











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los bosques y selvas proveen bienes y servicios ambientales importantes, que pueden contribuir al mejoramiento de la economía local (Méndez et al., 2018). Los bosques templados son ecosistemas con gran diversidad en el continente americano; mismo que se extienden desde Estados Unidos de América hasta Honduras (Mora-Donjuán y Alanís-Rodríguez, 2016).
El conocimiento de la estructura y composición de comunidades vegetales es fundamental, ya que permite crear estrategias que promuevan el crecimiento y desarrollo de las masas forestales, lo que asegura un aprovechamiento racional sin comprometer los escenarios originales (Aguirre-Calderón, 2015; Manzanilla et al., 2020). Además, es necesario conocer la riqueza, composición y grado de similitud de las comunidades (Domínguez et al., 2018); así como los componentes que influyen en la diversidad y riqueza, entre los que destacan los factores ambientales y las características físicas del terreno (Saldaña, 2013).
La diversidad de especies puede disminuir hacia latitudes y altitudes mayores (Malizia et al., 2020). García-Aguilar et al. (2017) confirman que la capacidad de desarrollo de los bosques está estrechamente ligada a las condiciones físicas del terreno. De igual manera, McIntire et al. (2016) ratifican que la exposición es un factor limitante para el establecimiento de especies poco tolerantes a la sombra.
En la actualidad, se han realizado diversos estudios de la estructura y composición florística en ecosistemas forestales, los cuales se centran en los cambios sobre el gradiente altitudinal (Alves et al., 2010; Rascón et al., 2018); sin embargo, no se ha considerado la influencia de la exposición geográfica, lo que resulta en información insuficiente para México.
Por lo anterior, el objetivo de la presente investigación fue determinar diferencias en la estructura, riqueza, composición y grado de similitud de las comunidades vegetales, respecto a la exposición y el intervalo de altitud en un bosque templado del estado de Durango. Se analizan las siguientes hipótesis: (i) la exposición norte presentará mayores valores de área basal; (ii) conforme aumente la altura, la riqueza y diversidad de especies disminuirá; y (iii) el grado de similitud en la composición de especies estará definida por la exposición.
Materiales y Métodos
El área de estudio se ubicó en el sistema montañoso denominado Sierra Madre Occidental en el estado de Durango, entre las coordenadas geográficas 26°26'25.7" N, 106° 03'57.5" O y 23°10'29.5" N, 105° 2' 09.2" O (Figura 1).

Conforme con la clasificación de Köppen modificada por García (1988), los tipos de clima predominantes en el área son templado subhúmedo en cuatro subdivisiones: (A)c(w2), (A)c(w1), C(E)(w2) y C(E)(W2)(x'). La variación de temperatura oscila entre 12 y 18 °C, con excepción de algunas áreas que presentan valores de 10 a 12 °C; las lluvias suelen presentarse en verano y sequías en invierno (Quiñones et al., 2012). Los tipos de suelo conforman asociaciones entre Regosol, Litosol, Cambisol y Feozem (INEGI, 2007). Los tipos de vegetación existentes son bosque de pino, bosque de encino, bosque de pino-encino, encino-pino y vegetación secundaria arbustiva en todas las comunidades vegetales (González et al., 2012).
Obtención y análisis de información
La información dasométrica se obtuvo de 30 parcelas de monitoreo forestal permanente cada una de 2 500 m2, ubicadas aleatoriamente en seis áreas, tres intervalos altitudinales (entre 1 500 y 3 000 m), con cinco parcelas por exposición (norte y sur). Se consideraron el diámetro normal (Dn> 7.5 cm), el cual se midió con cinta diamétrica 122450 Ben Meadows; altura total (H), con un clinómetro Suunto Pm5/360pc; y el registro por especie, los nombres científicos se corroboraron en la página web The Plant List (http://www.theplantlist.org/).
