










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de biodiversidad
versión On-line ISSN 2007-8706versión impresa ISSN 1870-3453
Rev. Mex. Biodiv. vol.84 no.1 México mar. 2013
https://doi.org/10.7550/rmb.26831
Ecología
Fitoplancton del embalse Yacyretá (Argentina-Paraguay) a una década de su llenado
Phytoplankton of the Yacyretá Reservoir (Argentina-Paraguay) after a decade from its filling
Norma Meichtry de Zaburlín1, Roberto Eugenio Vogler2,3, Victor Martín Llano1 e Irmgard Sandra Mabel Martens1*
1 Facultad de Ciencias Exactas Químicas y Naturales, Universidad Nacional de Misiones, Rivadavia 2370 (N3300LDX). Posadas. Misiones, Argentina. *ismartens@hotmail.com
2 División Zoología Invertebrados, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Paseo del Bosque s/n (B1900FWA), La Plata, Argentina.
3 Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
Recibido: 23 agosto 2011
Aceptado: 03 septiembre 2012
Resumen
Se estudió la estructura y dinámica del fitoplancton del embalse Yacyretá en relación con las principales variables limnológicas durante el periodo diciembre de 2004 a noviembre de 2005, a 10 años del llenado del embalse. Las muestras cualitativas y cuantitativas fueron recolectadas en 6 estaciones, previamente seleccionadas siguiendo un criterio de zonación longitudinal. Se analizó la densidad, composición, riqueza, diversidad específica, equitatividad y grupos taxonómicos dominantes. Durante el período de estudio, el embalse Yacyretá no presentó estratificación térmica y la concentración de oxígeno disuelto fue alta en toda la columna de agua. Se registraron 200 especies distribuidas en 9 clases. El número de especies por muestra osciló entre 14 y 45 taxa y la diversidad específica entre 1.55 y 4.61 bits. La densidad varió entre 66 y 10 060 ind/ml, con dominancia de Cryptophyceae y Bacillariophyceae en las distintas regiones del embalse. Al inicio del otoño y final de la primavera, se registró un aumento en la densidad de Cyanobacteria en las estaciones con mayor tiempo de residencia del agua. La composición y densidad del fitoplancton presentó una marcada variación espacial y temporal determinada principalmente, por las variaciones del régimen hidrosedimentológico del río Paraná.
Palabras clave: río Paraná, variables ambientales, florecimiento, variación espacio-temporal.
Abstract
In this study, we analyzed the structure and dynamics of the phytoplankton of the Yacyretá Reservoir and its relationship with the main limnological variables between December 2004 and November 2005, 10 years after the filling of the reservoir. Qualitative and quantitative samples were collected at 6 sampling stations, following a longitudinal zonation criterion. Density, composition, specific richness, diversity, equitability and dominance were analyzed. During the studied period, the Yacyretá Reservoir did not present thermal stratification, and the dissolved oxygen concentration in the water column was high. Two hundred species in 9 algae classes were recorded. The number of species per sample varied between 14 and 45 taxa and the specific diversity between 1.55 y 4.61 bits. Density ranged among 66 and 10 060 ind/ml, with dominance of Cryptophyceae and Bacillariophyceae in different regions of the reservoir. An increase in the Cyanobacteria density at the begining of Autumn and at the end of Spring, mainly in the stations with longer time of permanence of water, was recorded. During the analysis period the composition and density of the phytoplankton presented a noticeable spatial and temporal variation, determined mainly by the hydrosedimentologic cycle variations.
Key words: Paraná River, environmental variables, blooms, spatial-temporal variation.
Introducción
En América del Sur, las construcciones de embalses para generación de energía hidroeléctrica, abastecimiento humano e irrigación han aumentado notablemente en las últimas décadas (Nogueira et al., 2005; Meichtry de Zaburlín et al., 2010). El río Paraná, uno de los principales ríos de América del Sur y cuya cuenca concentra la mayor densidad poblacional del subcontinente, ha sido afectado durante el siglo XX por la construcción de más de una veintena de represas (COMIP, 1994). Estas intervenciones han modificado sustancialmente las características del sistema fluvial (Tundisi y Matsumura-Tundisi, 2003; Thomaz et al., 2007; Souza-Fhilo, 2009). A finales del siglo pasado (abril de 1990), se produjo una nueva alteración en la fisiografía del río Paraná en su recorrido entre Argentina y Paraguay, debido al cierre de la represa Yacyretá (Peso y Bechara, 1999), la cual constituye el último eslabón de una cadena de embalses construidos sobre este gran río. Como consecuencia, se registraron cambios notables en las comunidades acuáticas ubicadas aguas arriba de la presa a partir de agosto de 1994, cuando se alcanzó la cota 76 m snm (Garrido, 1999; Meichtry de Zaburlín, 1999; Peso y Bechara, 1999). Esta primera etapa del llenado del embalse, se caracterizó por un período rico en materia orgánica y nutrientes, con disminución del flujo del agua y aumento del tiempo de residencia, estableciéndose un gradiente longitudinal de las características limnológicas, influenciadas por el ciclo hidrológico y el manejo operativo de la represa. Ello produjo un importante impacto sobre las comunidades acuáticas existentes. Se observaron cambios en la composición del fitoplancton, en su densidad y riqueza de especies, vinculados principalmente a la disminución de la velocidad de la corriente y a la incorporación de nuevos y diversos hábitats. El número total de especies registradas durante el primer ciclo anual se incrementó en un 40% respecto a estudios previos al llenado del embalse (Meichtry de Zaburlín, 1999). Durante el primer año se registró un elevado número de especies por muestra ( = 41), altos valores de diversidad específica (= 4.25 bits), el incremento de la densidad del fitoplancton en la zona lacustre y cambios en la dominancia de especies, con aumento de Cryptophyceae (Meichtry de Zaburlín, 1999, 2002). Si bien son numerosos los trabajos que hacen referencia al fitoplancton de embalses en zonas tropicales y subtropicales, en Argentina los estudios de distribución espacial y temporal de esta comunidad son escasos (Quirós y Luchini, 1982; Mariazzi et al., 1991; Meichtry de Zaburlín, 2002; Meichtry de Zaburlín et al., 2010). El objetivo del trabajo fue describir la variación espacial y temporal del fitoplancton en relación a variables ambientales durante el periodo diciembre de 2004 a noviembre 2005 de un gran embalse subtropical, luego de transcurrir 10 años de permanencia en la cota inicial de 76 m snm, previsto en su cronograma de llenado. Se plantearon las siguientes hipótesis: i) la presencia de un ambiente más estable como resultado de mantenerse la cota del embalse durante una década con el consecuente proceso de envejecimiento, conduce a una menor fluctuación en la composición de especies, estabilidad biótica y variables ambientales, con respecto a los primeros años después del llenado del embalse; ii) siguiendo un patrón de zonación longitudinal, la comunidad fitoplanctónica en las distintas zonas del embalse difiere en sus atributos y estas diferencias pueden ser interpretadas en términos de características distintivas de cada zona, incluyendo tiempo de residencia y variación de la transparencia del agua y iii) la densidad y diversidad del fitoplancton es mayor durante periodos de estiajes, con mayor tiempo de residencia del agua en el embalse y cuando la heterogeneidad espacial se incrementa.
