











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El yodo (I) es un elemento no esencial para las plantas; sin embargo, se ha demostrado que la aplicación de dicho elemento genera efectos positivos en el crecimiento de diversas plantas (Medrano-Macías et al., 2016). Pequeñas cantidades de este elemento, en el rango de concentraciones de microelementos requeridas por las plantas, han mejorado el crecimiento, desarrollo y productividad de estas (Cakmak et al., 2017; Lyons, 2018; Duborská et al., 2020). Dicho impacto positivo se ha observado desde la modulación en la expresión génica hasta ser parte estructural de varias proteínas (Kiferle et al., 2021), así como el incremento en la concentración de minerales esenciales en las hojas (Cortés-Flores et al., 2016; Sularz et al., 2020).
De igual manera, las aplicaciones de I han mostrado efectos diversos sobre la capacidad antioxidante en varias especies; dicha capacidad antioxidante varía dependiendo de las fuentes, la concentración y el tipo de aplicación (Halka et al., 2020; Sabatino et al., 2021). Contrario a lo anterior, en algunas especies altos niveles de I en la solución nutritiva han provocado una marcada reducción de la expansión de la hoja y en la actividad fotosintética, lo cual fue atribuido a una gran acumulación de este elemento en los tejidos foliares (Incrocci et al., 2020).
Por otro lado, bajas concentraciones de I han mejorado la respuesta al estrés por salinidad, particularmente ocasionada por cloruro de sodio (NaCl). El efecto de la aplicación de I disminuyó la concentración de iones tóxicos como el Na+ y el Cl-; además de aumentar la concentración de azúcares solubles (sacarosa, glucosa y fructosa) que participan como osmoprotectores durante el ajuste osmótico, así como provocar la activación de enzimas antioxidantes como superóxido dismutasa (SOD) y ascorbato peroxidasa (APX), y el sistema antioxidante no enzimático en el que se encuentran los compuestos fenólicos, con el fin de mantener las especies reactivas de oxígeno (ROS), por sus siglas en inglés, como el O2 y H2O2 en niveles mínimos (Leyva et al., 2011; Gonzali et al., 2017; Kiferle et al., 2019).
Por estas razones, el I puede considerarse como un elemento beneficioso para ayudar a contrarrestar los efectos nocivos del estrés por salinidad, pero es necesario investigar de manera puntual el impacto en las diversas especies y estados de desarrollo (Leyva et al., 2011; Pérez-Salas y Medrano-Macías, 2021). Considerando lo antes mencionado, el objetivo de este experimento fue evaluar el impacto de la aplicación del yodo sobre la concentración de compuestos fenólicos, capacidad antioxidante y pigmentos fotosintéticos en hojas de tres cultivares de higuera sometidas a estrés salino.
Materiales y métodos
Ubicación del experimento y material vegetal
La investigación se llevó a cabo en el Centro de Agricultura Protegida de la Facultad de Agronomía de la Universidad Autónoma de Nuevo León, México, dentro de un invernadero tipo español. Las condiciones ambientales promedio dentro del invernadero fueron 25 °C de temperatura y 70% humedad relativa. Plantas de higuera de ocho meses de edad, con seis hojas cada una, fueron trasplantadas a bolsas de polietileno color negra de una capacidad de 10 L llenas de sustrato perlita. El experimento se estableció bajo un diseño experimental completamente al azar con arreglo factorial 3x2x2: tres cultivares de higuera (Ficus carica L.): Brown Turkey (BT), Kadota (K) y Black Mission (BM); dos niveles de NaCl (0 y 100 mmol L-1) y dos niveles de I (0 y 10 mg L-1) utilizando yodato de potasio (KI) como fuente de yodo. Ambos factores, NaCl y I fueron aplicados mediante riego gravitacional.