Para cada área se calculó el Índice de Valor de Importancia Ecológica (IVI); se obtuvieron valores porcentuales de 0 a 100 (Alanís-Rodríguez et al., 2020) por especie a partir de la sumatoria de los parámetros estructurales relativos de abundancia (densidad), frecuencia y área basal (Whittaker, 1972; Moreno, 2001). La riqueza de especies se determinó mediante el Índice de Margalef y la diversidad alfa se estimó con el Índice de Diversidad Verdadera de Shannon (Jost, 2006). El cálculo de cada uno de los parámetros se realizó con las ecuaciones del Cuadro 1.
Cuadro 1 Ecuaciones para calcular parámetros de estructura y diversidad.
| Parámetro | Ecuación | Descripción |
|---|---|---|
| Abundancia |
|
A
i
= Abundancia absoluta AR i = Abundancia relativa de la especie i, con respecto a la abundancia total N i = Número de individuos de la especie i S = Superficie de muestreo (ha) |
| Frecuencia |
|
F
i
= Frecuencia absoluta FR i = Frecuencia relativa de la especie i, con respecto a la frecuencia total P i = Número de sitios en la que la especie se encuentra presente i NS = Número total de sitios de muestreo |
| Área basal |
|
D
i
= Dominancia absoluta DR i = Dominancia relativa de la especie i, con respecto a la dominancia total Ab = Área basal de la especie i, S = Superficie (ha) |
| IVI |
|
Ar
i
= Abundancia relativa Fr i = Frecuencia relativa Dr i = Dominancia relativa |
| Índice de Margalef |
|
S = Número de especies presentes N = Número total de individuos n = Número de individuos de la especie i |
| Índice de Diversidad Verdadera de Shannon |
1 D = 𝑒𝑥𝑝(𝐻´) |
pi = Abundancia proporcional de la i-ésima especie ni = Número de individuos por especie N = Total de individuos presentes 1 D = Índice de Diversidad Verdadera de Shannon exp = Exponencial H´ = Índice de Diversidad de Shannon |
La verificación del cumplimiento de los supuestos de normalidad de los residuales, se hizo con base en la prueba estadística de Shapiro-Wilk; además, se revisó la homogeneidad de varianzas mediante la Prueba de Levene. Para determinar diferencias entre las áreas se aplicó un análisis de varianza (ANOVA) de dos factores (exposición e intervalo de altitud). Se utilizó la prueba de comparaciones múltiples de Tukey para determinar diferencias con un nivel de significancia de p<0.05. Los análisis estadísticos se realizaron utilizando el Software IBM © SPSS © Statistic versión 19 (Zar, 2010).
Para el cálculo de la diversidad beta, definida como el reemplazamiento de especies en comunidades con escenarios ambientales diferentes (Whittaker, 1972), se desarrolló un modelo de clasificación utilizando algoritmos de similitud de muestras, mediante el cálculo del porcentaje de similitud entre ellas (0 % a 100 %) con el dendrograma de similitud de Bray-Curtis, el cual es adecuado para efectuar análisis del comportamiento de especies vegetales (Rascón et al., 2018). El análisis se llevó a cabo por medio del software Past 4.01 (Hammer, 2001), en el cual se usó como matriz de agrupamiento el parámetro de abundancia en los diferentes intervalos de altitud por área.
Resultados
Se registraron 48 especies (Cuadro 2), distribuidas en ocho familias; de ellas, Fagaceae resultó con un mayor número taxones e integró a 39.58 % del total; seguida de Pinaceae, con 31.25 %; Convolvulaceae y Betulaceae presentaron los valores más bajos.