= 41), altos valores de diversidad específica (= 4.25 bits), el incremento de la densidad del fitoplancton en la zona lacustre y cambios en la dominancia de especies, con aumento de Cryptophyceae (Meichtry de Zaburlín, 1999, 2002). Si bien son numerosos los trabajos que hacen referencia al fitoplancton de embalses en zonas tropicales y subtropicales, en Argentina los estudios de distribución espacial y temporal de esta comunidad son escasos (Quirós y Luchini, 1982; Mariazzi et al., 1991; Meichtry de Zaburlín, 2002; Meichtry de Zaburlín et al., 2010). El objetivo del trabajo fue describir la variación espacial y temporal del fitoplancton en relación a variables ambientales durante el periodo diciembre de 2004 a noviembre 2005 de un gran embalse subtropical, luego de transcurrir 10 años de permanencia en la cota inicial de 76 m snm, previsto en su cronograma de llenado. Se plantearon las siguientes hipótesis: i) la presencia de un ambiente más estable como resultado de mantenerse la cota del embalse durante una década con el consecuente proceso de envejecimiento, conduce a una menor fluctuación en la composición de especies, estabilidad biótica y variables ambientales, con respecto a los primeros años después del llenado del embalse; ii) siguiendo un patrón de zonación longitudinal, la comunidad fitoplanctónica en las distintas zonas del embalse difiere en sus atributos y estas diferencias pueden ser interpretadas en términos de características distintivas de cada zona, incluyendo tiempo de residencia y variación de la transparencia del agua y iii) la densidad y diversidad del fitoplancton es mayor durante periodos de estiajes, con mayor tiempo de residencia del agua en el embalse y cuando la heterogeneidad espacial se incrementa.
Materiales y métodos
Área de estudio. La presa de Yacyretá se encuentra localizada al norte de la provincia de Corrientes, Argentina, sobre el río Paraná (27°28' S, 56°44' O), en el inicio de su tramo potámico. Es una represa de llanura, con nivel del agua relativamente estable y corto tiempo de residencia, menor a 15 días. El embalse tiene una superficie de 1 140 km2, un volumen de 7 000 hm3, profundidades medias entre 6-7 m y una máxima de 23 m, a la cota inicial de 76 m snm. Se ubica fitogeográficamente en la región Neotropical, dominio Amazónico, provincia Paranaense (Cabrera, 1971). El clima es subtropical, cálido y húmedo, sin estación seca con una temperatura media de 21° C. Las precipitaciones durante el período de estudio fueron de 1 740 mm anuales, registrándose los máximos en otoño (abril y junio) y los mínimos en invierno (julio, agosto) y verano (diciembre, enero, febrero).
Muestreo. Para el estudio de la comunidad fitoplanctónica se dividió el área utilizando el criterio de zonación longitudinal propuesto por Thorton et al. (1990) y Kalff (2002) en: zona de entrada, fluvial o lótica (I: E1); zona de transición, intermedia o río-embalse (II: E2); zona del embalse o lacustre (III: E3MI, E3C, E3MD); zona de salida, aguas abajo de la presa (IV: E4) (Fig. 1).
Los estudios se realizaron sobre material recolectado en las 6 estaciones citadas, con una frecuencia mensual o bimensual, durante el período comprendido entre diciembre de 2004 y noviembre de 2005. Las muestras cualitativas para el estudio sistemático se obtuvieron con red de plancton de 20 de abertura de malla y fueron fijadas con una concentración final de formol al 5%. Las muestras cuantitativas fueron obtenidas subsuperficialmente mediante un muestreador de tipo Van Dorn o recolectadas manualmente utilizando frascos de 250 ml de capacidad y fijadas in situ con solución de Lugol y ácido acético. El análisis cuantitativo se realizó siguiendo la técnica de Utermöhl (1958), con un microscopio invertido Olympus a 400x, utilizando cámaras de sedimentación de 10 ml. Los resultados se expresaron como individuos (células, cenobios, colonias o filamentos) por mililitro (ind/ml). Se contaron un mínimo de 100 individuos de la especie más frecuente, de modo que el error de recuento fuese inferior al 20% con un nivel de significación del 95% (Lund et al., 1958).
En cada sitio de muestreo fueron medidos in situ la transparencia del agua (disco de Secchi de 25 cm), temperatura, pH, conductividad eléctrica y oxígeno disuelto (analizador digital Cole-Palmer 5566-15). Los análisis de sólidos, color, turbidez, alcalinidad, clorofila-a y nutrientes (fósforo total, ortofosfato, nitrito, nitrato) fueron realizados de acuerdo a APHA (1992). Los datos de cota, caudal, tiempo de residencia del agua y variables físicas y químicas, fueron proporcionados por la Entidad Binacional Yacyretá (EBY) en el marco del Programa Calidad de Agua del Embalse Yacyretá (Convenio EBY-Universidad Nacional de Misiones).
