











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El venado cola blanca (Odocoileus virginianus Mearns, 1898) es la especie cinegética más importante en el mundo desde la perspectiva económica (Villarreal et al., 2014; Retana et al., 2015). En México, O. virginianis ssp. texanus es la subespecie de mayor talla corporal y valor cinegético, incluida en los registros de trofeos internacionales (Villarreal et al., 2014). Por ello, desde la década de 1960, propietarios de predios en el noreste de México han enfocado las actividades de sus áreas hacia su aprovechamiento y conservación, como una alternativa de producción (Gallina et al., 2009; Hernández et al., 2018).
En México y en el sur de Texas, Estados Unidos de América, se le relaciona principalmente con matorrales desérticos (Mandujano et al., 2010); debido a que proveen alimentos y protección a machos y a hembras con crías. El hábitat ideal para el venado cola blanca incluye una asociación de vegetación arbustiva y pastos de porte medio y alto (Bello et al., 2006). Además, la presencia de nopales, agaves y otras suculentas es necesaria para amortiguar periodos prolongados de sequía (Espino-Barros y Fuentes, 2005). En ambos países se han realizado estudios profundos sobre requerimientos de su hábitat y estructura poblacional (Stocker y Gilbert, 1977; Gallina, 1993; Arceo et al., 2005; Bello et al., 2006), en particular sobre este tipo de matorrales (Gallina et al., 2010; Gallina y Bello, 2010; Gallina y Bello, 2014). En el estado de Coahuila, se carece de investigaciones referentes a la composición, diversidad e importancia de esta comunidad para el desarrollo y mantenimiento de las poblaciones de venado cola blanca.
Conocer e interpretar la función de la cobertura vegetal en el desarrollo de las poblaciones de O. virginianus es fundamental para el correcto manejo de su hábitat (Stocker y Gilbert, 1977; Delfín et al., 2009). Con el propósito de que el personal técnico, autoridades y propietarios de predios dedicados al manejo de esta especie tengan información pertinente para la implementación de herramientas y acciones dirigidas a su conservación en los matorrales desérticos del noreste de México, los objetivos del presente trabajo fueron identificar y evaluar la diversidad, composición y productividad de un matorral desértico micrófilo para inferir su importancia en la conservación de Odocoileus virginianus, en una localidad de manejo controlado.
Materiales y Métodos
Área de estudio
El estudio se realizó en un área de 1 030 ha destinada al venado cola blanca, dentro de la Unidad de Manejo para la Conservación de la Vida Silvestre (UMA) Rancho San Juan (clave DGVS-CR-EX-3133-COA), en el municipio Monclova, Coahuila (Figura 1), ubicada a 38 km en línea recta al este de la cabecera municipal y a 43 km al oeste del municipio Candela, entre los 26°49’31.11’’ N y los 101°01’57.77’’ O.

Figura 1 Localización y tipo de vegetación de la UMA Rancho San Juan, Monclova, Coahuila, México (INEGI, 2013).
El clima es seco (BSohw), con una temperatura promedio anual de 21 °C. La precipitación anual varía de 200 mm a 900 mm, con un intervalo altitudinal de 600 a 1 000 m (García, 1988). El tipo de vegetación dominante es matorral desértico micrófilo, con asociación de pastizal mediano abierto (Miranda y Hernández, 1963). Las especies vegetales representativas son arbustos como Acacia berlandieri Benth., Acacia rigidula Benth., Castela texana Torr & Gray, Celtis pallida Torr., Flourensia cernua DC., Forestiera angustifolia Torr., Karwinskia humboldtiana (Schult.) Zucc., Larrea tridentata (Sessé & Moc. ex DC.) Coville; pastos del género Bouteloua y suculentas como Opuntia engelmannii Salm-Dyck ex Engelm., entre las principales.