Cuadro 2 Lista de especies arbóreas presentes en el área de estudio.
| Familia | Especie |
|---|---|
| Betulaceae | Alnus jorullensis Kunth |
| Convolvulaceae | Ipomoea arborescens Humb. et Bonpl. |
| Cupressaceae | Juniperus deppeana Steud. |
| Cupressaceae | Cupressus lusitanica Mill. |
| Ericaceae | Arbutus arizonica (A. Gray) Sarg. |
| Ericaceae | Arbutus bicolor S. González, M. González & P.D. Sorensen |
| Ericaceae | Arbutus madrensis S. González |
| Ericaceae | Arbutus tesselata P.D. Sorensen |
| Ericaceae | Arbutus xalapensis Kunth |
| Ericaceae | Comarostaphylis polifolia Kunth |
| Fabaceae | Lysiloma acapulcense Benth. |
| Fabaceae | Acacia pennatula Benth. |
| Fagaceae | Quercus albocincta Trel. |
| Fagaceae | Quercus castanea Née |
| Fagaceae | Quercus coccolobifolia Trel. |
| Fagaceae | Quercus crassifolia Humb. & Bonpl. |
| Fagaceae | Quercus depressipes Trel. |
| Fagaceae | Quercus durifolia Seemen ex Loes. |
| Fagaceae | Quercus elliptica Née |
| Fagaceae | Quercus fulva Liebm. |
| Fagaceae | Quercus gentryi C. H. Mull. |
| Fagaceae | Quercus jonesii Trel. |
| Fagaceae | Quercus laeta Liebm. |
| Fagaceae | Quercus obtusata Humb. & Bonpl. |
| Fagaceae | Quercus resinosa Liebm. |
| Fagaceae | Quercus rugosa Née |
| Fagaceae | Quercus salicifolia Benth. |
| Fagaceae | Quercus scytophylla Liebm. |
| Fagaceae | Quercus sideroxyla Bonpl. |
| Fagaceae | Quercus urbanii Trel. |
| Fagaceae | Quercus viminea Trel. |
| Pinaceae | Pinus arizonica Engelm. |
| Pinaceae | Pinus cembroides Zucc. |
| Pinaceae | Pinus chihuahuana Martínez |
| Pinaceae | Pinus cooperi C.E. Blanco |
| Pinaceae | Pinus douglasiana Martínez |
| Pinaceae | Pinus durangensis Martínez |
| Pinaceae | Pinus engelmannii Carrière |
| Pinaceae | Pinus herrerae Martínez |
| Pinaceae | Pinus leiophylla Schiede ex Schltdl. et Cham. |
| Pinaceae | Pinus lumholtzii B. L. Rob et Fernald |
| Pinaceae | Pinus luzmariae Pérez de la Rosa |
| Pinaceae | Pinus oocarpa Schiede ex Schltdl. |
| Pinaceae | Pinus strobiformis Engelm. |
| Pinaceae | Pinus teocote Schiede ex Schltdl. et Cham. |
| Pinaceae | Pseudotsuga menziesii (Mirb.) Franco |
| Rosaceae | Crataegus mexicana DC. |
| Rosaceae | Prunus serotina Ehrh. |
Abundancia (densidad). Las áreas 1 y 2 tuvieron valores más altos. En exposición sur, las especies con mayor abundancia fueron: P. arizonica Engelm. con 600 ind ha-1 y J. deppeana Steud. con 66 ind ha-1. En exposición norte coincidieron P. arizonica con 344 ind ha-1 y Q. sideroxyla Bonpl. con 111 ind ha-1. Las áreas 3 y 5 presentaron similitud en el total de individuos por hectárea. Los taxones más abundantes en el área 3 fueron: Q. laeta Liebm., con 122 ind ha-1 y P. leiophylla Schiede ex Schltdl. et Cham., con 68 ind ha-1. En el área 5, P. durangensis y P. oocarpa Schiede ex Schltdl. tuvieron más presencia (Cuadro 3). Las áreas 4 y 6 registraron una diferencia en comparación con los intervalos de mayor altitud (p<0.05). En el área 4, las especies con más densidad fueron: Q. crassifolia Humb. & Bonpl., con 121 ind ha-1 y P. leiophylla, con 83 ind ha-1. En el área 6, los taxa más abundantes fueron: P. durangensis, con 77 ind ha-1 y Q. resinosa Liebm., con 47 ind ha-1.