Análisis de datos. Se analizó riqueza de especies (número de especies presentes en cada muestra cuantitativa), composición, densidad, grupos taxonómicos dominantes, diversidad específica (Shannon y Weaver, 1963) y equitatividad (Lloyd y Ghelardi, 1964) de la comunidad fitoplanctónica.
Para la comprobación de diferencias en los atributos del fitoplancton (riqueza, densidad, diversidad específica y equitatividad) y variables ambientales (temperatura, oxígeno disuelto, transparencia del agua, turbidez, pH, conductividad, fósforo total, ortofosfato, nitrito, nitrato, nitrógeno amoniacal, clorofila-α) entre estaciones de las diferentes zonas del embalse, se aplicó un análisis univariado de la varianza (ANOVA), realizando una prueba de Tukey a posteriori, para reconocer las diferencias significativas entre estaciones. Para el análisis de las relaciones entre las variables ambientales y atributos de la comunidad fitoplanctónica se utilizó la matriz de correlación de Pearson (STATISTICA ed. '98, StatSoft, Inc, Tulsa, OK, EUA).
Resultados
Características ambientales. El comportamiento del río Paraná registrado en el tramo lótico (zona de entrada) presentó 2 periodos de creciente, uno en verano (enero-febrero) y otro en primavera (octubre-noviembre). Los caudales mínimos se observaron al final del verano (marzo) y durante el invierno (julio-agosto) (Fig. 2). El nivel hidrométrico del río se correlacionó significativamente con el caudal (r= 0.968; p< 0.001) y con el nivel de cota del embalse (r= 0.396; p< 0.001). El ciclo estudiado fue estable con valores medios mensuales de cota de 75.80 m snm, inferiores al nivel previsto para esta etapa de 76 m snm y con un mayor tiempo de residencia del agua. Entre las variables más influenciadas por el pulso hidrológico se destacaron la transparencia del agua y la turbidez. En la zona lacustre se observaron los valores más altos de transparencia del agua con un máximo de 340 cm en la E3MI, asociados éstos a la magnitud de los pulsos de creciente. La transparencia del agua se correlacionó de manera inversa con el aumento del nivel del agua del embalse (r= -0.256; p< 0.05). Los valores de turbidez fueron generalmente bajos y los mínimos se registraron en las estaciones ubicadas en la margen izquierda del embalse y aguas abajo de la presa durante el estiaje, y los más elevados en la estación de entrada durante las crecientes con altos caudales (Figs. 3A, B). La turbidez se correlacionó inversamente con la transparencia del agua (r= -0.656; p< 0.001) y positivamente con el caudal (r= 0.443, p<0.001). La concentración de sólidos suspendidos fue más elevada en la zona de entrada durante los periodos de creciente y los mínimos durante los estiajes, especialmente en la zona lacustre. La concentración de fósforo total varió entre 10 y 36 μg/l y el ortofosfato entre 5 y 13 μg/l; fue más elevada en la estación de entrada y transición, y más baja en la estación de salida (Figs. 4A, B). Los valores más altos se registraron en periodos de crecientes con elevados caudales, hallándose una correlación significativa positiva entre ambas variables (r= 0.422, p< 0.001).
La temperatura del agua fluctuó entre 17.7° C (julio) y 30.6° C (enero) y presentó valores medios similares en las distintas estaciones (Cuadro 1). La variación temporal de la temperatura del agua dependió de la fluctuación de la temperatura del ambiente, se registraron diferencias de hasta 12° C entre el invierno (julio) y el verano (enero). Durante el período analizado, no se registró estratificación térmica en el cuerpo principal del embalse y la concentración de oxígeno disuelto fue alta en toda la columna de agua (Guillermo Gavilán, comunicación personal). La concentración de oxígeno disuelto se correlacionó positivamente con el caudal (r= 0.342;p< 0.01) y de manera inversa con la temperatura del agua (r= -0.599; p< 0.001). La conductividad eléctrica del agua fue baja con un máximo de 57.8 μS/cm y un mínimo de 51 μS/cm, presentó valores medios homogéneos en las distintas regiones del embalse, reflejando un escaso contenido de electrolitos característico para el tramo del río analizado (Figs. 5A, B).
Los valores medios anuales más bajos de clorofila-a, se registraron en la región fluvial y en la zona de transición, hallándose los más altos en el embalse. En el mes de noviembre se registró el valor más alto de 7.32μg/l, en la estación E3MI, coincidente con un aumento en la densidad del fitoplancton total. La concentración de clorofila-a se correlacionó significativa y positivamente con la densidad del fitoplancton (r= 0.954, p< 0.001) y con el pH del agua (r= 0.706, p< 0.001).
El ANOVA reveló diferencias de medias significativas (p< 0.05) tanto espacial (para las variables pH, concentración de nitrito y clorofila-a) como temporal (para las variables temperatura, oxígeno disuelto, pH, transparencia, turbidez, conductividad y fósforo total) entre las estaciones de muestreo (Cuadro 2).

Composición taxonómica delfitoplancton. Se identificaron 200 especies (Apéndice) distribuidas en 9 clases: 23 Cyanobacteria, 62 Chlorophyceae, 18 Zygophyceae, 1 Xanthophyceae, 3 Chrysophyceae, 74 Bacillariophyceae, 11 Cryptophyceae, 2 Dinophyceae y 6 Euglenophyceae. La mayoría de las especies de Cyanobacteria fueron de aparición esporádica; entre las especies más constantes se destacaron Chroococcus minutus (Kützing) Nágeli, Microcystis aeruginosa (Kützing) Kützing, Chroococcus limneticus Lemmerman, C. limneticus var. subsalsus Lemmerman y Chroococcus sp., las cuales fueron más frecuentes y abundantes en las zonas de remanso, principalmente en la E3MI, donde se registraron florecimientos de Microcystis aeruginosa durante periodos con altas temperaturas y estabilidad de la columna de agua como ocurrió en el mes de abril.
Las Chlorophyceae presentaron mayor diversidad de especies durante los meses más cálidos (noviembre a febrero). Los taxones más constantes, se presentaron en más del 70% de las muestras, Chlamydomonas spp., Sphaerocystis schroeteri Chodat, Monoraphidium arcuatum (Kros.) Hindák, Dictyosphaerium pulchellum Wood, Eudorina elegans Ehrenberg y Pandorina morum (Müller) Bory; las Volvocales fueron más frecuentes y abundantes en las riberas del embalse.
