










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgrociencia
versión On-line ISSN 2521-9766versión impresa ISSN 1405-3195
Agrociencia vol.42 no.6 Texcoco ago./sep. 2008
Protección vegetal
Detection of Citrus sudden death–associated virus and Haplotypes of Citrus tristeza virus in Toxoptera citricida and Aphis spiraecola and implication on citrus sudden death in Brazil
Detección del Citrus sudden death–associated virus y Haplotipos del Citrus tristeza virus en Toxoptera citricida y Aphis spiraecola e implicaciones en la muerte súbita de los cítricos en Brasil
Emiliano Loeza–Kuk1 , Daniel L. Ochoa–Martínez1 , Gustavo Mora–Aguilera1,* , Patricia Rivas–Valencia1 , M. Alejandra Gutiérrez–Espinosa2 , Waldir Cintra de Jesús Junior3 , Ángel Villegas–Monter2 , Nelson Arno–Wulff4 and Eugenio Perez–Molphe–Balch5
1 Fitopatología y
2 Fruticultura. Campus Montecillo. Colegio de Postgraduados. 56230. Montecillo, Estado de México. (loeza01@colpos.mx) (ldaniel@colpos.mx) *Author for correspondence: (morag@colpos.mx).
3 Universidade Federal do Espírito Santo, CEP 29500–000, Alegre, ES, Brasil.
4 FUNDECITRUS A. C. CEP 14807–040, Araracuara, SP, Brasil.
5 Universidad Autónoma de Aguascalientes. Avenida Universidad 940. 20100 Aguascalientes, Aguascalientes. México.
Received: November, 2007.
Aproved: June, 2008.
Abstract
Individuals of Toxoptera citricida (Kirkaldy) and Aphis spiraecola (Patch) collected in 16 commercial citrus orchards of the São Paulo and Minas Gerais States of Brazil were analyzed for virus presence. Orchards of Minas Gerais were selected because citrus trees exhibit symptoms of citrus sudden death (CSD) and in São Paulo due the presence of severe strains of Citrus tristeza virus (CTV), in order to detect genomic sequences of any of these strains that could be associated to CSD. Samples made up of 20 aphids from the localities of Comendador Gomes, Frutal, Prata and Uberlândia, in the State of Minas Gerais, a CSD associated virus (CSDaV) was often found through nested RT–PCR; in samples from São Paulo, none showed positive for the same primers. With the aim to find relationships between the presence of a specific strain of CTV and the expression of CSD, specific probes were used for sequences associated to severity of isolates. The results were negative and the most promising option was to detect CTV sequence variants acquired by aphids through SSCP, to determine if there was a preference for any of the variants. No CTV sequence variants exclusive to trees with CSD symptoms were detected in aphids, but CTV was more prevalent than CSDaV in both aphid species (98 and 90% vs. 42 and 10%). Results confirm the presence of CSDaV in aphids colonizing trees with or without CSD symptoms. However, detection of CSDaV was more frequent under the CSD condition and five times higher in T. citricida.
Key words: Aphids, virus detection, Citrus tristeza virus, Citrus sudden death–associated virus.
Resumen
Se analizaron individuos de Toxoptera citricida (Kirkaldy) y Aphis spiraecola (Patch) recolectados en 16 huertas comerciales de cítricos en los Estados de São Paulo y Minas Gerais, para evaluar la presencia de virus. Las huertas de Minas Gerais se seleccionaron debido a que los árboles de cítricos exhibían síntomas de muerte súbita de los cítricos (MSC) y en São Paulo debido a la presencia de variantes severas del Citrus tristeza virus (CTV), que pudieran permitir detectar secuencias genómicas de cualquiera de estas variantes en asociación con MSC. En grupos muestrales de 20 áfidos, provenientes de las localidades de Comendador Gomes, Frutal, Prata y Uberlândia, del Estado de Minas Gerais, se encontró con frecuencia un CSD associated virus (CSDaV) mediante el uso de RT–PCR y PCR anidado; en muestras de São Paulo, ninguna fue positiva para los mismos iniciadores. Con el objetivo de encontrar relación entre la presencia de una variante específica de CTV y la expresión de MSC, se usaron sondas específicas para secuencias asociadas a la severidad de los aislamientos. Los resultados fueron negativos y la opción más viable fue detectar variantes de secuencias de CTV adquiridas por áfidos a través de SSCP para determinar si existió preferencia por alguna de las variantes. No se detectaron en áfidos variantes de secuencias de CTV exclusivas a árboles con síntomas de MSC, pero el CTV fue más común que el CSDaV en ambas especies de áfidos (98 y 90% vs. 42 y 10%). Los resultados confirman la presencia del CSDaV en áfidos que colonizaron árboles con o sin síntomas de MSC. Sin embargo, la detección del CSDaV fue más frecuente en la condición de MSC y cinco veces mayor en T. citricida.