Descripción de la cobertura vegetal
Para evaluar la composición y estructura de la vegetación, se utilizó el método de línea Canfield, porque permite estimar valores relativos de abundancia, frecuencia y dominancia (González et al., 2012). Se colocaron 18 líneas permanentes, al azar de 25 m de longitud en cada estación del año: otoño (octubre 2018), invierno (febrero 2019), primavera (mayo 2019) y verano (agosto 2019). Se midió la altura y cobertura de las plantas que interceptaron la línea. Las especies se clasificaron de acuerdo a su altura en: estrato bajo (≤ a 50 cm), medio (51-150 cm) y alto (> a 150 cm); las cuales se midieron desde la base hasta la punta, con cinta métrica rígida de 1 1/4’’ de ancho y 3 m de longitud marca Truper. Con los parámetros evaluados se determinó el Índice de Valor de Importancia (IVI) según Curtis y Mclntosh (1951), mediante la Ecuación 1:
Donde:
AR = Abundancia relativa
FR = Frecuencia relativa
DR = Dominancia relativa
La diversidad específica (() del matorral en cada estación del año, se estimó con la Ecuación 2 para calcular el índice de Diversidad de Shannon (1948).
Donde:
P
i
= Proporción de individuos de la especie i
respecto al total de individuos en la estación (abundancia relativa de la
especie i):
n = Número de individuos de la especie i
N = Número de todos los individuos de todas las especies en la estación
Para conocer si existe diferencia significativa en la diversidad de plantas entre
estaciones, se aplicó la prueba t-student (
Estimación de la producción de biomasa
La productividad de forraje en las cuatro estaciones del año se expresó como la cantidad de biomasa por estrato vegetal en kg ha-1 (Fulbright y Ortega, 2006). Con el Método de Adelaide (Foroughbakhch et al., 2005) se evaluó la producción de biomasa del estrato alto y medio, en 18 parcelas de 50 m2 y 25 m2, respectivamente. El método consiste en tomar una unidad de referencia de cada planta dentro de las parcelas (representativa en forma y densidad foliar de toda la planta). Con ella se estimó el número de unidades por ejemplar y especie muestreada. Sin embargo, para el estrato bajo (pastos y herbáceas) se realizó su corta total en 18 parcelas de 1 m2 (Chávez, 2000). Las muestras de biomasa de pastos, herbáceas y las unidades de referencia se colocaron en bolsas de papel, se etiquetaron y secaron en un horno tipo INOX. 120VAC. 60HZ. a 75 °C hasta un peso constante. Por último, las muestras se pesaron en una balanza ENTRIS 8201-1S para obtener el peso seco.
Resultados y Discusión
Diversidad y composición de la cobertura vegetal
Se registró un total de 42 especies vegetales pertenecientes a 21 familias, de las cuales Poaceae (siete), Asteraceae (seis.), Fabaceae (cinco) y Cactaceae (cuatro) fueron las más comunes. De ellas, 16 estuvieron presentes durante las cuatro estaciones del año (Cuadro 1).
Cuadro 1 Índice de Valor de Importancia (IVI) de las especies registradas en las cuatro estaciones del año en la UMA Rancho San Juan, Monclova, Coahuila, México.