El agua base de riego tenía las siguientes características: Ca: 3.5 Mg, 1.8, Na: 0.9, HCO3: 2.5, Cl: 1.35, SO4: 2.35; HCO3: 2.5 meq L-1; pH: 7.5 y conductividad eléctrica (CE): 0.6 dS m-1. De la combinación de los factores antes mencionados surgieron 12 tratamientos con seis repeticiones cada uno. Cada planta fue considerada como una unidad experimental. El contenido relativo de clorofila, expresado en unidades SPAD, se midió utilizando el SPAD-502 portátil de Minolta (Konica Minolta, Osaka, Japón). Las unidades SPAD se midieron en el lóbulo medio de las hojas número 4, 5 y 6 de la base al ápice.
Los valores de dicha variable son el promedio de tres mediciones realizadas a los 5, 10 y 15 días después del inicio de los tratamientos. Asimismo, después de la última medición se colectaron muestras de cuatro plantas por tratamiento para la determinación de peso seco de hojas nuevas, compuestos antioxidantes y pigmentos. El peso seco se obtuvo mediante un horno de secado (Yamato modelo DX602C, Yamato Scientific America, Santa Clara, CA, USA) donde las muestras fueron sometidas a una temperatura de 60 oC durante tres días. El material resultante se pesó con balanza analítica con precisión 0.001 g.
Extracción de compuestos fenólicos
La extracción de compuestos fenólicos se realizó con base a lo reportado por Carballo-Méndez et al. (2019) con ligeras modificaciones. Cuatro gramos de tejido de hoja fresca fueron suspendidos en 40 ml de agua:metanol (20:80 v/v) (80%) y homogeneizó con una licuadora Oster (M4655-813/465-42, Sunbeam, Ciudad de México, México) en un vaso de cristal para papilla por 30 s. Después, se filtró con una tela de organza, colocó en tubos Corning de 50 ml y centrifugó a 4 500 rpm por 5 min a 25 °C. El sobrenadante se recuperó y almacenó protegido de la luz a -20 °C, hasta su posterior análisis.
Fenoles y flavonoides totales
El contenido de fenoles totales y flavonoides totales se evaluó con base a lo reportado por Rodríguez-Salinas et al. (2020), con las siguientes modificaciones: los fenoles totales, se evaluaron basado en el método colorimétrico de la reacción colorimétrica de Follin-Ciocalteu, con 200 µl del extracto metanólico en 2 600 µl de agua, seguido se agregaron 200 µl y alcalinizó agregando 2 000 µl de Na2CO3, después se dejó reaccionar por 90 min y midió absorbancia en un espectrofotómetro SP-830 Plus (Barnstead, Turner, EUA) a una longitud de onda de 750 nm.
La curva patrón se preparó de ácido gálico en concentraciones de 0 a 200 mg L-1, como blanco se usó metanol al 80%. Los resultados se expresaron en miligramos equivalentes de ácidos gálico por gramo de tejido fresco (mg EAG g-1). Los flavonoides totales, se midieron por el método del cloruro de aluminio (AlCl3) e hidróxido de sodio (NaOH), brevemente, fueron agregados 200 µl del extracto metanólico a 3 500 µl de agua, seguido de 150 µl de una solución al 5% de NaNO2 y dejó reposar por 5 min, después se agregó 150 µl de una solución al 10% de AlCl3 y finalmente 5 min después se agregó 1 000 µl de NaOH al 1 N.
Se dejó reaccionar durante 15 min y se midió la absorbancia de las muestras en un espectrofotómetro SP-830 Plus a la longitud de 510 nm. Las concentraciones se calcularon con base con una curva de catequina realizada en concentraciones de 0 a 200 mg L-1. Los resultados se expresaron en miligramos equivalentes de catequina por gramo de tejido fresco (mg EC g-1).