Palabras clave: Áfidos, detección viral, Citrus tristeza virus, Citrus sudden death–associated virus.
INTRODUCTION
In 1999 a new disease causing the accelerated collapse of the affected plants appeared in some citrus producing areas of Brazil being named citrus sudden death (CSD). Although etiology of the disease is unknown, Roman et al. (2004) reported that it is transmitted by grafting and that fungi, viroids, and endogen bacteria were not the causal agents.
CSD shares several common aspects with the disease caused by Citrus tristeza virus (CTV), such as deterioration at the bud union of the graft, fast decline, and temporal and spatial epidemiological parameters (Bassanezi et al., 2003). For these reasons it was proposed that CSD was caused by a new strain of CTV. This strain would have broken the effect conferred by the use of CTV tolerant rootstocks and cross protection, which has been extensively used in Brazil (Müller and Costa, 1977; Müller et al., 1999; Machado et al., 2004). Spatial and temporal epidemiological analysis of CSD progress indicates that its spreading could be associated with a winged vector, such as an aphid (Bassanezi et al., 2003). This would be in agreement with the hypothesis that CSD is caused by a mutant of CTV. Certain severe CTV isolates are acquired and transmitted at higher rates by its aphid vectors (Ballester–Olmos et al., 1993; Brlansky et al., 2003; Huang et al., 2005). However, no particular CTV isolate has been directly implicated in CSD after extensive molecular analysis using plant tissue (Targon et al., 2003; Román et al., 2004; Rivas–Valencia et al., 2008). A second hypothesis about the etiology of CSD has implicated a marafivirus named Citrus sudden death–associated virus (CSDaV). This virus has been consistently found in samples but in areas free of the disease (Macherroni et al., 2005). Due to the fact that it is a fast spreading disease (Bassanezi et al., 2003; Román et al., 2004), a highly effective vector should be involved. In addition, virus detection on aphid vectors may narrow than the number of haplotypes being actively moved on the field, opening another approach to study a possible association of CTV with CSD.
The objective of this study was to determine if the population of CTV haplotypes detected in Toxoptera citricida and Aphis spiraecola from regions affected with CSD differs from those found in CSD free regions, in order to provide evidence of CTV association with CSD as well as to relate the detection of CSDaV with aphid species and structure of CTV haplotypes.
MATERIALS AND METHODS
Aphid sampling
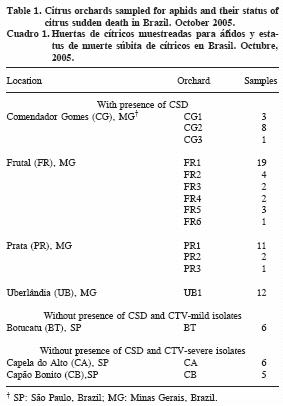
The collections were done in September and October of 2005, in sixteen sites of citrus regions of the States of Minas Gerais and São Paulo, Brazil (Table 1). Groups of 50–100 aphids colonizing citrus shoots were collected, and kept in alcohol 70 %. Two species were collected separately, T. citricida and A. spiraecola based on visual characteristics further confirmed in the laboratory (Stoetzel, 1994). In orchards where insecticides prevented aphid colonization, individuals were taken from peripheral orchards with the same attribute regarding CSD.