Cuadro 3 Parámetros estructurales estimados por área e intervalo de altitud (IVI ordenado de mayor a menor valor porcentual, se incluyen solo las tres especies con valor más alto.
| Exposición/ Altitud | Especie | Densidad | Frecuencia | Dominancia (Área basal) | IVI | |||
|---|---|---|---|---|---|---|---|---|
| Absoluta (N ha-1) |
Relativa (%) |
Absoluta | Relativa (%) |
Absoluta (m ha-1) |
Relativa (%) |
|||
| Sur 2 700-3 000 msnm (Área 1) | P. arizonica | 600 | 75.6 | 5 | 25 | 21.26 | 74.98 | 58.53 |
| J. deppeana | 66 | 8.27 | 5 | 25 | 0.15 | 5.32 | 12.86 | |
| P. strobiformis | 32 | 4.03 | 4 | 20 | 1.51 | 4.02 | 9.35 | |
| Norte 2 700-3 000 msnm (Área 2) | P. arizonica | 344 | 45.03 | 4 | 13.33 | 8.9 | 31.71 | 30.02 |
| Q. sideroxyla | 111 | 14.55 | 5 | 16.67 | 8.81 | 31.38 | 20.87 | |
| A. xalapensis | 73 | 9.53 | 3 | 10 | 2.02 | 7.2 | 8.91 | |
| Sur 2 100-2 400 msnm (Área 3) | Q. laeta | 122 | 24.21 | 3 | 10.71 | 4.65 | 19.28 | 18.04 |
| P. leiophylla | 68 | 13.49 | 2 | 7.14 | 4.22 | 17.5 | 12.71 | |
| Q. sideroxyla | 58 | 11.51 | 1 | 3.57 | 4 | 16.6 | 10.53 | |
| Norte 2 100-2 400 msnm (Área 4) | Q. crassifolia | 121 | 29.38 | 2 | 5.56 | 2.25 | 14.32 | 16.42 |
| P. teocote | 50 | 12.26 | 4 | 11.11 | 2.72 | 17.29 | 13.55 | |
| P. leiophylla | 66 | 16.15 | 2 | 5.56 | 2.94 | 18.75 | 13.49 | |
| Sur 1 500-1 800 msnm (Área 5) | P. durangensis | 206 | 39.24 | 2 | 7.14 | 14.29 | 47.97 | 31.47 |
| P. oocarpa | 177 | 33.74 | 3 | 10.71 | 5.99 | 20.12 | 21.53 | |
| Q. viminea | 8 | 1.53 | 3 | 10.71 | 0.62 | 2.08 | 4.78 | |
| Norte 1 500-1 800 msnm (Área 6) | P. durangensis | 76.8 | 21.92 | 2 | 8.33 | 3.36 | 21.06 | 17.1 |
| Q. gentryi | 25.6 | 7.31 | 2 | 8.33 | 1.49 | 9.35 | 8.33 | |
| P. oocarpa | 26.4 | 7.53 | 2 | 8.33 | 1.41 | 8.88 | 8.25 | |
Área Basal. La exposición sur presentó el área basal más alta en los tres intervalos de altitud. Para el intervalo 1 (2 700-3 000 m) el promedio fue de 56.42 m2 ha-1; sobresalió P. arizonica con 21.26 m2 ha-1 en el área 1 (Cuadro 3). La prueba de ANOVA no evidenció diferencias significativas para las áreas 4 y 6; sin embargo, el promedio fue menor con 15.70 m2 ha-1 y 15.94 m2 ha-1, respectivamente. Las especies con mayor área basal fueron P. leiophylla con 2.94 y P. durangensis con 3.36 m2 ha-1 (Figura 2B).