Las Bacillariophyceae estuvieron integradas por 14 formas céntricas y 60 pennadas. Las diatomeas céntricas más frecuentes fueron: Aulacoseira granulata (Ehrenberg) Simonsen, Aulacoseira sp., Cyclotella sp. y entre las pennadas Synedra ulna (Nitz.) Ehrenberg.
Las Cryptophyceae constituyeron un grupo constante en el embalse, presentes en más del 90% de las muestras, principalmente las especies Cryptomonas caudata Schiller, C. erosa Ehrenberg, Chroomonas acuta Uterm., Chroomonas sp. Las Xanthophyceae, Chrysophyceae, Dinophyceae y Euglenophyceae fueron de aparición más esporádica y en porcentajes menores, contribuyendo a formar una pequeña fracción del fitoplancton total.
Variación espacial y temporal. En la zona de entrada o fluvial la densidad del fitoplancton fue baja, varió entre 73 y 230 ind/ml, con un valor medio anual () de 135 ind/ml y un coeficiente de variación (cv) del 36% (Fig. 6A). La mayor densidad se registró a finales del invierno (agosto), con el río estabilizado en aguas bajas, alta transparencia, baja concentración de sólidos y turbidez, y la menor a fines del otoño (abril), con abundantes precipitaciones en los días previos al muestreo y baja transparencia del agua. Cryptophyceae fue dominante principalmente en el verano (febrero, marzo) y otoño (mayo), con porcentajes de composición que oscilaron entre el 24 y 55% en los distintos muéstreos ( = 39%; cv= 28%) (Fig. 7). El segundo grupo en abundancia fue Bacillariophyceae, con un porcentaje que varió entre el 15 y 64% ( = 32%; cv= 51%). Cyanobacteria, con un porcentaje de participación promedio del 15% fueron dominantes a mediados y fines de la primavera (octubre, noviembre) y subdominantes en verano (enero-febrero). Las especies más abundantes, en orden de importancia numérica, fueron: Chroomonas acuta, Cryptomonas caudata, Chroomonas sp., Aulacoseira sp., A. granulata, Cyclotella sp. Adicionalmente, Microcystis aeruginosa, Chroococcus minutus y C. limneticus, fueron también abundantes en el verano (enero-febrero) y primavera (octubre-noviembre).
En la zona de transición la densidad total del fitoplancton (Fig. 6A) fue similar a la observada en la zona fluvial, osciló entre 66 y 204 ind/ml (* = 127;cv= 41%).El grupo de las Cryptophyceae resultó dominante en el 70% de las muestras, con un porcentaje de participación medio anual del 42% (cv= 36%). Las Bacillariophyceae ( =33%; cv= 49%) fueron más importantes numéricamente en diciembre, abril y junio, siendo subdominantes en los muestreos restantes. Las Chlorophyceae y Cyanobacteria presentaron abundancias relativas más elevadas en verano y primavera, al igual que lo observado en la zona de entrada (Fig. 7).
En la zona de embalse la densidad algal fue más elevada, varió entre 72 y 10 060 ind/ml, con el valor medio anual más alto (= 2 092 ind/ml) en la margen izquierda del lago (E3MI) con mayor tiempo de residencia del agua (Fig. 6A). El valor máximo registrado en el mes de noviembre, coincidió con el aumento de dos especies oportunistas, de pequeño tamaño y desarrollo rápido: Chroomonas acuta y Cryptomonas caudata que alcanzaron el 63% del fitoplancton total. La clase Cryptophyceae fue dominante en el 86% de las muestras con mayores porcentajes de participación en las estaciones ubicadas en las riberas del embalse (Fig. 7). Bacillariophyceae fue de presencia constante en el lago, con abundancias relativas inferiores a las registradas en la zona fluvial y de transición. Las densidades más elevadas se observaron al final de la primavera (diciembre), donde Aulacoseira sp. fue dominante, especialmente en las estaciones con mayor velocidad de la corriente (E3MD y E3C). También se registró un incremento en la abundancia de este grupo en el mes de agosto y en ambos casos se asoció con un aumento de la mezcla en la columna de agua provocada por fuertes vientos y oleaje durante el muestreo (Nelson Pividori, comunicación personal). Chlorophyceae fueron más diversas y abundantes en el verano, destacándose por su constancia y mayor densidad Chlamydomonas sp., Sphaerocystis schroeteri, Monoraphidium arcuatum y Dictyosphaerium pulchellum. El grupo de las Cyanobacteria, representada principalmente por Microcystis aeruginosa, aumentó significativamente su densidad durante el verano y principios del otoño con un máximo del 49% en abril en la estación E3MI (54 ind/ml= 16 200 cél/ml= 300 células por colonia). Se encontró una correlación significativa entre la densidad del fitoplancton y la concentración de clorofila-a en todas las estaciones de la zona de embalse (r= 0.472; p< 0.001).
En la zona de salida, la densidad del fitoplancton fue más elevada que la registrada en la región fluvial, con un valor medio de 361 ind/ml y con un alto coeficiente de variación (cv= 72%), donde fueron dominantes las Cryptophyceae en la mayoría de los muestreos y subdominantes las Bacillariophyceae. Al igual que lo registrado en la zona del embalse las diatomeas fueron más importantes por su abundancia en los meses de agosto y diciembre (Figs. 6A, 7).
En las distintas zonas del embalse las Dinophyceae, Chrysophyceae, Zygophyceae y Euglenophyceae fueron también encontradas en las muestras cuantitativas, con una baja densidad de organismos, contribuyendo a formar una pequeña fracción del fitoplancton total.