Total RNA extraction
For the extraction of RNA, 20 wingless adult aphids were taken, removing the alcohol and macerating them with a pistil in sterile microcentrifuge tubes with 100 µL CTAB 2% (NaCl 1.4M, Tris–HCl 0.1M pH 8.0) buffer, and were kept in cold conditions (Freitas–Astúa et al., 2003). After maceration, 500 µL of CTAB 2% +0.5 % de β–mercaptoethanol buffer prepared with DEPC water was added, mixed in vortex, and incubated at 55 °C for 20 min. Then 400 µL chloroform:isoamyl alcohol 24:1 were added, mixed in vortex, and centrifuged at 12 000 rpm for 10 min at 4 °C. From the water phase 400 µL were taken and transferred to a sterile microcentrifuge tube where another 400 µL of 24:1 chloroform: isoamilic alcohol were added. From the upper phase 400 µL were taken, transferred to a new microcentrifuge tube, added 1/10 volume of ammonium acetate (40 µL), one volume of isopropyl alcohol (400 µL) and precipitated at —20 °C for 15 min. Afterwards, it was centrifuged at 12 000 rpm for 5 min, the upper phase was discarded, the pellet was washed with 1 mL ethanol 70% prepared with DEPC water, and centrifuged at 12 000 rpm for 1 min. After centrifugation, the liquid was removed from the tubes and dried at room temperature and the pellets were suspended in 30 µL DEPC water. The quality and quantity of RNA obtained were corroborated in agarose gel at 1 %.
RT–PCR and nested PCR for virus detection in aphids
For detection of CTV and CSDaV in aphids, one step RT–PCR was done with primers CN119 and CN120 flanking the gene of protein p25 of CTV (Cevik et al., 1996b), as well as the primers CSDaV–F (Alellix Applied Genomics) and CSDaV–R (FUNDECITRUS A. C.) for the detection of CSDaV (Table 2). The reaction mixture consisted of 50 mM KCl, 20 mM Tris–HCl (pH 8.4), 10 mM DTT, 2.5 mM MgCl2, 100 µM dNTPs, 1 ng of each primer, 10 U RNaseOUT (Invitrogen, Carslbad, CA, USA), 25 U MMLV reverse transcriptase (Invitrogen) and 0.75 U Taq DNA polymerase (Invitrogen).
From the reaction mixture, 20 µL were taken and added to 5 µL of total RNA, denatured at 70 °C for 5 min for a total volume of 25 µL. The program for the RT–PCR was: 60 min of reverse transcription at 42 °C, 2 min at 94 °C for RT inactivation, 35 cycles of 30 s at 94 °C, 1 min at 55 °C, 1 min at 72 °C and a final extension of 5 min at 72 °C for CTV. In the case of CSDaV, 35 cycles of 30 sec at 94 °C, 1 min at 55 °C, and 3 min at 72 °C and a final extension of 10 min at 72 °C.
The RT–PCR product of both reactions (p25 of CTV and RdRp of CSDaV) was used to carry out a second amplification using the internal primers for each amplified sequence CPK for CTV (Kong et al., 2000); C54 and C1 for genomic variants of CSDaV, both kindly provided by T. Nagata (Universidade Católica de Brasilia, Brazil).
The reaction mixture consisted of 50 mM KCl, 20 mM Tris–HCl (pH 8.4), 2.5 mM MgCl2, 100 /iM dNTPs, 100 /iM of each primer and 0.6 U Taq pol (Invitrogen). PCR started with a denaturing at 94 °C for 2 min. Followed by 35 amplification cycles of 94 °C for 30 s. (Tm=50 °C for CTV and C54, 55 °C for C1) for 30 s and 72 °C for 30 s, followed by a final extension of 72 °C for 5 min. For reactions, the absolute negative control was sterile water, and as positive control virions purified by density gradients were used from tissue of infected trees symptomatic to CSDaV and CTV.
Detection of genomic strains of CTV and CSDaV acquired by aphids through single–strand conformation polymorphism (SSCP)
The identification of patterns of SSCP was done from the CPK product, as reported by Sambade et al. (2002). Strands were separated in a non–denaturing gel of polyacrylamide at 12% in a constant voltage of 200 V for 2.5 h. The gels were visualized using silver nitrate staining (Beidler et al., 1982).
RESULTS AND DISCUSSION
Eight six aphid samples were collected from 16 orchards. The most common citrus colonizing species were T. citricida (65%) and A. spiraecola (34%). In several orchards insecticides were being used for effective pest control and thus aphids were not found.