| Familia | Especie | Estrato | Índice de Valor de Importancia (%) | |||
|---|---|---|---|---|---|---|
| Primavera | Verano | Otoño | Invierno | |||
| Fabaceae | Acacia berlandieri Benth. | Alto | 7.13 | 12.25 | 2.48 | 1.77 |
| Fabaceae | Acacia rigidula Benth. | Alto | 19.39 | 14.58 | 2.39 | 6.02 |
| Agavaceae | Agave lechuguilla Torr. | Medio | 59.78 | 35.31 | 27.99 | 29.66 |
| Agavaceae | Agave scabra Salm-Dyck | Medio | 7.53 | 4.26 | 3.38 | 8.73 |
| Verbenaceae | Aloysia macrostachya Moldenke | Medio | 2.68 | 4.9 | 3.39 | 1.21 |
| Poaceae | Aristida adscensionis L. | Bajo | - | - | 1.87 | - |
| Poaceae | Bothriochloa laguroides (DC.) Herter | Bajo | - | - | 3.29 | - |
| Poaceae | Bouteloua curtipendula (Michx.) Torr. | Bajo | 2.22 | - | 8.22 | 2.08 |
| Poaceae | Bouteloua gracilis (Kunth) Lag. ex Griffiths | Bajo | - | 29.78 | 22.99 | - |
| Asteraceae | Brickellia glutinosa S. Watson | Medio | - | - | - | 2.27 |
| Fabaceae | Calliandra conferta Benth. | Medio | 15.09 | 8.97 | - | - |
| Simaroubaceae | Castela texana Torr & Gray | Medio | - | - | - | 1.17 |
| Cannabaceae | Celtis pallida Torr. | Alto | 1.44 | 0.97 | 1.14 | 1.44 |
| Poaceae | Cenchrus ciliaris L. | Bajo | 10.68 | 1.02 | 7.66 | 107 |
| Cactaceae | Cylindropuntia leptocaulis (DC.) F. M. Knuth | Medio | 9.19 | 1.92 | 5.43 | 7.06 |
| Fabaceae | Dalea greggii A. Gray | Bajo | 5.93 | - | - | - |
| Ebenaceae | Diospyros texana Scheele | Alto | 1.61 | - | - | - |
| Ephedraceae | Ephedra pedunculata Engelm. ex S. Wats. | Medio | 35.4 | - | - | - |
| Euphorbiaceae | Euphorbia antisyphilitica Zucc. | Medio | 17.39 | 23.8 | 12.94 | 9.53 |
| Cactaceae | Ferocactus sp. Britton & Rose | Bajo | 1.43 | 5.85 | - | 0.99 |
| Asteraceae | Flourensia cernua DC. | Medio | - | 17.87 | 20.13 | 31.92 |
| Oleaceae | Forestiera angustifolia Torr. | Medio | 11.01 | 1.34 | 0.9 | 1.38 |
| Asteraceae | Gochnatia hypoleuca (DC.) A. Gray | Medio | 1.03 | - | 1.91 | - |
| Zygophyllaceae | Guaiacum angustifolium Engelm. | Medio | 14.93 | 4.63 | 7.13 | 8.09 |
| Bromeliaceae | Hechtia glomerata Mez. | Bajo | 1.12 | 2.79 | 1.69 | - |
| Poaceae | Hilaria mutica (Buckley) Benth. | Bajo | - | 62.02 | 86.59 | - |
| Euphorbiaceae | Jatropha dioica Sessé ex Cerv. | Medio | 20.24 | 9.55 | 9.96 | 18.62 |
| Rhamnaceae | Karwinskia humboldtiana (Schult.) Zucc. | Medio | 7.55 | 4.2 | 1.94 | 1.14 |
| Koeberliniaceae | Koeberlinia spinosa Zucc. | Medio | - | - | - | 2.4 |
| Zygophyllaceae | Larrea tridentata (Sessé & Moc. ex DC.) Coville | Medio | - | - | 1.1 | - |
| Scrophulariaceae | Leucophyllum frutescens (Berland) I. M. Johnston | Medio | 4.61 | 2.23 | 1.72 | 2.36 |
| Verbenaceae | Lippia graveolens Kunth | Medio | - | 10.72 | 19.1 | 9.05 |
| Cactaceae | Opuntia engelmannii Salm-Dyck ex Engelm. | Medio | 25.23 | 15.16 | 13.08 | 23.62 |
| Cactaceae | Opuntia microdasys (Lehm.) Pfeiff. | Medio | 1.46 | 1.03 | - | - |
| Asteraceae | Parthenium argentatum A. Gray | Bajo | 1.32 | - | - | - |
| Asteraceae | Parthenium hysterophorus L. | Bajo | - | - | 8.7 | - |
| Poaceae | Paspalum notatum Flüggé | Bajo | - | 12.41 | 9.48 | 7.03 |
| Achatocarpaceae | Phaulothamnus spinescens A. Gray | Medio | - | - | 1.71 | - |
| Fabaceae | Prosopis glandulosa Torr. | Medio | 14.64 | 10.96 | 7.99 | 12.79 |