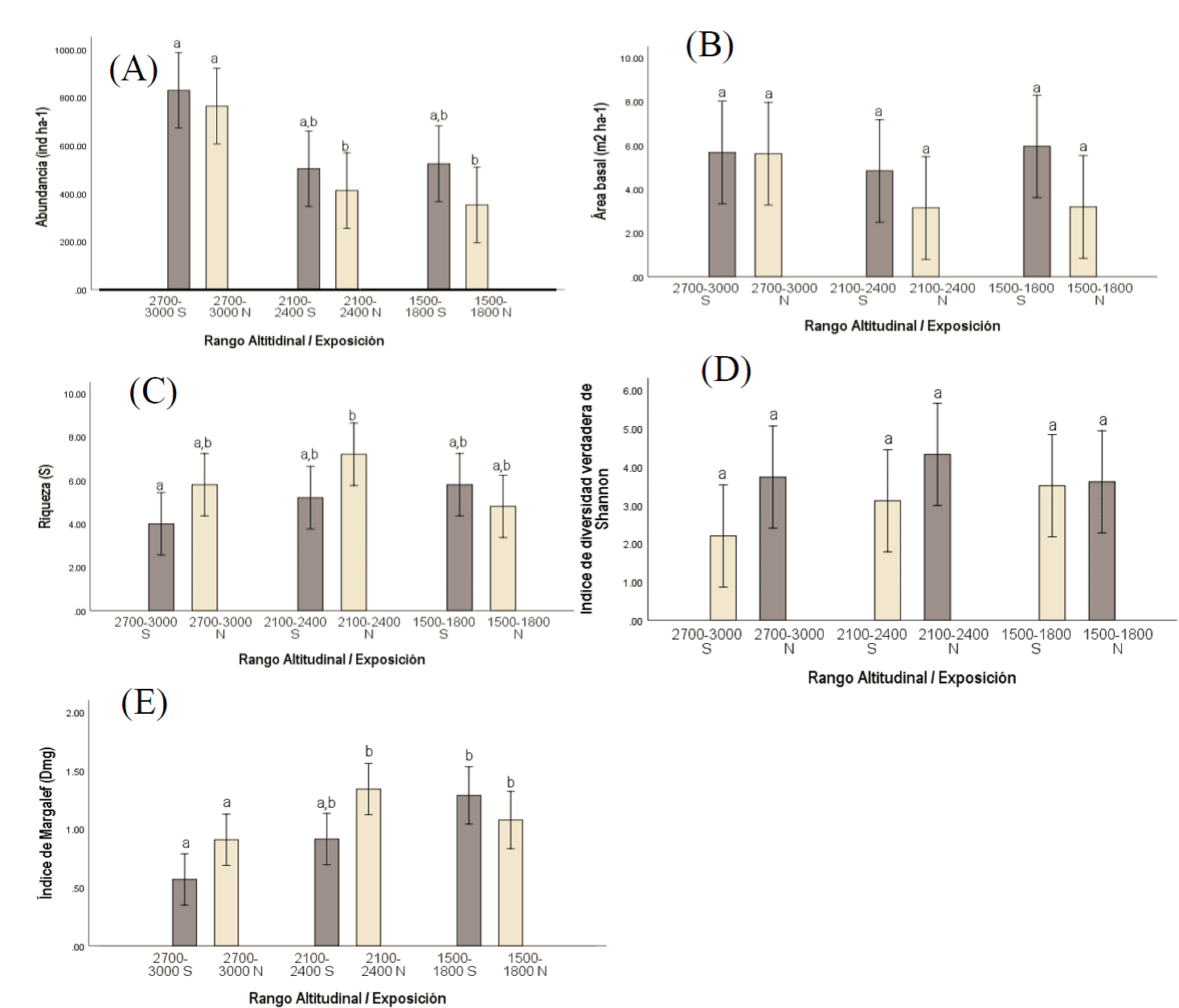
Figura 2 Medias y error estándar de (A) Abundancia (ind ha-1), (B) Área basal (m2 ha-1), (C) Riqueza (S), (D) Índice de diversidad verdadera de Shannon (1D), (E), Índice de Margalef (Dmg). Medias con letras diferentes (a, b) indican diferencias (p<0.05).