Attempts to detect CTV by direct RT–PCR in aphids failed probably due to the low concentration of the viral sequences. The option was to perform nested RT–PCR using CN119 and CN120 as external primers (Cevik et al., 1996b) and CPK as internal primers (Kong et al., 2000). With this adaptation, 55/56 samples of T. citricida and 27/30 of A. spiraecola were positive for CTV presence independently of virus aggressiveness (Table 3). The use of cross protection and the endemic condition of CTV in the region can explain the high incidence of CTV (Müller et al., 1999; Machado et al., 2004).
Detection of CSDaV in plants and aphids has been performed using primers for the genomic region of the viral replicase (Macherroni et al., 2005). In our study, CSDaV was also detected in aphid and plant samples using primers for the genomic region which encodes for the RNA dependent–RNA polymerase (RdRp), which presumably discriminate between C54 and C1 genomic variants (Macherroni et al., 2005).
Strain C54 was found in 23/56 of T. citricida samples and in 3/30 of A. spiraecola aphid samples from orchards of Minas Gerais (Table 3). This amplification product was sequenced and compared with other sequences in the Genbank, which coincided with sequences of CSDaV and CSDaV variant P15, codes AY884005.1 and DQ185573.1. These results confirmed that band obtained was indeed the targeted sequence. Samples from orchards free of CSD located in São Paulo were negatives.
Discrimination of CTV strains present in aphids
Discrimination of CTV strains present in aphids was originally proposed with probes marked in the 3' end with digoxigenine. However, even when using probe 0, which detects all CTV variants (Cevik et al., 1996a), reaction was detected only in test samples with RNA extracted from plants. These results show that sensibility of this technique is low and is not able to detect low CTV sequence concentrations in aphids. Therefore, the variation of haplotypes of CTV present in the collected aphids was analyzed by SSCP; each band was considered to correspond to a strand of the duplex DNA of the gene p25 fragment.
SSCP analysis of sequence variants of CTV in T. citricida and A. spiraecola did not show a specific profile which would indicate a lower or greater capability of CTV genomic variant acquisition, but seems to place both vector species at the same capability level, at least in the acquisition stage (Table 4 and 5), although there is evidence of the greater efficiency of T. citricida as a vector (Huang et al., 2005). However, T. citricida had higher capability to acquire CSDaV than A. spiraecola when CTV was present. This result may indicate a possible role of CTV as helper virus as reported by Maccherroni et al. 2005, but there is not enough data to prove it. If such helper condition does exist it may also depend on the aphid species, being T. citricida more suitable than A. spiraecola. Although other aphid species, such as A. gossypii, may not be ruled out (Maccherroni et al., 2005), the effective citrus colonizing condition may be determinant in a possible CTV–CSDaV–vector interaction.
CSDaV detection rates were 48% and 88% less effective in T. citricida and A. spiraecola, than that of CTV (Table 3), evidencing that the later virus is more abundant and maybe efficiently acquired.
The transmission of CSDaV by A. spiraecola has only been suggested (Macherroni et al., 2005); besides the importance of this aphid species as a vector of CSDaV, if any, could be relatively low at field level. Thus focus should be placed in T. citricida regarding its role on CSD under the hypothesis of the CSDaV. This hypothesis was appealing due to the fact that CSDaV was in general associated with aphids collected from orchards with CSD symptoms, even from those sub–grafted with Citrus reshni (Hort.) that did not cause symptoms remission as expected (PR1), such as samples 11 and 12 (Figure 1). None aphid sample from São Paulo, free of CSD, showed positive to CSDaV.
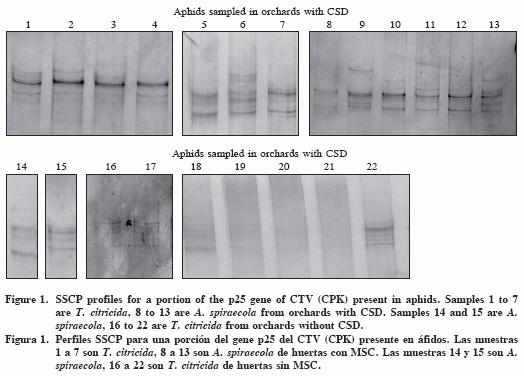
Similarly, samples 1, 3, and 4 of T. citricida from an orchard with all trees symptomatic to CSD (FR1) were positive for CSDaV. Sample 7 positive for CSDaV and collected from a one year established orchard (CG2), without CSD symptoms but surrounded by plots with CSD symptoms, could provide evidence of the role of CSDaV in the etiology of CSD disease and its transmission to new plantations. In the Uberlândia orchard (UB1) FUNDECITRUS's where technicians classified the trees in function of CSD severity according to Román et al. (2004), there was no specific association of SSCP profiles with CSD severity other than higher complexity.
