











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Las selvas tropicales son formaciones naturales con diferentes tamaños, parches, formas y etapas de regeneración (Leigh, 1999; Hubbell, 2001; Basáñez et al., 2008). Estos ecosistemas representan la vegetación más exuberante del planeta, donde a menudo se encuentran cientos de especies de plantas por hectárea (Challenger y Soberón, 2008), compuestas por vegetación densa, enredaderas y epifitas. Esta composición, diversidad y distribución de las especies vegetales está relacionada con factores como las variaciones en los gradientes altitudinales, el relieve, la temperatura y la precipitación (Grubb, 1977; Sánchez-Gutiérrez et al., 2021).
En México, las selvas ocupan alrededor del 16.3 % de la superficie total del territorio, lo que equivale aproximadamente a 22 793 822.2 ha (INEGI, 2021). De éstas, 10 532 938.03 ha están ocupadas por selvas medianas y en Tabasco están reportadas 60 119.33 ha (SNMF, 2020), las cuales se caracterizan por la dominancia de árboles por debajo de los 25 m de altura, donde abundan los líquenes, musgos y helechos (Pennington y Sarukhán, 2005). Estas selvas han sido tradicionalmente una fuente de extracción de materias primas como la madera, leña, plantas y animales necesarios para la subsistencia de las comunidades rurales. Además, en estos ecosistemas se llevan a cabo procesos vitales como el ciclo del agua y los nutrientes, la formación y el mantenimiento del suelo, la regulación del clima, la erosión y la conservación de la biodiversidad (Challenger y Dirzo, 2009). En Tabasco, las selvas se distribuyen en la región sierra de los municipios de Teapa, Tacotalpa, Macuspana, Tenosique y Huimanguillo, (Maldonado-Sánchez y Maldonado-Mares, 2010; Vázquez-Negrín et al., 2011). En este último podemos encontrar algunos relictos de selvas mejor conservados de Tabasco (Moguel-Ordóñez et al., 2022), donde se desarrollan diferentes actividades ecoturísticas.
Este entorno natural y conservado propició el establecimiento del complejo ecoturístico “Agua Selva", el cual está conformado por ocho comunidades rurales, entre ellas se localiza el Ejido Villa de Guadalupe, en el municipio de Huimanguillo (Rodríguez Ocaña y Banda Izeta, 2016). En los últimos años en la selva cercana a esta localidad se han reportado ocho nuevos registros de herpetofauna para el estado de Tabasco, entre los cuales se encuentra a Quilticohyla zoque, una especie endémica de México que está asociada a sitios conservados (Canseco-Márquez et al., 2017; Ríos-Rodas et al., 2020). En cuanto a la riqueza florística de este sitio, se han reportado 28 especies de helechos con amplia distribución en el país, pero que no habían sido mencionadas para el estado (Carvajal-Hernández et al., 2018), así como la descripción de una nueva especie de la familia Lentibulariaceae, Pinguicula olmeca (Burelo-Ramos et al., 2018).
La selva mediana perennifolia que alberga esta localidad representa un ecosistema con una alta riqueza biológica, a pesar de la relevancia ecológica del sitio, han sido pocos los estudios de investigación sobre su vegetación realizados hasta el momento. Por lo cual el objetivo de esta investigación fue determinar la estructura y diversidad arbórea de la selva mediana perennifolia en el complejo ecoturístico Agua Selva, Tabasco, México.
Métodos
Área de estudio
El estudio se realizó en un área de selva mediana perennifolia que se localiza en el ejido Villa de Guadalupe (17°21’38.23’’ N y 93°36’30.97’’ O), dentro del Complejo Ecoturístico “Agua Selva” en el municipio de Huimanguillo, Tabasco, México. El sitio se encuentra en la región fisiográfica conocida como Sierra Norte de Chiapas, con elevaciones que van desde los 200 a 1000 msnm (Ríos-Rodas et al., 2020). El clima predominante es de tipo cálido húmedo con lluvias todo el año (Af), con precipitación media anual de 3 638 mm y temperatura media anual de 20.6 °C (Carvajal-Hernández et al., 2018; Ríos-Rodas et al., 2020).
En el sitio podemos encontrar dos tipos de vegetación: Primaria, es una selva alta perennifolia, donde predominan árboles de hasta 45 m de altura tales como caoba (Swietenia macrophylla King), ramón (Brosimum alicastrum Swartz), ceiba (Ceiba pentandra Gaertn), jobo (Spondias mombin L), zopo (Guatteria anomala R. E. Fr.) y palo mulato (Bursera simaruba L). La vegetación secundaria está representada por herbáceas de hasta 5 m de altura, arbustos entre 5 y 10 m y árboles de 10 y 30 m como guarumo (Cecropia obtusifolia Bertol), guácimo (Guazuma ulmifolia LAM), jonote (Heliocarpus donnellsmithii Rose), ciruelillo (Trichilia havanensis Jacq) y naranjillo (Bernardia interrupta Sessé) (Palma-López et al., 2011; Rodríguez Ocaña y Banda Izeta, 2016; INEGI, 2017; Ríos-Rodas et al., 2020).