Capacidad antioxidante DPPH
La determinación de capacidad antioxidante por el método de 2,2-difenil-1-picrilhidrazilo (DPPH) de los extractos metanólicos del tejido de hoja fresca de higuera se realizó en base a lo descrito por Rodríguez-Salinas et al. (2020), con ligeras modificaciones que a continuación se describen: DPPH, se evaluó mediante la solución stock de trabajo preparada al 60 µM disuelto en metanol al 80%, ajustada a una absorbancia a 1 nm a una longitud de onda de 517 nm. La reacción se realizó agregando 0.51 µl del extracto metanólico a 1.5 ml de la solución stock, se dejó reposar por 30 min y medió absorbancia en un espectrofotómetro SP-830 Plus a una longitud de onda de 517 nm. Como curva patrón se utilizó Trolox en concentraciones de 0 a 500 µmol L-1. Los resultados se expresaron en Trolox equivalentes por cada gramo de tejido fresca (µmol ET g-1).
Se calculó la IC50 de DPPH para la cantidad de antioxidante necesaria para inhibir 50% de la oxidación del radical, se consideró la absorbancia ajustada a 1 en las soluciones de trabajo como 100% de oxidación y se utilizó metanol como control. Los resultados se expresaron en miligramos por gramo de tejido de hoja fresca (mg g-1).
Pigmentos fotosintéticos
Los pigmentos fotosintéticos clorofilas (a, b y totales) y carotenoides totales, se extrajeron y calcularon con base a la reportado por Rajput y Patil (2017), con ligeras modificaciones. Brevemente, en promedio 4 g de tejido fresco de hoja fueron homogeneizadas en una licuadora Oster con un vaso de cristal para papilla por 30 s en presencia de 40 ml de agua:acetona (20:80 v/v) (80%) que contenía 200 mg de Na2CO3. Después, el homogeneizado se pasó a través de una tela de organza, colocó en tubos Corning de 50 ml de capacidad y centrifugó a 6 000 rpm por 5 min a temperatura ambiente. El sobrenadante se recuperó y almacenó protegido de la luz a -20 °C, hasta su posterior análisis por espectrofotometría.
Las clorofilas y carotenoides totales se cuantificaron en un espectrofotómetro SP-830 Plus a longitudes de onda de 663-645 y 480-510 nm, respectivamente. Los resultados se expresaron en miligramos por gramo de tejido fresco (mg g-1).
Donde: A= absorbancia obtenida a la longitud de onda especifica; V= volumen de extracción; y PM= peso de la muestra.
Análisis estadístico
Los valores obtenidos fueron analizados con la prueba de Levene para homogeneidad de varianzas y con la prueba de Kolmogorov-Smirnov para la comprobación de la normalidad. Para el análisis de varianza de las variables fue considerada el covariable contenido relativo de clorofila, la cual fue evaluada un día antes del inicio de los tratamientos. En las variables donde se encontró diferencia significativa se realizó una comparación de medias mediante la prueba de Tukey (p< 0.05). Para dichos análisis se utilizó el paquete estadístico SPSS versión 22.0, IBM.
Resultados y discusión
El análisis de varianza mostró que los factores principales (cultivares, niveles de NaCl, niveles de I) y la interacción entre estos fue diferente en las variables evaluadas (Cuadro 1).
Cuadro 1 Valores de F y niveles de significación (P) observados en los análisis de varianza.