A sample collected from a tree rated with severity–1 (loss of swelling and shine in leaves) had two CTV sequence variants, while a sample with greater severity (2) had three CTV sequence variants. Other samples with the same severity showed two sequence variants and another two asymptomatic samples showed a single sequence variant.
A drawback in associating CSDaV with CSD was that no correlation was found among CSDaV presence in all aphid samples collected from symptomatic plants. Yet the presence of this virus in possible vectors provides more elements to understand the behavior of the disease and seem to reduce the feasibility of the hypothesis of a mutant CTV isolate as the cause of CSD. Besides, CSDaV haplotypes homogenecity, even among samples from different orchards, may suggest the recent development or establishment of this virus, since only a small variability was found.
If in fact the putative agent is aphid or winged vector transmitted as the epidemiological data suggest (Bassanezi et al., 2003; Román et al., 2004), then the vector system should be exploited to support the CSD etiology. But in order to confirm the mutant CTV isolate inducing CSD hypothesis, more studies are needed where CTV profiles are analyzed simultaneously in aphids and the shoot where aphids are collected due the uneven distribution of this virus. In our study, only two pairs of primers and seven probes for the p25 genomic region of CTV were used. Probably other genomic regions may provide more insight results and additional studies with molecular and biological approaches should be conducted, since CTV haplotype complexity was lower on the vector than that on the plant (Rivas et al., 2008) confirming our assumption for the study.
CONCLUSIONS
SSCP analysis of Citrus tristeza virus sequence variants present in T. citricida and A. spiraecola, sampled from citrus sudden death symptomatic or asymptomatic trees but endemic to CTV, did not show a relation between patterns. Therefore a mutant CTV isolate inducing CSD hypothesis was not confirmed.
In contrast, Citrus sudden death–associated virus was in general also detected in both aphid species sampled on trees from citrus regions with presence of citrus sudden death. Thus the CSDaV hypothesis could be more feasible. However, detection was 48 (T. citricida) and 88% (A. spiraecola) lower than that of CTV. Therefore, under the CSDaV scenario, the high CSD epidemic intensity could be explained for a CTV–CSDaV–vector interaction system.
ACKNOWLEDGMENTS
The authors wish to thank to CONACYT for finantial support, FUNDECITRUS for technical and logistical assistance and to Juliana Freitas–Astúa for the RNA isolation protocol.
LITERATURE CITED
Ballester–Olmos J. F., J. A. Pina, E. A. Carbonell, P. Moreno, A. Hermoso de Mendoza, M. Cambra, and L. Navarro. 1993. Biological diversity of citrus tristeza virus (CTV) isolates in Spain. Plant Pathol. 42: 219–229. [ Links ]
Bassanezi R. B., A. Bergamin–Filho, L. Amorim, N. Gimenez– Fernándes, T. R. Gottwald, and J. M. Bové. 2003. Spatial and temporal analyses of citrus sudden death as a tool to generate hypotheses concerning its etiology. Phytopathology 93: 502–512. [ Links ]
Beidler J. L., P. R. Hilliard, and R. L. Rill. 1982. Ultrasensitive staining of nucleic acid with silver. Analytical Biochem. 126: 374–380. [ Links ]
Brlansky R. H., V. D. Damsteeggt, D. S. Howd, and A. Roy. 2003. Molecular analyses of citrus tristeza virus subisolates separated by aphid transmission. Plant Dis. 87: 397–401. [ Links ]