De acuerdo con la clasificación de vegetación propuesta por Pennington y Sarukhán (2005), el área donde se establecieron las unidades de muestreo corresponde a selva alta perennifolia. Este tipo de ecosistema se caracteriza por su gran biodiversidad y la presencia de especies arbóreas de gran porte. Entre las especies representativas se encuentran Swietenia macrophylla King (caoba), Brosimum alicastrum Swartz (ramón), Ceiba pentandra Gaertn (ceiba), Calophyllum brasiliense (bari) y Guatteria anomala R. E. Fr. (zopo), las cuales alcanzan alturas de hasta 45 metros.
Muestreo de la vegetación
Mediante recorridos de campo se establecieron aleatoriamente cinco unidades de muestreo (UM) de 20 x 30 m (600 m2). Cada UM fue dividida en 6 subunidades de muestreo (SUM) de 10 x 10 m (100 m2) para facilitar el inventario de las especies arbóreas (Vázquez-Negrín et al., 2011; López-Toledo et al., 2012; Sánchez-Gutiérrez et al., 2021). En cada unidad de muestreo (UM) se registraron todos los individuos arbóreos con un diámetro normal (DN) de 1.3 m del suelo >2.5 cm. El diámetro normal (DN) se midió con una cinta métrica, la altura total (Ht) con un altímetro (Pistola marca Hagan, modelo SKU 43890, Alemania) y el diámetro de copa, tanto menor como mayor, siguiendo el protocolo de Sánchez-Gutiérrez et al. (2021). Además, cada UM fue descrita y comparada entre sí, con el objetivo de conocer y detallar las características de composición y estructura de cada unidad, lo que permitió una comprensión más precisa de la variabilidad y características particulares de cada sitio de muestreo.
Identificación taxonómica y evaluación del estado de conservación
La identificación y clasificación taxonómica de las especies se realizó utilizando manuales y guías de campo (Pennington y Sarukhán, 2005; Ochoa-Gaona et al., 2012). Adicionalmente, se emplearon claves botánicas y comparaciones con especímenes de herbario, siguiendo los métodos descritos por Gentry (1993). La verificación de los nombres científicos y sus sinonimias se efectuó mediante la consulta de bases de datos internacionales reconocidas, como Trópicos (Missouri Botanical Garden, 2025) y Plants of the World Online (POWO) (Royal Botanic Gardens, 2023).
En los casos donde no fue posible identificar los individuos en campo, se recolectó material botánico (incluyendo corteza, frutos, semillas, ramas y flores) para su determinación posterior en el laboratorio. Cada especie identificada fue evaluada con respecto a su inclusión en: La Lista Roja de la Unión Internacional para la Conservación de la Naturaleza (UICN), a través de su plataforma oficial (UICN, 2023). La NOM-059-SEMARNAT-2019, para determinar su estatus de conservación en México (DOF, 2019).
Análisis de datos
Diversidad arbórea. La diversidad se calculó mediante el índice de diversidad verdadera de orden 1 (1D), donde todas las especies son consideradas en el valor de diversidad, ponderadas proporcionalmente según su abundancia en la comunidad; y la medida de diversidad de orden 2 (2D), en el cual se toman en cuenta las especies más comunes (Jost, 2006, 2007; Tuomisto, 2010; Moreno et al., 2011), con ayuda del programa EstimateS v9.1.0.
La equidad de la comunidad se analizó utilizando el índice de Pielou (J’) con el programa Past v3.14, el cual mide la proporción de la diversidad observada con relación a la máxima diversidad esperada, con valores que van de 0 a 1, donde 1 nos dice que las especies presentan la misma abundancia dentro de la comunidad.
Curvas de rango-abundancia. Se realizaron curvas de rango-abundancia para representar de manera gráfica la abundancia de las especies. Ésta calcula la proporción de individuos respecto al tamaño de la muestra, ordenando las especies de mayor a menor abundancia, a través de la expresión: pi= ni/N, donde “pi” es la abundancia proporcional de la especie i, “ni” es el número de individuos de la especie i y “N” es el número total de individuos, posteriormente se calculó el logaritmo base 10 para cada valor de pi (Feinsinger, 2004).
Estimador no paramétrico Bootstrap. La cobertura de la muestra se estimó a través de Bootstrap obtenido por Smith y van Belle (1984), el cual se basa en la proporción de cuadrantes que contienen cada especie y, por lo tanto, solo requiere datos de incidencia (presencia/ausencia) mediante intervalos de confianza al 95% obtenidos a través de técnicas de remuestreo (Chazdon et al., 1998; Badii et al., 2007).