Índice de Valor de Importancia Ecológica (IVI). Los géneros Pinus y Quercus mostraron mayor porcentaje de IVI; P. arizonica alcanzó el valor más alto en el intervalo 1, norte y sur; Q. laeta Liebm. en exposición sur y Q. crassifolia en la norte dominaron los intervalos de altitud media. En el intervalo de altura más bajo, el mayor IVI lo obtuvo P. durangensis; en el Cuadro 3 se muestran las tres especies de más importancia por área.
El dendrograma de comunidades vegetales en función de intervalos de altitud demostró que existe similitud entre las áreas 1 y 2, con 65 % y entre las áreas 4 y 6, con 46 % (Figura 3). Lo anterior indica que en estas existe una composición florística similar, cuyas especies dominantes suelen ser las mismas. Asimismo, la exposición sur del área 5 presentó valores de similitud menores, en comparación con el resto; se observó, de manera general, que en sitios con altitudes más bajas, la exposición es precisamente la que define el grado de similitud. Las especies generalistas que se registraron en ambas exposiciones y en los tres intervalos altitudinales fueron: P. durangensis, Q. sideroxyla Bonpl., J. deppeana y A. xalapensis Kunth. En sitios con menor altura, se presentaron taxones particulares por tipo de exposición: Q. coccolobifolia Trel. en exposición norte; y A. pennatula Benth., L. acapulcense Benth., P. serotina Ehrh. e I. arborescens Humb. et Bonpl. en exposición sur.

Figura 3 Dendrograma de similitud basado en análisis Bray Curtis entre áreas con diferentes intervalos altitudinales.
Riqueza. En las áreas 1 y 4 se obtuvieron diferencias (p<0.05), con una disminución en el número de especies en sitios con mayor altitud. Se apreció que el número de taxones dentro del área 4 fue cerca del doble, en comparación con el área 1. En las áreas restantes el intervalo promedio fue de cinco taxones por sitio (Figura 2C).
Índices de Diversidad. La diversidad verdadera de Shannon no mostró diferencias (p<0.05) entre áreas, con un promedio de 2.94 ± 1.58 en exposición sur y 3.89 en la norte (Figura 2D). En relación al Índice de Margalef, se observaron diferencias (p<0.05) en las áreas de mayor altitud, con respecto a las de menor altura. Los valores promedio fueron de 0.73 para ambas exposiciones en el intervalo 1, y de 0.98 en el intervalo 3 (Figura 2E).
Discusión
Los resultados obtenidos en densidad coinciden con los citados por Delgado et al. (2016), quienes calcularon 565 ind ha-1 y 16 taxones en bosques templados de Durango, en un intervalo altitudinal de 2 400 a 2 500 m. Por el contrario, López-Hernández et al. (2017) registraron valores más bajos, 389 ind ha-1 y 11 especies en bosques del estado de Puebla, lo cual puede atribuirse a que los bosques evaluados presentan actividades más intensivas de aprovechamiento, en comparación con las del presente estudio.
Respecto al área basal, se identificó que al aumentar la altitud, lo hace el área basal tal como lo señalan Muñoz et al. (2020); asimismo, la exposición y la pendiente pueden influir en la productividad de diversas áreas. Los resultados del estudio muestran que el área basal es mayor en sitios con más altitud, con valores de hasta el doble. Los resultados son similares a los estimados por Graciano-Ávila et al. (2017), quienes calcularon un área basal semejante en bosques templados de Durango.
