










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgrociencia
versión On-line ISSN 2521-9766versión impresa ISSN 1405-3195
Agrociencia vol.42 no.3 Texcoco abr./may. 2008
Ciencia animal
Respuesta productiva y perfil metabólico y su efecto en días abiertos durante el posparto temprano en vacas holstein
Productive response and metabolic profiles and their effect on open days during early lactation in holstein cows
M. Teresa Jaramillo–Jaimes1, Jorge I. Olivera–López2, M. Teresa Sánchez–Torres Esqueda1, Germán D. Mendoza–Martínez, Miguel Molina–Hernández3 y Carlos M. García–Bojalil1
1 Ganadería. Campus Montecillo. Colegio de Postgraduados. Montecillo, Estado de México. (jajt01@prodigy.net.mx).
2 Departamento de Biología de la Reproducción. CBS. UAMI. San Rafael Atlixco 186. Colonia Vicentina. 04390. México D.F. (jiol@xanum.uam.mx).
3 Instituto de Investigaciones Psicológicas. Universidad Veracruzana. 91000. Xalapa, Veracruz. POB 361. (mimoli@todito.com).
Recibido: Julio, 2007.
Aprobado: Febrero, 2008.
Resumen
La relación entre variables productivas y los niveles séricos de metabolitos que puedan afectar el número de días abiertos (DA) durante el posparto, se estudió en 35 vacas Holstein multíparas. Las vacas fueron clasificadas en cuatro grupos de acuerdo con los DA (I: <80 DA; II: 80 a 110 DA; III: 111 a 150 DA y IV: > 150 DA). Los cambios de peso, condición corporal (CC) y producción de leche se evaluaron semanalmente, además de los niveles séricos de proteína (PT), nitrógeno uréico (N–UREA), lipoproteínas de baja densidad (LBD) y colesterol (CT). El diseño experimental fue completamente al azar y los datos se analizaron como mediciones repetidas; además, se hizo una prueba de homogeneidad de regresión y un análisis de correlación. La producción de leche y el peso corporal no fueron estadísticamente diferentes. La CC fue diferente (p<0.01), para un contraste entre el grupo I con los grupos II, III y IV, y diferente (p<0.05) para un segundo contraste entre los grupos II y III. En los metabolitos sólo hubo diferencias (p<0.01) para proteínas entre los grupos con mayor número de DA. Hubo una alta correlación entre el CT y la producción de leche (R=0.53), la semana posparto (R = 0.72) y las LBD (R=0.61). Se concluye que las variables productivas y los metabolitos en este estudio no influencian la función reproductiva reflejada en los DA de vacas Holstein en el posparto temprano.
Palabras clave: Días abiertos, metabolitos séricos, posparto temprano, vacas Holstein, variables productivas.
Abstract
The relationship between productive variables and the serum levels of metabolites that can affect the number of open days (OD) during early lactation, was studied in 35 multiparous Holstein cows. The cows were classified in four groups according to the OD (I: 80 OD; II: 80 to 110 OD; III: 111 to 150 OD and IV: > 150 OD). The changes in weight, body condition (BC) and milk production were evaluated weekly, along with the serum levels of protein (PT), ureic nitrogen (N–UREA), low density lipoproteins (LDL) and cholesterol (CT). The experimental design was completely randomized and the data were analyzed as repeated measurements; in addition, a test of regression homogeneity was made along with an analysis of correlation. Milk production and weight gain were not statistically different. The BC was different (p<0.01), for a contrast between group I against groups II, III and IV, and different (p<0.05) for a second contrast between groups II and III. In the metabolites there were only differences (p<0.01) for proteins among the groups with the highest number of OD. There was a high correlation between CT and milk production (R = 0.53), the week of post–partum (R = 0.72) and the LDL (R=0.61). It is concluded that the productive variables and metabolites in this study do not influence the reproductive function reflected in the OD of Holstein cows in early lactation.
Key words: Open days, serum metabolites,early lactation, Holstein cows, productive variables.
INTRODUCCIÓN
La capacidad genética para la producción de leche en vacas ha aumentado significativamente debido a la selección, pero las tasas de concepción para vacas lactantes se han reducido entre 40 y 50% (Butler, 2000). La disminución en la fertilidad se puede explicar por su relación inversa con la producción de leche (Ahmad et al., 1996; Butler, 2000), por problemas de detección de estros en situaciones de confinamiento y por la condición corporal (CC) al parto. Esta última está relacionada con los cambios metabólicos requeridos para aumentar la producción de leche, influenciando el número de días abiertos (Staples et al., 1992).