El ANOVA mostró diferencias espaciales significativas en la densidad total del fitoplancton (F= 3.516; p< 0.01). La prueba de Tukey evidenció la ausencia de diferencias significativas entre la zona de entrada y transición, pero si de éstas con respecto a la estación E3MI del embalse (p< 0.05) (Cuadro 3). La diferencia de medias en la concentración de clorofila-α fue estadísticamente significativa (F= 4.183; p< 0.01), mostrando que la estación E3MI, con valores más elevados se diferenció claramente del resto de las estaciones que presentaron valores mas bajos (p< 0.05). Influencia de los factores ambientales sobre la densidad del fitoplancton. No se registraron correlaciones significativas entre la abundancia del fitoplancton y el nivel del agua, ni entre la abundancia y el caudal. La densidad del fitoplancton se correlacionó significativamente con la transparencia del agua en la zona de entrada (r= 0.746; p< 0.05) y de manera inversa con la concentración de sólidos suspendidos (r= -0.651; p< 0.05). La clorofila-a se asoció positivamente con la temperatura del agua (r= 0.266; p< 0.05) y el pH (r= 0.706; p< 0.001).
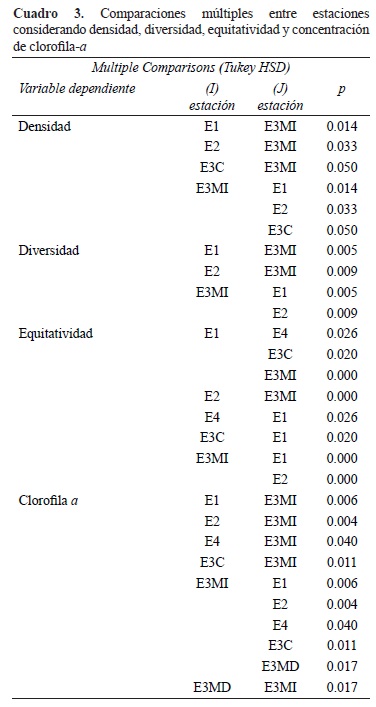
Riqueza de especies, diversidad específica y equitatividad. El número de especies por muestra varió entre 14 y 45 entidades (Fig. 6B). El valor medio anual más elevado (= 31 especies) se registró en la margen izquierda del embalse (E3MI) y los más bajos (= 25 especies) se presentaron en la zona fluvial y de transición. Para todas las estaciones, se registraron valores más altos en el verano y principios del otoño (entre diciembre y abril) y los más bajos en el invierno (junio a agosto). La riqueza de especies se correlacionó significativamente con la temperatura (r= 0.444; p< 0.001) y la transparencia del agua (r= 0.281; p< 0.05) y de manera inversa con la turbidez (r= -0.326; p< 0.05) y el nivel de cota del embalse (r= -0.297; p< 0.05), lo que implicó un aumento de la riqueza de especies en periodos de estabilidad del agua y durante los meses más cálidos.
El índice de diversidad específica varió entre 1.55 y 4.61 bits, con valores medios más altos en la zona de entrada y transición y el más bajo en la E3MI del embalse (Fig. 6C). El ANOVA evidenció diferencias significativas entre los valores del índice de diversidad específica entre fechas de muestreo (F= 2.992, p< 0.001) y estaciones (F= 4.443, p< 0.01); la prueba de Tukey separó claramente la estación del embalse E3MI del resto de las estaciones analizadas (p< 0.01).
La equitatividad varió entre 0.11 y 1 (Fig. 6D), con valores medios más elevados en la región de entrada y transición, y más bajos en las márgenes del embalse, principalmente en la E3MI. Las diferencias de medias entre fechas de muestreo y estaciones fueron significativas (F= 2.695;p< 0.01 y F= 7.545;p< 0.001, respectivamente). Al igual que lo observado para la diversidad, el test a posteriori discriminó claramente a la estación E3MI del resto de las estaciones (p< 0.05). La equitatividad se correlacionó de manera inversa con la transparencia del agua (r= -0.406; p< 0.01), y positivamente con la turbidez (r= 0.375; p< 0.01) lo que indicó que en periodos de aguas altas las especies presentaron una distribución más uniforme en las distintas secciones del embalse.
Discusión
La represa Yacyretá presenta características de sistema polimíctico con mezcla continua de la columna de agua y homogénea distribución de la temperatura y oxígeno disuelto debido a la alta tasa de renovación, bajos valores de conductividad y moderada transparencia, que la convierten en un sistema singular.
En este estudio la comunidad fitoplanctónica presentó una composición taxonómica similar a la registrada en los primeros años del llenado del embalse (Meichtry de Zaburlín, 1999; Meichtry de Zaburlín et al., 2010), y comparable a otros embalses de Brasil y Venezuela, donde las clases más abundantes en especies fueron las Bacillariophyceae y Chlorophyceae (Gonzalez et al., 1991; Henry y Nogueira, 1999). También se detectó una elevada riqueza de especies en comparación con otros tramos del Alto Paraná influenciados por la presencia de presas (Rodrigues et al., 2009), embalses argentinos más pequeños de la región noroeste (Tracanna et al., 1996), otros de la región central de tamaño similar a Yacyretá (Quirós y Luchini, 1982; Mariazzi et al., 1991) y embalses uruguayos y brasileños de diferentes tamaños (Gorja et al., 2002; Train et al., 2005). En cambio, el número de especies por muestra manifestó una notable disminución en relación a la registrada durante el primer año del llenado del embalse (= 41 especies) (Meichtry de Zaburlín, 1999).
En la región de entrada y transición, el fitoplancton mostró características lóticas, con una comunidad dominante alternativa entre Cryptophyceae y Bacillariophyceae. En contraste, en la zona lacustre con características lénticas, se registró la dominancia de pequeños fitoflagelados nanoplanctónicos (entre 10 y 25 um) de la clase Cryptophyceae, que se convirtió en el grupo mejor adaptado a las condiciones de embalse. La disminución de la densidad de Bacillariophyceae en las distintas zonas se observó desde el llenado de la represa, coincidente con la disminución de la velocidad de la corriente del agua (Meichtry de Zaburlín, 1999, 2002) y se acentúo durante este ciclo anual. Estas observaciones coinciden además con lo registrado en el Paraná Superior por Rodrigues et al. (2009), quienes observaron una disminución de las diatomeas céntricas en ese tramo del río, después del llenado del embalse de Porto Primavera en el 2001, que provocó una disminución de la descarga y de la concentración de sólidos suspendidos aguas abajo. Esto pone en evidencia la dependencia de estas algas de altos coeficientes de mezcla por turbulencia para mantenerse en suspensión.