| Lamiaceae | Salvia coccinea Buc'hoz ex Etl. | Medio | - | 1.48 | - | - |
| Apocynaceae | Telosiphonia macrosiphon (Torr.) Henrickson | Medio | - | - | - | 2.67 |
| Asteraceae | Wedelia texana (A. Gray) B. L. Turner | Bajo | - | - | 2.55 | - |
| Asparagaceae | Yucca filifera Hort. ex Engelm. | Alto | - | - | 1.17 | - |
Algunas especies registradas en este estudio se han consignado previamente en la dieta del venado cola blanca en el noreste de México, por ejemplo: Acacia rigidula Benth., Acacia berlandieri Benth., Calliandra conferta Benth., Celtis pallida Torr., Dyospiros texana Scheele, Forestiera angustifolia Torr., Karwinskia humboldtiana (Schult.) Zucc., Opuntia engelmannii Salm-Dyck ex Engelm. y Prosopis glandulosa Torr. (Ramírez, 2004). Sin embargo, se consideran decrecientes, debido a que disminuye su disponibilidad en función del ramoneo (Dyksterhuis, 1948; Ramírez et al., 1996; Ramírez, 2004). En particular, Acacia berlandieri y Acacia rigidula son plantas forrajeras importantes para el venado cola blanca. Ramírez et al. (1996) concluyeron que en el noreste de México ambas especies constituyen 75 % de la dieta anual del venado. Además, sus inflorescencias representan una importante fuente de alimento, necesario para recuperar el gasto energético de la época reproductiva.
En el matorral existen taxones de emergencia (consumidas en ausencia o baja disponibilidad de las preferidas), también consideradas como crecientes, ya que aumentan su disponibilidad en relación al bajo consumo (Dyksterhuis, 1948; Ramírez, 2004) e incluyen: Larrea tridentata (Sessé & Moc. ex DC.) Coville, Lippia graveolens Kunth, Parthenium argentatum A. Gray, Parthenium hysterophorus L., Phaulothamnus spinescens A. Gray, Salvia coccinea Buc'hoz ex Etl., Wedelia texana (A. Gray) B. L. Turner; además de, algunos pastos del género Bouteloua. Aunque estas no son preferidas por el venado, pueden ser fundamentales en épocas de baja disponibilidad y deficiente calidad de forraje. Sin embargo, Aguiar et al. (2011) indican que las plantas poco consumidas por el venado producen efectos adversos en la tasa de reproducción. Por ello, es indispensable mantener zonas con leguminosas.
Por otro lado, en zonas áridas y semiáridas la presencia del venado cola blanca está condicionada por la disponibilidad de especies como las suculentas, que funcionan como plantas amortiguadoras durante la temporada de sequía (Gallina y Bello, 2010). En este sentido, Opuntia engelmannii se identificó como dominante dentro del matorral, y se observaron evidencias de ramoneo por venados (Figura 2). Es una planta forrajera de interés para Odocoileus virginianus en el noreste de México y sur de Texas, por su alto contenido de agua (90 %) (Ramírez et al., 2000). Otras suculentas como Agave lechuguilla Torr., Agave scabra Salm-Dyck, Hechtia glomerata Mez., Cylindropuntia leptocaulis (DC.) F. M. Knuth y Opuntia microdasys (Lehm.) Pfeiff., se identificaron en el área de estudio. De ellas, solo Cylindropuntia leptocaulis se registra en la dieta del venado para el noreste de México.

Figura 2 Especies dominantes en relación al Índice de Valor de Importancia por estación del año, en la UMA Rancho San Juan, Monclova, Coahuila, México.