| Variables | Cultivar (C) | NaCl (S) | Iodo (I) | C*S | C*I | S*I | C*S*I | |
| Contenido relativo de clorofila | F | 14.271 | 0.022 | 17.485 | 2.021 | 1.992 | 3.896 | 7.734 |
| p | <0.001 | ns | <0.001 | ns | ns | ns | 0.003 | |
| Número de hojas nuevas | F | 4.878 | 9.936 | 7.288 | 0.798 | 1.153 | 4.851 | 0.165 |
| p | 0.017 | 0.004 | 0.013 | ns | ns | 0.038 | ns | |
| Peso seco de hojas nuevas | F | 0.425 | 16.245 | 15.845 | 0.37 | 0.496 | 4.567 | 0.099 |
| p | ns | 0.001 | 0.001 | ns | ns | 0.043 | ns | |
| Fenoles totales | F | 9.275 | 0.241 | 4.588 | 0.163 | 1.197 | 21.886 | 8.205 |
| p | 0.001 | ns | 0.043 | ns | ns | <0.001 | 0.002 | |
| Flavonoides totales | F | 65.69 | 0.891 | 0.611 | 0.573 | 1.556 | 2.435 | 1.821 |
| p | <0.001 | ns | ns | ns | ns | ns | ns | |
| DPPH | F | 4.497 | 3.566 | 3.501 | 0.846 | 1.027 | 2.312 | 4.028 |
| p | 0.022 | ns | ns | ns | ns | ns | 0.032 | |
| IC50 | F | 5.544 | 3.068 | 4.905 | 0.401 | 6.231 | 2.146 | 0.677 |
| p | 0.011 | ns | 0.037 | ns | 0.007 | ns | ns | |
| Clorofila a | F | 20.8 | 23.283 | 7.974 | 0.657 | 2.656 | 0.198 | 2.377 |
| p | <0.001 | <0.001 | 0.01 | ns | ns | ns | ns | |
| Clorofila b | F | 47.641 | 68.021 | 11.578 | 19.475 | 2.202 | 0.157 | 3.927 |
| p | <0.001 | <0.001 | 0.002 | <0.001 | ns | ns | 0.034 | |
| Carotenoides | F | 14.557 | 23.262 | 1.079 | 7.702 | 0.695 | 0.005 | 1.275 |
| p | <0.001 | <0.001 | ns | 0.003 | ns | ns | ns |
ns= no significativo.
Contenido relativo de clorofila y crecimiento
En las unidades Spad se mostró interacción triple entre cultivares, nivel de NaCl y presencia de I. En el cultivar BT el I redujo 3% las unidades Spad en condiciones de baja salinidad, mientras que en condición de salinidad las aumentó 6%. Para el cultivar K el comportamiento fue similar en ambas condiciones de NaCl, incrementando dichas unidades 11 y 3% en condiciones de baja y alta salinidad, respectivamente. En el cultivar BM, el efecto del I sólo se presentó en el nivel alto de salinidad, con un incremento de 13% (Figura 1A). Estos resultados encontrados en las hojas de higuera son similares a los reportados por Cortes et al. (2016), quienes registraron que la presencia de I incrementó hasta en 5% la concentración de nitrógeno relacionado con las unidades Spad, en hojas de pimiento morrón (Capsicum annum L.).
El número de hojas nuevas presentó diferencia entre los cultivares, siendo la BT la que presentó mayor cantidad. Para esta misma variable se mostró una interacción entre nivel de NaCl y I, donde la presencia de I incrementó 204% el número de hojas en la solución con NaCl (Figura 1B). El peso seco de hoja sólo mostró interacción entre nivel de NaCl y I, donde el I incrementó el peso seco de hoja 273% en la presencia de NaCl (Figura 1C). Al igual que nuestros resultados, Salimpour et al. (2019), reportó que diferentes cultivares e higuera muestran diferencias en el crecimiento de hojas, como consecuencia de la diversidad genética. Kiferle et al. (2019), reportó que en condiciones no salinas la presencia de I no incrementó la materia seca en albahaca (Ocimum basilicum L.), mientras que Leyva et al. (2011); Blasco et al. (2013) reportaron que en condiciones de salinidad la presencia de I incrementó la biomasa aérea de lechuga (Lactuca sativa L.), lo cual se le atribuyó a la reducción de Na y Cl y al aumento en la relación K/Na.

Figura 1 Efecto de los cultivares NaCl y I, el contenido relativo de clorofila (SPAD) (A); en el número de hojas nuevas (B); y peso seco de hojas nuevas (C). Las líneas sobre las barras representan el error estándar. Las letras diferentes en cada barra significan que los tratamientos fueron estadísticamente diferentes (Tukey, p< 0.05).