La reanudación temprana de la actividad ovárica en vacas posparto es necesaria para lograr la gestación al primer servicio en intervalos de 55 a 85 d. Después de la regresión del cuerpo lúteo de gestación, existe un periodo de inactividad antes de la primera ovulación. La duración de este periodo cambiaría debido al nivel de nutrición, la CC, y la lactancia, entre otros factores (Savio et al., 1990).
El propósito del presente estudio fue identificar algunos indicadores del reinicio de la actividad ovárica posparto reflejados en los DA en vacas Holstein, en función de los niveles de metabolitos séricos (colesterol, lipoproteínas de baja densidad, proteína y nitrógeno uréico), y variables productivas (CC, peso corporal y producción de leche semanal).
MATERIALES Y MÉTODOS
Animales experimentales y alimentación
En este estudio se usaron 35 vacas Holstein de registro multíparas (segundo a séptimo parto) de alta producción de un hato comercial altamente tecnificado, en Zumpango, Estado de México. Las vacas fueron clasificadas en cuatro grupos de acuerdo con el número de días abiertos (DA): Grupo I (GPO I; N = 8), con menos de 80 DA; Grupo II (GPO II, N = 8), entre 80 y 110 DA; Grupo III (GPO III; N = 5), entre 111 y 150 DA; Grupo IV (GPO IV; N=14), con más de 150 DA. Las vacas recibieron una dieta integral que en las dos primeras semanas posparto contenía 16.09% PC y 1.54 Mcal EN kg –1 MS y después se cambió a 19% PC y 2.4 Mcal EN.
Muestras de sangre
Para determinar metabolitos se obtuvo una muestra semanal de 10 mL de sangre por punción de la vena caudal hasta la 7ª semana posparto (SPP). La proteína (PT) se midió por el método de Biuret (Kit 6370, Ames, Technicon, Tarrytown, N.Y.); el nitrógeno uréico (N–UREA) en suero por diacetilmonoxamina (Kit 535, Sigma Diagnostics, St. Louis, MO, USA); las lipoproteínas de baja densidad (LBD) por el método PVS (polivinilsulfonato; Kit 726290, adicionalmente MPR 1/2/3 colesterol CHOD–PAP; Kit 290 319, Boehringer Mannheim, France); y el colesterol (CT) por el método CHOD–PAP, prueba enzimática colorimétrica (Kit 290 319, Boehringer Mannheim, France).
Condición corporal, peso corporal (PC) y producción de leche (PL)
Al inicio del estudio se pesaron todas las vacas y se clasificaron de acuerdo con su CC, siguiendo la escala de Wildman et al. (1982) y Ferguson et al. (1994). Esta evaluación se hizo semanalmente para ambas variables hasta la 7ª SPP. La leche se pesó una vez por semana (mañana y tarde) usando el pesador de leche (garrafón Alfa Laval).
Análisis estadístico
El diseño experimental fue completamente al azar y se hizo un análisis de varianza de mínimos cuadrados usando el procedimiento GLM (SAS, 1990). El modelo lineal para el análisis de las variables productivas y metabolitos séricos incluyó los efectos de grupo, animales dentro del grupo, SPP, y la interacción del grupo con SPP. Las comparaciones entre grupos se hicieron con contrastes ortogonales.
Se realizó una prueba de homogeneidad de regresión para comparar cambios en las variables a través del tiempo, por simples curvas de regresión polinomial, para probar la ganancia en ajuste de las regresiones individuales y por grupo (Wilcox et al., 1990).
La relación entre las concentraciones de los metabolitos séricos (PT, N–UREA, LBD y CT) y las variables productivas (PC, CC, PL y SPP) con DA, se determinó con el procedimiento CORR; además, se calculó la regresión de PC, CC y PL sobre DA, utilizando el procedimiento REG (SAS, 1990).
RESULTADOS Y DISCUSIÓN
Cambios de peso, condición corporal y producción de leche
El PC de las vacas (Cuadro 1) fue diferente (p<0.03) en el contraste 1. La CC en el grupo I presentó la menor (p<0.01) calificación (3.03) mientras que el contraste 2 fue diferente (p<0.05) en los grupos II y III. Para PL no hubo diferencias entre grupos.