La dominancia de Cryptophyceae también fue observada por diversos autores para embalses con alta tasa de renovación del agua en ecosistemas tropicales de Sudamérica, los que se caracterizan por la dominancia del plancton de pequeño tamaño con gran importancia de las Cryptomonadales (e.g. Margalef, 1978; Lewis y Riehl, 1982; Tundisi, 1990, 1999; Calijuri et al., 1999). Este grupo, que incluye algas de pequeño tamaño, con alta relación superficie/volumen, buenas competidoras, y tolerantes a una amplia variedad de condiciones ambientales y nutricionales, se vio favorecido por las nuevas condiciones generadas por el represamiento, principalmente por la reducción de la velocidad de la corriente. Las mismas, de acuerdo a la terminología de Reynolds (1997), pueden considerarse como organismos con rasgos morfológicos y fisiológicos intermedios entre dos estrategias primarias (R-aclimatable y C- invasora).
La dominancia de Aulacoseira sp. en Yacyretá, estuvo asociada a periodos de mayor turbulencia provocados por la acción del viento, lo que probablemente favoreció su resuspensión y permanencia en la zona fótica, al igual que lo observado en otros embalses de Brasil (Nogueira y Matsumura-Tundisi, 1996; Rodrigues et al., 2009). Su dominancia en diferentes embalses y ríos de distintas regiones geográficas de América del Sur, fue atribuida al hecho que se trata de un alga típico R estratega (Reynolds, 1997) tolerante a altas turbulencias y frecuentes cambios en las condiciones ambientales (De León y Chalar, 2003, Zalocar de Domitrovic et al., 2007).
Durante el período analizado, a diferencia de lo registrado en estudios previos (Meichtry de Zaburlín, 1999; Meichtry de Zaburlín et al., 2010), se observó el primer florecimiento de Microcystis aeruginosa que afectó a todo el embalse durante el verano y principios del otoño, y el cual se extendió al canal principal del río Paraná, aguas abajo de la presa (Zalocar de Domitrovic et al., 2007). Este incremento en la densidad de Cyanobacteria puede atribuirse a un prolongado periodo de estabilidad de las condiciones ambientales, como el nivel del agua en el lago, mayor tiempo de residencia, escasas precipitaciones y altas temperaturas. Además, la presencia de Cyanobacteria en los tramos fluviales, aguas arriba de Yacyretá, constituyó una fuente de aporte importante de organismos al embalse, donde el aumento en su densidad y permanencia indica que el sistema potenció las condiciones para su desarrollo. En los últimos años, también otros autores registraron aumentos en la abundancia de este grupo en el río Paraná, en el Uruguay y en zonas costeras de Argentina y Uruguay (Chalar et al., 2002; Zalocar de Domitrovic y Forastier, 2005; Rodrigues et al., 2009). Estos florecimientos, principalmente de Microcystis aeruginosa, provocaron un fuerte impacto sobre la opinión pública, por la presencia de sabor y olor desagradable en el agua utilizada para consumo humano y por la pérdida del valor recreacional del ecosistema acuático.
Los límites entre las zonas del embalse Yacyretá estuvieron condicionados por los eventos hidrológicos; así, en aguas altas, el embalse se comportó íntegramente como un río, con una distribución más homogénea del fitoplancton, mientras que en aguas bajas existió una mayor heterogeneidad en la composición y densidad. Cabe destacar que la estación de la margen izquierda del lago presentó condiciones más propicias para el desarrollo del fitoplancton donde se registró un mayor tiempo de residencia del agua.
Durante el período de estudio se encontraron diferencias significativas en la densidad, riqueza y diversidad del fitoplancton en sentido longitudinal y transversal, y en relación a las estaciones del año. La mayor estabilidad del embalse, en tiempos de bajos caudales favoreció al aumento del fitoplancton en las áreas más remansadas. En éstas, se verificó un incremento de la densidad media de hasta 15 veces, respecto de las otras zonas. Contrariamente, los caudales altos aumentaron el transporte y exportación de los organismos aguas abajo homogeneizando la comunidad fitoplanctónica. Según Tundisi et al. (1999), los pulsos en el flujo de entrada ejercen un fuerte impacto sobre las comunidades, pudiendo actuar como un disturbio intermedio que permitiría la coexistencia de algas de pequeño y gran tamaño.
En el embalse de Salto Grande, Argentina, Quirós y Luchini (1982) y Chalar et al. (2002) señalaron la importancia del régimen hídrico en la regulación de la comunidad fitoplanctónica. En atención a lo antes mencionado, los resultados obtenidos confirmaron la importancia del ciclo hidrológico y las variables directamente asociadas como modeladores de los cambios en la composición y abundancia del fitoplancton en Yacyretá, debido al bajo tiempo de residencia del agua.
Lo antes mencionado favoreció el incremento de Cryptophyceae, constituyendo el grupo más persistente y abundante en la etapa de estabilización del embalse. En general, las Cryptophyceae están asociadas a bajas condiciones de luz y a una mayor disponibilidad de nutrientes. En el caso de Yacyretá, este grupo de algas se destacó por su abundancia tanto en los primeros años después del llenado, con mayor turbidez y baja transparencia, como en esta etapa de estabilización donde se registró un importante aumento de la transparencia del agua y disminución de la concentración de nutrientes.
Durante el período de estudio, la densidad del fitoplancton del embalse Yacyretá disminuyó con el aumento del nivel del agua y el caudal por el efecto de dilución, y la riqueza se incrementó en los meses más cálidos. Los nutrientes, importantes para el desarrollo de la comunidad, como el fósforo total y ortofosfato mostraron una fuerte disminución respecto a los hallados en los primeros años después del llenado del embalse (Meichtry de Zaburlín, 2002); aunque las concentraciones en las distintas zonas del mismo serían suficientes para el mantenimiento de la comunidad algal existente, según los valores citados por Gorja et al. (2002).