Compuestos fenólicos y capacidad antioxidante
La concentración de compuestos fenólicos fue afectada por la interacción de los diferentes factores. Los cultivares BR y K redujeron la concentración de fenoles totales en 26 y 45% en la ausencia de NaCl, mientras que con la presencia de NaCl estos compuestos se incrementaron en 25 y 17%, respectivamente. Contrario a lo anterior, en el cultivar BM la presencia de I en situación salina redujo los fenoles totales en 9% (Figura 2A). La concentración de flavonoides totales sólo fue afectada por los cultivares, donde el cultivar BT superó a K y BM, de manera general, 150% (Figura 2B). La actividad antioxidante por DPPH mostró interacción entre los tres factores. En el cultivar BT la presencia de I en la solución sin NaCl disminuyó 11% dicha actividad, mientras que en la presencia de NaCl el I la incrementó 455%. En el cultivar BM sin la presencia de NaCl, la presencia de I incremento esta actividad 22% (Figura 2C).
La IC50 sólo fue afectada por la interacción de los cultivares y el I, particularmente en el cultivar BM donde la presencia de este elemento redujo 21% dicha concentración (Figura 2D). El incremento en la capacidad antioxidante por el ensayo DPPH de las plantas revela un aumento en la síntesis de moléculas reductoras de bajo peso molecular, principalmente antioxidantes no enzimáticos (Pérez-Salas y Medrano-Macías, 2021) como los fenólicos (Leyva et al., 2011). Debido a que DPPH basa su fundamento en la capacidad de medir los compuestos con capacidad de ceder átomos de H (Schaich et al., 2015).
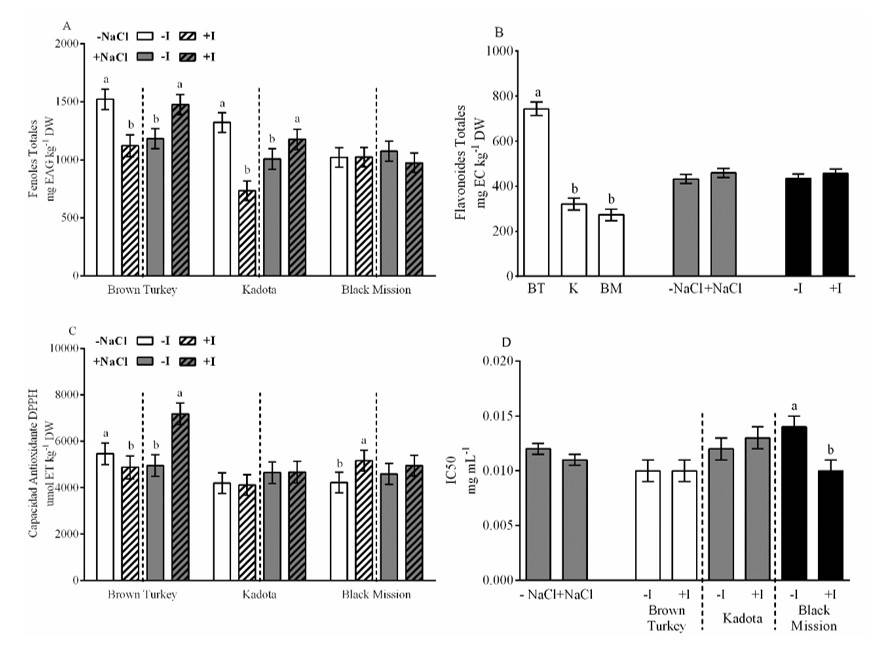
Figura 2 Efecto de los cultivares, NaCl y I en la concentración de fenoles totales (A); flavonoides totales (B); actividad antioxidante DPPH (C); y la concentración media inhibitoria (IC50) (D). Las líneas sobre las barras representan el error estándar. Las letras diferentes en cada barra significan que los tratamientos fueron estadísticamente diferentes (Tukey, p< 0.05).