La respuesta del PC durante el tiempo posparto fue de primer orden para los cuatro grupos experimentales (Figura 1). El grupo I fue diferente respecto a los otros grupos.
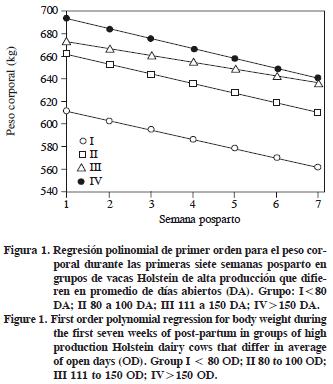
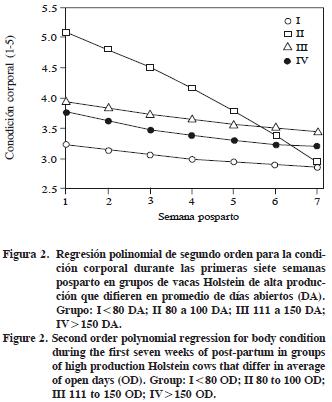
La CC tuvo una respuesta cuadrática y las vacas del grupo II perdieron 2 unidades durante las siete SPP (Figura 2). En vacas con un suplemento (3% de grasa), la máxima movilización de tejido corporal ocurrió entre las 2 a 5 SPP y perdieron 0.45 unidades de CC (Madhav et al., 1998). Según Garnsworthy y Huggett (1992), el incremento en el consumo de una dieta con un suplemento con grasa tiende a disminuir la pérdida de CC en vacas obesas al parto, pero en vacas flacas al parto aumentó la producción de leche. La CC no está relacionada con el número de DA ni influye la PL o función reproductiva, pero los cambios en la CC se relacionan con el pico de producción para la curva de lactancia y con el intervalo del parto a la primera inseminación (Pedron et al., 1993).
En los grupos experimentales hubo una respuesta cuadrática para PL (Figura 3); el grupo I presentó primero su pico con mayor producción, pero disminuyó antes que los otros grupos. Vacas en el posparto temprano alimentadas con un suplemento de grasa en la dieta, no mostraron cambios significativos en consumo de alimento y producción de leche (Komaragiri et al., 1998). Según Pedrón et al. (1993), en vacas que movilizaron más grasa corporal (mayor pérdida en CC) se usaron estas reservas para la producción de leche, sin efectos negativos en la función reproductiva o en incidencias de retenciones placentarias.

Respuestas metabólicas
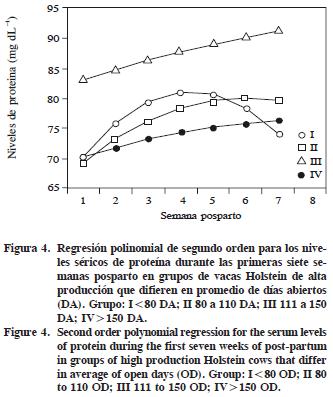
El grupo III presentó la mayor concentración de PT (87.48 mg dL –1) respecto a los otros grupos, y además, hubo diferencias entre los grupos III y IV (p<0.001; Cuadro 2). En la Figura 4 se muestra la respuesta de segundo orden para los niveles séricos de PT de cada grupo durante las 7 SPP. Niveles similares a los de los grupos I, II y IV se encontraron en vacas Gyr durante 12 SPP(Patil y Deshpande, 1979): 7.3 g dL –1 en vacas que manifestaron estro entre 60 y 65 d posparto y 7.1 g dL –1 en aquellas sin estro manifiesto.
El N–uréico no fue diferente (p>0.19) entre los grupos y sus medias (9.64, 9.67, 9.52 y 11.96 mg dL–1) fueron menores a los valores normales (12–18 mg dL–1; Butler 1998, 2000; Hammon et al., 2004). Los niveles de N–UREA durante el posparto en el grupo IV (11.96 mg dL–1) fueron mayores que en los grupos I, II y III (Figura 5). Los niveles de N–uréico se utilizan como indicadores de alteraciones reproductivas (Hammon et al., 2004).