De la comparación con estudios previos (Meichtry de Zaburlín, 1999, 2002; Meichtry de Zaburlín et al., 2010), a 10 años del represamiento se pudo constatar que algunas variables tendieron a aumentar (Cuadro 4, transparencia y conductividad eléctrica del agua) o a disminuir (turbidez, fósforo total y ortofosfato, nitrato), mientras que otras, se mantuvieron más o menos estables dentro de una escala de variación anual (pH, oxígeno disuelto). La dominancia de Cryptophyceae de pequeño tamaño persiste en esta fase de estabilización del embalse a la cota inicial de 76 m snm, con un incremento en su abundancia relativa. Estas observaciones concuerdan con las realizadas por Rodrigues et al. (2009) en el río Paraná Superior después del llenado del embalse de Porto Primavera, donde se observó una notable disminución de la descarga, concentración de sólidos suspendidos, fósforo total y un incremento de la transparencia del agua, como así también un aumento de la abundancia de Cyanobacteria y decrecimiento de Bacillariophyceae.
Las hipótesis formuladas fueron aceptadas teniendo en cuenta los resultados obtenidos, donde el mantenimiento del embalse a la cota inicial de 76 m snm por más de 10 años, permitió una mayor estabilidad en la estructura del fitoplancton. Sin embargo, los cambios en el nivel del agua del río (caudal) y las variables ambientales asociadas (tiempo de residencia, transparencia del agua) permitieron distinguir diferencias entre las distintas regiones del embalse, con un incremento de la densidad durante periodos de estiaje en las zonas con mayor tiempo de residencia del agua.
Actualmente y finalizado el cronograma del llenado del embalse Yacyretá, el cual alcanzó su cota máxima definitiva de 83 m snm (febrero de 2011), se continúan observando cambios en la comunidad fitoplanctónica, no sólo debido a las modificaciones del tiempo de residencia del agua y la incorporación de nuevas zonas inundables en el propio embalse, sino también como consecuencia de los procesos operacionales de las presas ubicadas aguas arriba en el Paraná Superior, las cuales podrían considerarse como un factor adicional en la explicación de la variabilidad del fitoplancton del embalse Yacyretá.
Agradecimientos
A la Entidad Binacional Yacyretá por el aporte financiero para la realización del presente trabajo y por proporcionar los datos ambientales. A Nelson Pividori por las labores de monitoreo. A la Dra. Juana Peso por sus valiosos aportes. A los evaluadores anónimos que contribuyeron con sus sugerencias a mejorar sustancialmente la primera versión del manuscrito.
Literatura citada
APHA (American Public Health Association). 1992. Standard methods for the examination of water and wastewaters, 18th ed. A. E. Greengerg. L. S. Clesceri y A. D. Eaton (eds.). American. Public Health Association, Washington. 815 p. [ Links ]
Cabrera, A. L. 1971. Fitogeografia de la República Argentina. Boletín de la Sociedad Argentina de Botánica 14:1-42. [ Links ]
Calijuri, M. C., G. L. B. Deberd y R. T. Minoti. 1999. Produtividade Primária pelo fitoplâncton na Represa de Salto Grande. En Ecologia de Reservatorios: Estrutura, Função e Aspectos Sociais, R. Henry (eds.). FAPESP-FUNDIBIO. Botucatu, Brasil. p. 109-148. [ Links ]
COMIP (Comisión Mixta Paraguayo-Argentina del Río Paraná). 1994. La fauna íctica del Río Paraná. Comisión mixta Paraguayo-Argentina del Río Paraná. Argentina. 255 p. [ Links ]
Chalar, G., L. De León, E. Brugnoli, J. Clemente y M. Paradiso. 2002. Antecedentes y nuevos aportes al conocimiento de la estructura y dinámica del Embalse de Salto Grande. In El agua en Sudamérica: de la Limnología a la Gestión en Sudamérica, A. Fernández-Cirelli y G. Chalar (eds.). CYTED, Buenos Aires. p. 123-142. [ Links ]
De León, L. y G. Chalar. 2003. Abundancia y diversidad del fitoplancton en el embalse de Salto Grande (Argentina-Uruguay). Ciclo estacional y distribución espacial. Limnetica 22:103-113. [ Links ]
Garrido, G. G. 1999. Composición y abundancia del zooplancton en dos estaciones de muestreo del embalse Yacyretá (Argentina), en las primeras etapas después del llenado a cota 76 msnm. Revista de Ictiología 7:27-35. [ Links ]
Gonzalez, E., J. Paolini y A. Infante. 1991. Water chemistry, physical features and primary production of phytoplankton in a tropical blackwater reservoir (Embalse de Guri, Venezuela). Verhandlungen des Internationalen Verein Limnologie. 24:1477-1481. [ Links ]
Gorja, J., M. Paradiso, L. De León, E. Brignoli, M. Mandiá y D. Conde. 2002. Problemática de la calidad de agua en el Sistema de Grandes Embalses del Río Negro. Revista Comisión de Integración Energética Regional (CIER) [ Links ]
Henry, R. y M. G. Nogueira. 1999. A Represa de Jurumirim (Sao Paulo). Primeira Síntese sobre o conhecimento Limnológico. In Ecologia de Reservatórios, R. Henry (eds.). Fundação de Aparo à Pesquisa do Estado de São Paulo-Fundação do Instituto de Biociências (FAPESP-FUNDIBIO), Botucatu, Brasil. p. 651-686. [ Links ]
Kalf, J. 2002. Limnology. Prentice Hall, New Jersey. 592 p. [ Links ]
Lewis, W. M. y W. Riehl. 1982. Phytoplankton composition and morphology in Lake Valencia, Venezuela. International Review of Hydrobiology 67:297-322. [ Links ]
Lloyd, M. y R. J. Ghelardi. 1964. A table for calculating the "equitability" component of species diversity. Journal of Animal Ecology 33:217-225. [ Links ]
