens, until recently housed at the Gorgas Institute in Panama, may represent M. armiensis sp. n., but confirmation will require further molecular and morphological analysis (MSB 262086 -88, MSB 262219-262224, 268090-93, MSB 327505-327516, MSB 327520, MSB 327522 -327527, MSB 327575-578, MSB 327601-602, MSB 327649-50, MSB 327700, MSB 327703, MSB 327705, MSB 327709, MSB 327712, MSB 327956).
Based on this report and other museum collections records, the diversity of Myotis comprises at least six species in Panamá: M. albescens, M. oxyotus gardneri, M. riparius, M. pilosatibialis, M. nigricans s.l., and M. armiensis sp. n. In Ecuador, the recognition of this newly identified lineage, increases the diversity to eight species of Myotis: M. albescens, M. riparius, M. simus, M. oxyotus oxyotus, M. diminutus, M. keaysi, M. nigricans s.l., and M. armiensis sp. n. In Costa Rica, Myotis diversity increases to seven species: M. elegans, M. riparius, M. albescens, M. pilosatibialis, M. nigricans s.l., M. oxyotus. gardneri and now M. armiensis sp. n.
Nomenclatural statement.- A life science identifier (LSID) number was obtained for new species described herein: urn:lsid:zoobank.org:pub:9EB39E62-C9AC-41C0-AE76-9C5325608BEE.
Conservation. Myotis armiensis sp. n. is restricted to higher and cooler mountain forest of Panamá, Costa Rica and Cordillera Oriental (Ecuador). These habitats are susceptible to the effects of climate change, in addition to ongoing habitat destruction. This report aims to contribute to efforts to study these environments and more narrowly add to our understanding of species limits of this elusive group of bats.