Cevik B., S. S. Pappu, R. F. Lee, and C. L. Niblett. 1996a. Detection and differentiation of strains of citrus tristeza closterovirus using point mutation and minor sequence differences in their coat protein genes. Phytopathology 86: S101. [ Links ]
Cevik B., S. S. Pappu, H. R. Pappu, D. Benscher, M. Irey, R. F.Lee, and C. L. Niblett. 1996b. Application of bi–directional PCR to citrus tristeza virus: Detection and strain differentiation. In: Proceeding of the 13th Conference of IOCV. Riverside, CA. pp: 17–24. [ Links ]
Freitas–Astúa J., E. C. Locali, R. Antoniolo, V. Rodrigues, E. W. Kitajima, and M. A. Machado. 2003. Detection of citrus leprosis vírus in citrus stems, fruits and the mite vector. Virus Rev. Res. 8(1): 196. [ Links ]
Huang Z., P. A. Rundell, X. Guan, and C. A. Powell. 2005. Evaluation of the transmission of different field sources of citrus tristeza virus and the separation of different genotypes by single brown citrus aphids. Hortscience 40: 687–690. [ Links ]
Kong P., L. Rubio, M. Polek, and B. W. Falk. 2000. Population structure and genetic diversity within California citrus tristeza virus (CTV) isolates. Virus Genes 21: 139–145. [ Links ]
Machado M. A., M. L. P. N. Targón, H. D. Coletta–Filho, e G. W. Müller. 2004. Morte súbita dos citrus. Laranja 25: 69–79. [ Links ]
Macherroni W., M. C. Alegria, C. C. Greggio, J. P. Piazza, R. F. Kamla, P. R. A. Zacharias, M. Bar–Joseph, E. W., Kitajima, L. C. Assumpsao, G. Camarote, J. Cardozo, E. C. Casagrande, F. Ferrari, S. F. Franco, P. F. Giacheto, A. Girasol, H. Jr. Jordao, V. H. A. Silva, L. C. A. Souza, C. I. Aguilar–Vildoso, A. S. Zanca, P. Arruda, J. P. Kitajima, F. C. Reinach, J. A. Ferro, and A. C. R. da Silva. 2005. Identification and genomic characterization of a new virus (Tymoviridae Family) associated with citrus sudden death disease. J. Virol. 79: 3028–3037. [ Links ]
Müller G. W., and A. S. Costa. 1977. Tristeza control in Brazil by preimmunization with mild strains. Proc. Int. Soc. Citriculturist 3: 868–872. [ Links ]
Müller G. W., M. L. P. N. Targón, e M. A. Machado. 1999. Trinta anos do uso do clone pré–imunizado 'Pera IAC' na citricultura paulista. Laranja 20: 385–398. [ Links ]
Rivas–Valencia P., E. Loeza–Kuk, G. Mora–Aguilera, D. Ochoa–Martínez, M. A. Gutiérrez–Espinosa, W. C. Jesús J., C. Malvas, y N. A. Wulff. 2008. Estructura poblacional de aislamientos del Citrus tristeza virus y su asociación con la muerte súbita de los cítricos en Brasil. Agrociencia 42: 85–93. [ Links ]
Román M. P., M. Cambra, J. Juárez, P. Moreno, N. Duran–Vila, F. A. O. Tanaka, E. Alves, E. W. Kitajima, P. T. Yamamoto, R. B. Bassanezi, D. C. Teixeira, W. C. Jesus J., A. J. Ayres, N. Gimenez–Fernandes, F. Rabestein, L. F. Girotto, and J. M. Bove. 2004. Sudden death of citrus in Brazil: a graft–transmissible bud union disease. Plant Disease 88: 453–467. [ Links ]
Sambade A. L. Rubio, S. M. Garnsey, N. Costa, G. W. Müller, M. Peyrou, J. Guerri, and P. Moreno. 2002. Comparison of viral RNA populations of pathogenically distinct isolates of citrus tristeza virus: Application to monitoring cross–protection. Plant Pathol. 51: 257–263. [ Links ]
Stoetzel M. B. 1994. Aphids (homoptera:aphididae) of potential importance on citrus in the United States with illustrated keys to species. Proc. Entomol. Soc. Washington 96: 74–90. [ Links ]
Targon M. P. N., G. Astúa–Monge, L. Kishi, J. Freitas–Astúa, A. A. Souza, F. A. Santos, G. W. Muller, e M. A. Machado. 2003. Avaliaçao de haplotipos do CTV em plantas com sintomas de morte subita dos citros por SSCP e seqüenciamento dos genes da p20 e p23. Summa Phytopatologica 29: 71–72. [ Links ]