Diversos autores han reportado que aplicaciones de I aumentan la capacidad antioxidante de los tejidos de las plantas. Kiferle et al. (2019), mencionan que el I incrementó los fenoles totales en hojas de albahaca, así como Blasco et al. (2013), quienes reportaron que, en lechuga sometida a estrés salino, el I incrementó la concentración de fenoles totales, sin encontrar diferencias en la concentración de flavonoides. Dichos resultados concuerdan con lo observado en este experimento. Contrario a lo anterior Golubkina et al. (2018), reportaron que en hojas de mostaza (Brassica juncea L.) la aplicación de I incrementó la concentración de flavonoides.
De igual manera Cortés-Flores et al. (2016), reportaron que la aplicación foliar de KI incrementó la actividad antioxidante DPPH en plántulas de pimiento en condiciones óptimas de desarrollo, mientras que Kiferle et al. (2019) reportaron la disminución de esta actividad en plantas de albahaca. En el presente experimento dicho efecto fue diferente entre los cultivares y nivel de salinidad. En condiciones no salinas la presencia de iodo disminuyó la actividad antioxidante DPPH en el cultivar BT, mientras que la incrementó en el cultivar BM. En situación salina la presencia de I sólo benefició al cultivar BT. Particularmente para la IC50, Incrocci et al. (2020) reportó que, en albahaca, la presencia de I redujo la IC50, aunque el impacto fue diferente entre cultivares, situación muy similar a lo mostrado en este experimento.
Pigmentos fotosintéticos
La concentración de clorofila a mostró efectos principales de los tres factores. Los cultivares BT y K, superaron a BM en 35%. La presencia de NaCl redujo estos compuestos 18%, mientras que la presencia de I los incrementó 12% (Figura 3A). La concentración de clorofila b fue afectada por la interacción de los tres factores. En el cultivar BT la presencia de I incrementó estos la concentración de clorofila b en 17% sin la presencia de NaCl y 83% en la presencia de dicha sal. En el cultivar K el efecto del I fue diferente dado que, en la ausencia de NaCl, la clorofila b se incrementó 36%, mientras que, en la presencia de la sal, estos compuestos se redujeron 17%.
Para el cultivar BM la presencia de I en condición de salinidad incrementó estos pigmentos 65% (Figura 3B). La concentración de carotenoides solo fue afectada por la interacción entre el cultivar y NaCl. En los cultivares BT y K, la presencia de NaCl redujo la concentración de carotenoides en 36 y 20%, respectivamente (Figura 3C). Los resultados antes mostrados difieren parcialmente con los reportados por Pérez-Salas y Medrano-Macías (2021), quienes señalan que en plantas de tomate (Solanum lycopersicum L.) sometidas a estrés salino, la presencia de I no modificó la concentración de clorofila a, pero si incrementó la concentración de clorofila b.

Figura 3 Efecto de los cultivares, NaCl y I en la concentración de clorofila a (A); clorofila b (B); y carotenoides totales (C). Las líneas sobre las barras representan el error estándar. Las letras diferentes en cada barra significan que los tratamientos fueron estadísticamente diferentes (Tukey, p< 0.05).
Conclusiones
La aplicación de 10 mg L-1 de yodo en plántulas de higuera incrementó, en términos generales, el contenido relativo de clorofila, número y peso seco de hojas nuevas, así como la actividad antioxidante y la concentración de clorofilas, en condición de estrés salino. Por lo anterior el yodo puede ser considerado como una alternativa para mitigar el estrés provocado por NaCl en Ficus carica L. Sin embargo, de manera particular, es necesario considerar que existen diferencias en el impacto del yodo entre los cultivares por lo que es preciso conocer el efecto de la aplicación de este elemento en especies y variedades de interés específico.