Durante la primavera y el verano, las arbustivas presentaron mayor disponibilidad, 72.43 % y 50.07 %, respectivamente (Cuadro 1). Debido a que el venado cola blanca es selectivo en su alimento, las arbustivas son fundamentales en su hábitat. Por ejemplo, en Texas, EUA, el incremento en las poblaciones de venado cola blanca en el siglo XX se atribuyó al aumento en la disponibilidad de arbustos (Taylor y Hahn, 1947). Además, se ha documentado que árboles y arbustos representan la mayor parte de la dieta del venado (Ramírez et al., 1996; Ramírez, 2004; Arceo et al., 2005). La alta disponibilidad de ese tipo de plantas en primavera y verano indica que durante la época de parto (julio y agosto), los venados tendrán suficientes existencias de arbustivas para su consumo.
Los pastos predominaron durante el otoño y el invierno (59.94 % y 53.58 %, respectivamente). Lo anterior es significativo, ya que los manchones de pastos aportan cobertura térmica en los meses con bajas temperaturas. En este contexto, se constató que el venado utilizó esas formaciones vegetales, constituidas por Cenchrus ciliaris L., con una altura promedio de 50 cm (±13.41), como sitios de descanso y cobertura, principalmente en invierno. Ramírez (2004) consigna que los venados consumen gramíneas en ausencia de sus especies forrajeras preferidas.
En relación al IVI, se identificaron 19 especies dominantes (Figura 2). Con excepción de Agave lechuguilla, Agave scabra, Euphorbia antisyphilitica Zucc., Flourensia cernua DC., Guaiacum angustifolium Engelm., Lippia graveolens, Parthenium hysterophorus y Paspalum notatum Flüggé todas han sido registradas en la dieta del venado cola blanca en el noreste de México (Ramírez, 2004; Fulbright y Ortega, 2006; Olguín et al., 2017). En cambio, evitan consumir Euphorbia antisyphilitica, debido a su alto contenido de cera que dificulta su digestión (Ramírez, 2004).
Además de ser una planta consumida por el venado, Guaiacum angustifolium aporta cobertura de escape (Ramírez, 2004). Aunque no se identificó como un taxón dominante, Karwinskia humboldtiana aporta cobertura de escape, especialmente, en el invierno, porque permanece verde (Taylor et al., 1997). Además, los venados prefieren como sitios de alimentación a los espacios abiertos casi sin arbustos, en donde predominan los pastos y las herbáceas (Lippia graveolens, Paspalum notatum y Parthenium hysterophorus) (Stewart et al., 2000).Scarnecchia et al. (1988) indican que la existencia en el hábitat de esos sitios es fundamental para el consumo nocturno.
La diversidad de plantas es un importante componente del hábitat para la nutrición del venado cola blanca (Fulbright y Ortega, 2006); pues cuando es alta, propicia mayor estabilidad y flexibilidad del sistema para tolerar fenómenos tales como la sequía, y ofrece una dieta de valor nutricional superior para los animales (Ramírez, 2004). El índice de Diversidad de Shannon indica que la mayor diversidad de plantas se presentó en primavera (2.45) y la menor en invierno (1.87). Con excepción de las primera dos estaciones del año, se identificó una diferencia significativa (p≤0.05) en la diversidad (Cuadro 2).
Cuadro 2 Valores del índice de Diversidad de Shannon (H’) y nivel de significancia de la prueba t-student en la diversidad de plantas por estación en la UMA Rancho San Juan, Monclova, Coahuila, México.
| Estación | H’ | Nivel de significancia (p≤0.05) de la prueba t-student en la diversidad | |||
|---|---|---|---|---|---|
| Primavera | Verano | Otoño | Invierno | ||
| Primavera | 2.42 | NS | 0.036* | 3.354*e-11*** | |
| Verano | 2.42 | 0.005** | 1.442*e-15*** | ||
| Otoño | 2.25 | 2.457*e-8*** | |||
| Invierno | 1.86 | ||||
Nivel de significancia: ≤0.1*; ≤0.01**; ≤0.001***; NS = No significativo.