La concentración de LBD no fue diferente (p > 0.21) entre grupos (Cuadro 2). El promedio (29 mg dL–1) de este estudio difiere del valor (8 mg dL–1) reportado por Grummer y Carroll (1988). Los niveles de LBD durante las 7 SPP (Figura 6) para el grupo II muestran una respuesta atípica en relación a los otros grupos. Además, los grupos I, III y IV tuvieron una tendencia similar a partir de la semana tres y no hubo diferencia (p>0.05) durante el posparto. Esta respuesta puede deberse a que las células lúteas bovinas dependen de lipoproteínas que proveen el sustrato para la síntesis de P4. Si las vacas en el grupo IV tienen la mayor concentración de lipoproteínas y el mayor número de días abiertos, entonces las LBD posiblemente no se metabolizan para el proceso de la esteroidogénesis.

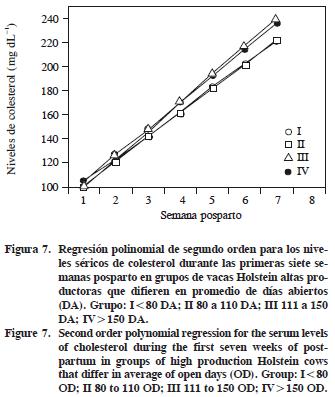
Los niveles de CT no fueron diferentes (p>0.47) entre grupos (Cuadro 2) y en la Figura 7 se muestra su evolución. Williams (1989) reporta un aumento lineal en colesterol desde el parto a las 9 SPP, alcanzando su máximo (215 mg dL–1) entre las 7 y 9 SPP. La concentración plasmática de CT para un grupo testigo fue 230 mg dL–1 y para un grupo tratado con suplemento de grasa inerte fue 250 mg dL–1 para las primeras 7 SPP, con diferencia significativa (p<0.05) en la 5ª SPP (Spicer et al., 1993). Tal resultado es similar al del presente estudio a partir de la 3ª SPP, sólo que los grupos III y IV presentaron concentraciones mayores de CT pero tuvieron más DA.
Correlación entre las variables productivas y metabólicas con DA
Las correlaciones entre los metabolitos sanguíneos y las variables productivas se muestran en el Cuadro 3. El PC y CC tuvieron una alta relación (R=0.78), lo que indica que los cambios de peso se reflejan en la CC. Estas relaciones coinciden con los resultados reportados por Pedron et al. (1993), Komaragiri y Erdman (1997) y Butler (2000), quienes observaron que vacas al parto con una alta CC tienen reservas de grasa suficientes para la demanda en producción láctea, pero reducen el consumo de alimento y hay un balance energético negativo. Esto reduce la actividad ovárica, ya que la glándula mamaria tiene prioridad sobre las funciones ováricas. El periodo posparto (SPP) se asoció negativamente con PC y CC (R=–0.21 y R=–0.33), y positivamente con la PL (R=0.38), PT (R=0.23), LBD (R=0.36) y CT (R=0.72).
El CT tuvo una alta correlación con PL (R=0.53) y LBD (R=0.61). Assmann y Carmena (1990) indican que entre 60 y 70% del total del colesterol circulante es transportado por las LBD. Además, el principal precursor para la síntesis de P4 es el colesterol (Staples et al., 1998). La correlación entre PT y CT fue baja (R=0.25), posiblemente debido a la fracción de proteína presente en la transportación del colesterol en sangre.
CONCLUSIONES
Las variables peso corporal y condición corporal resultaron diferentes para los grupos con más de 110 DA, pero la producción de leche no fue diferente.
Para los metabolitos séricos sólo la PT fue diferente entre los grupos III y IV. Sin embargo, hubo una alta correlación entre el colesterol y la producción de leche, el tiempo posparto y las lipoproteínas de baja densidad; y una mediana relación con la condición corporal, proteínas y nitrógeno uréico. Esto sugiere que hay factores desconocidos que influencian el problema del reinicio de la actividad ovárica postparto.