En la mayoría de los bosques templados de México, Pinus y Quercus suelen ser los más representativos, estos géneros engloban un mayor número de especies con variables dasométricas más dominantes; por lo cual tienden a presentar un valor de importancia ecológica más alto (Domínguez et al., 2018). En este caso, existieron variaciones en las especies con un valor de IVI superior; de tal manera que áreas de alta y menor altitud registraron mayor IVI en Pinus; lo anterior coincide con Hernández-Salas et al. (2013) y Graciano-Ávila et al. (2019), autores que consignan a P. durangensis como la especie de más importancia ecológica en bosques de Chihuahua y Durango. Paredes et al. (2019) concuerdan con los resultados obtenidos en el presente estudio, y citan a Q. crassifolia como el taxón con más importancia ecológica en un bosque templado húmedo de Hidalgo.
La altitud, inclinación y exposición tienen gran influencia en la composición florística y la riqueza de especies en las comunidades vegetales. Siles et al. (2017) registran 27 taxones sobre un gradiente altitudinal de 1 300 a 1 500 m; datos que se asemejan con los aquí documentados, en los cuales se presentaron de 18 a 23 especies en el intervalo altitudinal bajo. De igual forma, Castellanos-Bolaños et al. (2019) coinciden en que a mayor altitud, el número de taxa es menor.
En cuanto a la diversidad, Domínguez et al. (2018) indican registros semejantes en altitudes similares para un bosque templado del ejido Ruiz Cortínez, en la región de El Salto, Durango; donde los valores obtenidos para diversidad son similares a los de este estudio.
Para evaluar la diversidad y riqueza de especies entre comunidades es necesario conocer las características biológicas y su distribución proporcional (Moreno, 2001). El valor del Índice de Margalef fue inferior en las altitudes mayores, como lo señalan Báez et al. (2015), quienes indican que las bajas temperaturas y características topográficas contribuyen a la disminución de especies. Por otra parte, el alto número de taxones en altitudes menores es similar a lo estimado por Clark et al. (2015), quienes consignan alta riqueza en parcelas localizadas a baja altitud.
Zarco et al. (2010) citan que la exposición influye sobre el desarrollo de diferentes especies, sobre todo en altitudes media y baja, información acorde a lo registrado en la presente investigación, cuyo número de taxa fue superior en la exposición sur.
La similitud entre comunidades está ligada a la altitud y exposición, entre otros factores (Chust et al., 2006), los cuales determinan las especies que pueden adaptarse a diferentes comunidades vegetales. De acuerdo con lo anterior, es posible observar especies generalistas y específicas adaptadas a diferentes áreas con condiciones particulares.
Hernández et al. (2013) indican que la agrupación depende, en gran medida, del grado de adaptación de los taxones; de tal manera que diferentes especies de coníferas y encinos pueden aparecer en sitios con características diversas; lo anterior confirma los datos obtenidos para distintas especies de Pinus y Quercus, las cuales están presentes en todas las áreas estudiadas. De igual forma, se coincide con Delgado et al. (2016), quienes señalan que existen especies particulares que se establecen en microhábitats muy definidos, como se observa en este estudio, en el que se registraron taxones, únicamente, en intervalos de altitud baja y exposición sur.
Conclusiones
El área basal no presenta diferencias significativas entre las diferentes áreas e intervalos altitudinales; sin embargo, la densidad es más alta en sitios con una altura mayor, por lo cual se rechaza la hipótesis planteada al inicio del estudio.
Referente a la riqueza y la diversidad de especies, se cumple con el supuesto de la hipótesis planteada, ya que es mayor en áreas con exposición sur. De igual forma, los sitios con menor altitud registran el número más alto de especies, además se identifican especies generalistas y particulares en áreas específicas.
Los resultados indican que dentro de las áreas evaluadas en los diferentes intervalos altitudinales, la estructura, composición florística y riqueza de especies están fuertemente influenciadas por las características topográficas del terreno. Es posible identificar que el grado de similitud de comunidades vegetales varía de acuerdo con el intervalo altitudinal y exposición de laderas, donde la temperatura puede contribuir o demeritar el desarrollo de las especies.