Una diversidad de plantas relativamente alta en primavera es fundamental para la buena nutrición del venado, ya que esta es más relevante que la abundancia de las plantas que prefieren, puesto que una sola especie vegetal no cubre todos los requerimientos nutricionales del venado durante el año (Fulbright y Ortega, 2006; Aguiar et al., 2011). Un estudio realizado en Minnesota, EUA demostró que los venados cola blanca con una dieta más diversa mantienen un buen nivel nutricional de forma constante (Del Giudice et al., 1989).
Desde el punto de vista nutricional, se identificaron taxones arbustivos que destacan por su alto contenido de calcio (básico para el desarrollo de las astas), como Castela texana. En contraste, a pesar de su relativamente bajo contenido de este mineral, los pastos nativos del noreste de México satisfacen las necesidades metabólicas de calcio del venado cola blanca texano (Ramírez et al., 1996); por ejemplo, Aristida adscensionis L., Bouteloua gracilis (Kunth) Lag. ex Griffiths, Bouteloua curtipendula (Michx.) Torr. e Hilaria mutica (Buckley) Benth. Otras con alto contenido de vitamina K (necesario para la coagulación sanguínea) como Diospyros texana Scheele y Jatropha dioica Sessé ex Cerv. (Ramírez, 2004), se registraron en este estudio.
Producción de biomasa y su importancia en el manejo del venado cola blanca
Se estimó una producción de biomasa promedio de 621.20 (±85.08) kg ha-1 por estación. En el verano y el otoño el aporte fue mayor (744.36 ±44.20 kg ha-1 y 607.93 ±57.77 kg ha-1, respectivamente); mientras que, en invierno se estimó la menor producción (553.36 ±50.12 kg ha-1) (Figura 3). Esta producción es relativamente baja, en comparación con los 1 501 (± 492.35) kg ha-1 por estación calculados por Olguín et al. (2017) en el estado de Tamaulipas, y los 929.2 (±401.64) kg ha-1 citados por Navarro et al. (2018) en Zacatecas.

Figura 3 Variación en la producción de biomasa estacional por estrato vegetal en la UMA Rancho San Juan, Monclova, Coahuila, México.
La biomasa del hábitat es un factor fundamental para el desarrollo de los venados. Por ello, la baja producción influye directamente en la capacidad de carga del matorral. Está documentado que un individuo adulto requiere de un consumo diario de biomasa de 2 a 4 % de su peso corporal (Kie et al., 1983; Ramírez, 2004; Fulbright y Ortega, 2006). Sin embargo, el requerimiento varía en función del estado fisiológico, edad del venado, valor nutricional de las plantas disponibles, composición de las especies forrajeras y la distribución espacial del forraje (Stocker y Gilbert, 1977). Kie et al. (1983) señalan que el peso corporal de los machos adultos se incrementa rápidamente durante la primavera, y, por lo tanto, aumenta el consumo de forraje.
Las variaciones estacionales en la producción de biomasa en el presente estudio coinciden con las del consumo estacional de forraje por los venados del el sur de Texas. Se observa un decremento en la ingesta de forraje en verano, seguido de un incremento en otoño y en invierno el máximo decremento (Wheaton y Brown, 1983). El estrato medio aportó la mayor cantidad de biomasa en las cuatro estaciones del año (> 1 000 kg ha-1) y fue el verano, la estación con la más alta producción de biomasa (1 858.52 kg ha-1) (Figura 3). Lo anterior, se debió principalmente a que en julio se registraron en el área 80 mm de precipitación. Sin embargo, se estimó que los pastos y las herbáceas produjeron mayor cantidad en invierno (1 032.70 kg ha-1). El estrato alto fue el que menor biomasa aportó (≤ 250 kg ha-1) (Figura 3).