LITERATURA CITADA
Ahmad, N., S. W. Beam, W. R. Butler, D. R. Deaver, R. T. Duby, D. R. Elder, J. E. Fortune, L. C. Griel, Jr., L. S. Jones, R. A. Milvae, J. L. Pate, I. Revah, D. T. Schreiber, Jr., D. H. Townson, P. C. W. Tsang, and E. K. Inskeep. 1996. Relationship of fertility to patterns of ovarian follicular development and associated hormonal profiles in dairy cows and heifers. J. Anim. Sci. 74: 1943–1952. [ Links ]
Assmann, G. y R. Carmena. 1990. Trastornos del Metabolismo de Lípidos y Cardiopatía Coronaria. Ed. Médica MMV GMBH, Manchen, Munich, Alemania. pp: 42–45. [ Links ]
Butler, W. R. 1998. Review: Effect of protein nutrition on ovarian and uterine physiology in dairy cattle. J. Dairy Sci. 81: 2533–2539. [ Links ]
Butler, W. R. 2000. Nutritional interactions with reproductive performance in dairy cattle. Anim. Reprod. Sci. 60–61: 449–457. [ Links ]
Ferguson, J. D., D. T. Galligan, and N. Thomsen. 1994. Principal descriptors of body condition score in Holstein cows. J. Dairy Sci. 77: 2695–2703. [ Links ]
Garnsworthy, P. C., and C. D. Huggett. 1992. The influence of the fat concentration of the diet on response by dairy cows to body condition at calving. Anim. Prod. 54: 7–13. [ Links ]
Grummer, R. R., and D. J. Carroll. 1988. A review of lipoprotein cholesterol metabolism: Importance to ovarian function. J. Anim. Sci. 66: 3160–3173. [ Links ]
Hammon, D. S., G. R. Holyoak, and T. R. Dhiman. 2004. Association between blood plasma urea nitrogen levels and reproductive fluid urea nitrogen and ammonia concentrations in early lactation dairy cows. Anim. Reprod. Sci. 86: 195–204. [ Links ]
Komaragiri, M. V. S., and R. A. Erdman. 1997. Factors affecting body tissue mobilization in early lactation dairy cows. 1. Effect of dietary protein on mobilization of body fat and protein. J. Dairy Sci. 80: 929–937. [ Links ]
Komaragiri, M. V. S., D. P. Casper, and R. A. Erdman. 1998. Factors affecting body tissue mobilization in early lactation dairy cows. 2. Effect of dietary fat on mobilization of body fat and protein. J. Dairy Sci. 81: 169–175. [ Links ]
Patil, J. S., and B. R. Deshpande. 1979. Changes in body weight, blood glucose and serum proteins in relation to the appearance of post–partum oestrus in Gyr cows. J. Reprod. Fert. 57: 525– 527. [ Links ]
Pedron, O., F. Cheli, E. Senatore, D. Baroli, and R. Rizzi. 1993. Effect of body condition score at calving on performance, some blood parameters, and milk fatty acid composition in dairy cows. J. Dairy Sci. 76: 2528–2535. [ Links ]
SAS 1990. Language Guide for Personal Computers. Version 6.03. SAS Inst., Inc., Cary, NC. 558 p. [ Links ]
Savio, J. D., M. P. Boland, N. Hynes, and J. F. Roche. 1990. Resumption of follicular activity in the early post–partum period of dairy cows. J. Reprod. Fert. 88: 569–579. [ Links ]
Sejrsen, K., and A. Neimann–Sorensen. 1982. Nutritional physiology and feeding of the cow around parturition. Curr. Top. Vet. Med. Anim. Sci. 20: 325–357. [ Links ]
Spicer, L. J., R. K. Vernon, W. B. Tucker, R. P. Wettemann, J. F. Hogue, and G. D. Adams. 1993. Effects of inert fat on energy balance plasma concentrations of hormones, and reproduction in dairy cows. J. Dairy Sci. 76: 2664–2673. [ Links ]
Staples, C. R., W. W. Thatcher, C. M. García–Bojalil, and M. C. Lucy. 1992. Nutritional influences on reproductive function. In: Large Dairy Herd Management; Nutrition: Gainesville, Fl. pp: 382–392. [ Links ]
Staples, C. R., J. M. Burke, and W. W. Thatcher. 1998. Influence of supplemental fats on reproductive tissues and performance of lactating cows. J. Dairy Sci. 81: 856–871. [ Links ]
Wilcox, C. J., W. W. Thatcher, and F. G. Martin. 1990. Statistical analysis of repeated measurements in physiology experiments. In: Livestock Reproduction in Latin America. International Atomic Energy Agency, Vienna. pp:141–155. [ Links ]
Wildman, E. E., G. M. Jones, P. E. Wagner, R. L. Boman, H. F. Troutt, and T. N. Lesch. 1982. A dairy cow body condition scoring system and its relationship to selected production characteristics. J. Dairy Sci. 65: 495–501. [ Links ]
Williams, G. L. 1989. Modulation of luteal activity in postpartum beef cows through changes in dietary lipid. J. Anim. Sci. 67: 785–793. [ Links ]