Lund, J. W. G., C. Kipling y E. D. Le Cren. 1958. The inverted microscope method of estimation by counting. Hydrobiologia 11:143-170. [ Links ]
Margalef, R. 1978. Life forms of phytoplankton as survival alternatives in an unstable environment. Oceanologica Acta 1:493-509. [ Links ]
Mariazzi, A., V. Conzonno, R. Echenique y H. Labollita. 1991. Physical and chemical characters, phytoplankton and primary production of Ezequiel Ramos Mexía Reservoir (Argentina). Hydrobiologia 209:107-116. [ Links ]
Meichtry de Zaburlín, N. 1999. La comunidad fitoplanctónica durante las primeras etapas de llenado del embalse Yacyretá, Argentina. Revista de Ictiología 7:15-26. [ Links ]
Meichtry de Zaburlín, N. 2002. Estructura de la comunidad fitoplanctónica en el embalse Yacyretá, Argentina-Paraguay. Tesis, Universidad Nacional del Litoral. Santa Fe, Argentina. 128 p. [ Links ]
Meichtry de Zaburlín, N., J. G. Peso, G. G. Garrido y R. E. Vogler. 2010. Sucesión espacio-temporal del plancton y bentos en periodos posteriores al llenado del Embalse Yacyretá (Río Paraná, Argentina-Paraguay). Interciencia 35:897-904. [ Links ]
Nogueira, M. G. y T. Matsumura-Tundisi. 1996. Limnología de un sistema artificial raso (Represa de Monjolinho - São Carlos, SP). Dinâmica das populações planctónicas. Acta Limnologica Brasiliensia 8:149-168. [ Links ]
Nogueira, M. G., A. Jorcin, N. C. Vianna y Y. C. T. Britto. 2005. Reservatórios em cascata e os efeitos na limnologia e organização das comunidades bióticas (fitoplancton, zooplancton e zoobentos) um estudo de caso no rio Paranapanema. In Ecologia de reservatórios: impactos potenciais, ações de manejo e sistema em cascata, M. G. Nogueira, R. Henry y A. Jorcin (eds.). Rima, São Carlos. p. 83-125. [ Links ]
Peso, J. G. y J. A. Bechara. 1999. Estructura del zoobentos del embalse Yacyretá (Argentina), en dos estaciones de muestreo antes y después del llenado a cota 76 msnm. Revista de Ictiología 7:37-47. [ Links ]
Quirós, R. y L. Luchini. 1982. Características Limnológicas del Embalse de Salto Grande, III: Fitoplancton y su relación con parámetros ambientales. Revista de la Asociación de Ciencias Naturales del Litoral 13:49-66. [ Links ]
Reynolds, C. S. 1997. Vegetation processes in the pelagic: a model for ecosystem theory. Ecology Institute, Oldendorf-Luhe, Germany. 371 p. [ Links ]
Rodrigues, L. C., S. Train, V. M. Bovo-Scomparin, S. Jati, C. C. J. Borsalli y E. Marengoni. 2009. Interannual variability of phytoplankton in the main rivers of the Upper Paraná River floodplain, Brazil: influence of upstream reservoirs. Brazilian Journal of Biology 69:501-516. [ Links ]
Shannon, C. E. y W. Weaver. 1963. The mathematical theory of comunication. University of Illinois Press, Urbana. 177 p. [ Links ]
Souza-Fhilo, E. E. 2009. Evaluation of the Upper Paraná River discharge controlled by reservoirs. Brazilian Journal of Biology 69:707-716. [ Links ]
Thomaz, S. M., L. M. Bini y R. L. Bozelli. 2007. Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia 579:1-13. [ Links ]
Thornton, K. W., B. L. Kimmel y F. E. Payne. 1990. Reservoir limnology: ecological perspectives. John Wiley & Sons,Nueva York. 246 p. [ Links ]
Tracanna, B., C. Seeligmann y V. Mirande. 1996. Estudio comparativo de la comunidad fitoplanctónica de dos embalses del Noroeste Argentino. Revista de la Asociación de Ciencias Naturales del Litoral 27:13-22. [ Links ]
Train, S., S. Jati, L. C. Rodrigues y B. M. Pivato. 2005. Distribução espacial e temporal do fitoplâncton en três reservatórios da bacia do Rio Paraná. In Biocenoses em reservatórios: padrões espaciais e temporais, L. Rodrigues, S. M. Thomaz, A. A. Agostinho y L. C. Gomes (eds.). RiMa, São Carlos. p. 73-85. [ Links ]
Tundisi, J. G. 1990. Perspectives for ecological modelling of tropical and subtropical reservoirs in South America. Ecological Modelling 52:7-20. [ Links ]
Tundisi, J. G. 1999. Reservatórios como sistemas Complexos: teoría, aplicaçoes e perspectivas para usos múltiplos. In Ecología de Reservatórios: Estrutura, Função e Aspectos Sociais, R. Henry (ed.). Fundação de Aparo à Pesquisa do Estado de São Paulo-Fundação do Instituto de Biociências (FAPESP-FUNDIBIO). Botucatu, Brasil. p. 21-38. [ Links ]
Tundisi, J. G., T. Matsumura-Tundisi y O. Rocha. 1999. Theorical Basis for Reservoir Management. In Theorical Reservoir Ecology and its Application, J. G. Tundisi y M. Straskraba (eds.). Backhuys Publishers, São Carlos. p. 505-528. [ Links ]
Tundisi, J. G. y T. Matsumura-Tundisi. 2003. Integration of research and management in optimizing multiple uses of reservoirs: the experience in South America and Brazilian case studies. Hydrobiologia 500:231-242. [ Links ]
Utermöhl, H. 1958. Zur Vervollkommung der quantitativen Phytoplankton Methodik. Mitt. Vereinigung für theoretische und angewandte Limnologie 38 p. [ Links ]
Zalocar de Domitrovic, Y. y M. Forastier. 2005. Biodiversidad de Cyanophyceae (Cianobacteria) y especies toxigénicas del litoral fluvial argentino. Instituto Superior de Correlación Geológica (INSUGEO). Miscelánea 14:213-228. [ Links ]
Zalocar de Domitrovic, Y., A. S. G. Poi de Neiff y S. L. Casco. 2007. Abundance and diversity of phytoplankton in the Paraná River (Argentina) 220 km downstream of the Yacyretá reservoir. Brazilian Journal of Biology 67:53-63. [ Links ]