El venado cola blanca prefiere alimentarse de hojas y tallos jóvenes de arbustos, con más contenido de proteína y relativamente bajo en fibra y lignina (Ramírez, 2004). En este sentido, el matorral brinda los mayores valores de biomasa de arbustos en el año, en especial durante el verano (Figura 3), época cuando los venados requieren más forraje para producir el alimento de los cervatos (Fulbright y Ortega, 2006).
Por otro lado, la producción de herbáceas y gramíneas es necesaria, ya que el venado las consume si existe competencia por alimento (Ramírez, 2004). Como se consignó anteriormente, Acacia berlandieri y Acacia rigidula son plantas preferidas por el venado, y aunque fueron identificadas en este estudio como parte del matorral, su producción promedio de biomasa estacional (18.79 ±15.13 kg ha-1), en relación al peso promedio de biomasa por estación (621 ±85.08 kg ha-1) no fue alto (< 5.5 %). Por el contrario, en los pastos con alto contenido de lignina y bajo porcentaje de digestibilidad se verificó una mayor producción promedio de biomasa (187.11 ±45.28 kg ha-1) por estación (Figura 4), y hasta 43.84 % de la producción de biomasa en el invierno.
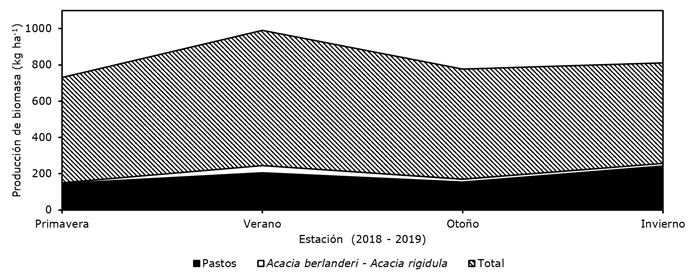
Figura 4 Comparación de la producción estacional de biomasa de especies básicas en la dieta del venado cola blanca en el noreste de México (Acacia berlandieri Benth. y Acacia rigidula Benth.) y pastos, en relación a la productividad total de biomasa, en la UMA Rancho San Juan, Monclova, Coahuila, México.
La relativa baja producción de biomasa de especies preferidas como forraje del venado en el área de estudio (Figura 4) se refleja en una baja capacidad de carga del matorral. Los resultados que aquí se exponen deben complementarse con un estudio sobre hábitos alimentarios del venado cola blanca en las cuatro estaciones del año. Aunque el matorral proporciona taxones forrajeros y de cobertura térmica, la dominancia de pastos como Cenchrus ciliaris puede ser un indicador de disturbio.
Además, es relevante que se continúen realizando estudios de diversidad de plantas en otros hábitats del venado cola blanca en el noreste de México, con el fin de tener un punto de referencia y establecer mejoras de la cobertura vegetal que favorezcan la diversidad de plantas en el matorral.
Conclusiones
Se identificaron 21 familias y 42 especies de plantas dentro del matorral. De ellas, 25 están registradas en la literatura como especies de las que se puede alimentar el venado cola blanca. Destacan Acacia berlandieri, Acacia rigidula y Opuntia engelmannii, como especies de su preferencia. El matorral reúne taxa que brindan cobertura térmica y de escape como Cenchrus ciliaris, Guaiacum angustifolium, Karwinskia humboldtiana y Yucca filifera.
Los taxones forrajeros no presentan una alta producción de biomasa, en relación a la producción de pastos, que son poco consumidos por el venado.
La producción de biomasa promedio por estación es, relativamente, menor que en otros sitios del noreste de México. Aunque, en el matorral existen especies que brindan alimento, cobertura térmica y de escape su productividad podría no mantener una población viable de venados a largo plazo.
La información que aquí se expone es útil para estimar, de forma más precisa, la capacidad de carga del matorral e identificar sitios para la repoblación del venado cola blanca; para ello, se recomienda complementar estos resultados con un estudio estacional sobre la dieta del venado cola blanca.