Estructura arbórea. Para la jerarquización de la dominancia de cada especie en rodales mezclados se calculó el Índice de Valor de Importancia (IVI) (López-Toledo et al., 2012; Sánchez-Gutiérrez et al., 2017) a través de la siguiente ecuación: IVI= Dr + Abr + Fr; donde: “Dr” se refiere a la densidad relativa; “Abr” = Dominancia relativa (empleando área basal); “Fr” = Frecuencia relativa; y se calcularon con las siguientes ecuaciones: Dr= (Número de individuos de una especie/Número de total de individuos de todas las especies) *100; Abr= (Área basal de una especie/área basal de todas las especies) *100. El área basal (AB) de los árboles se obtendrá con la siguiente ecuación: AB= π/4 *DN 2 ; Fr= (Número de parcelas en que se representa una especie/Número de parcelas donde aparecen todas las especies) *100 (Corral-Rivas et al., 2002; Basáñez et al., 2008).
El Índice de Valor Forestal (IVF), a diferencia del IVI, que sólo involucra variables horizontales, incluye la variable vertical de altura (Corella Justavino et al., 2001; López-Toledo et al., 2012; Sánchez-Gutiérrez et al., 2017) y se determinó mediante la ecuación: IVF= DNr + Hr + Cr, donde: “DNr” = Diámetro normal relativo, “Hr” = Altura total relativa, “Cr” = Cobertura de copa relativa; se obtuvieron con las siguientes ecuaciones: DNr= (Diámetro absoluto de cada especie/Diámetro absoluto de todas las especies) *100; donde: Diámetro absoluto= Diámetro de una especie/área muestreada. Hr= (Altura absoluta de cada especie/Altura de todas las especies) *100; donde: Altura absoluta= Altura de una especie/área muestreada. Cr= (Cobertura absoluta de cada especie/Cobertura absoluta de todas las especies) *100; donde: Cobertura absoluta= Cobertura de una especie/Área muestreada. La cobertura se estimó con la ecuación para el área de una elipse: A= π · r1 · r2; donde: π= 3.1416, r1= Radio uno, r2= Radio dos. Esta ecuación supone que las copas de los árboles tienen una forma elíptica regular y varían significativamente en función de diversos factores, como la especie, la competencia por la luz y las condiciones ambientales.
Resultados
En las cinco unidades de muestreo se registró un total de 400 individuos, se identificaron 59 especies de árboles agrupados en 52 géneros y 29 familias. Las familias con mayor riqueza fueron: Rubiaceae (con siete especies), Malvaceae (seis), Fabaceae (cinco), Moraceae (cinco), Meliaceae (cuatro) y Lauraceae (tres), las familias restantes estuvieron representadas por una o dos especies. La mayoría de los géneros (88%) se registraron con una especie. Sin embargo, del género Guarea se registraron tres especies; en cuanto a Ficus, Nectandra, Pouteria, Pseudolmedia y Psychotria fueron dos para cada uno. Las especies con mayor número de individuos fueron Astrocaryum mexicanum con 88, Brosimum alicastrum (37), Oecopetalum mexicanum (35), Dendropanax arboreus (21), Calophyllum brasiliense (16), Nectandra angustifolia (14), Alibertia edulis (11), Sterculia mexicana (11), Rinorea guatemalensis (11) y Urera caracasana (11), estas especies representan el 63.75% del total de las especies con mayor abundancia en las cinco UM (Tabla 1).
Tabla 1 Listado de especies arbóreas registradas en una selva mediana perennifolia en el complejo ecoturístico Agua Selva, Tabasco, México.Table 1. List of tree species recorded in a medium evergreen forest in the Agua Selva ecotourism complex, Tabasco, Mexico.
| Familia/Especie | Abrev. | UM1 | UM 2 | UM 3 | UM 4 | UM 5 | IUCN | NOM-059 |
|---|---|---|---|---|---|---|---|---|
| Anacardiaceae | ||||||||
| Astronium graveolens Jacq. | agr | 1 | 0 | 0 | 0 | 0 | LC | A |
| Spondias mombin L. | smo | 3 | 3 | 0 | 0 | 0 | LC | - |
| Annonaceae | ||||||||
| Annona cherimola Mill | ach | 0 | 1 | 0 | 1 | 0 | LC | - |
| Guatteria anomala R. E. Fr | gan | 1 | 0 | 0 | 0 | 0 | - | A |
| Apocynaceae | ||||||||
| Tabernaemontana donnell-smithii Rose ex J. D. Sm. | tdo | 0 | 0 | 0 | 2 | 1 | LC | - |
| Araliaceae | ||||||||
| Dendropanax arboreus (L.) Decne. & Planch. | dar | 2 | 1 | 2 | 7 | 9 | LC | - |
| Oreopanax capitatus (Jacq.) Decne. & Planch. | oca | 1 | 0 | 2 | 0 | 0 | LC | - |
| Arecaceae | ||||||||
| Astrocaryum mexicanum Liebm. ex Mart | ame | 20 | 19 | 21 | 18 | 10 | LC | - |
| Thrinax radiata Lodd. ex J.A. & J.H. Schult. | tra | 0 | 0 | 0 | 1 | 0 | LC | A |
| Boraginaceae | ||||||||
| Cordia alliodora (Ruiz y Pav.) Oken | cal | 1 | 1 | 0 | 1 | 1 | LC | - |
| Burseraceae | ||||||||
| Bursera simaruba (L.) Sarg. | bsi | 0 | 0 | 1 | 0 | 0 | LC | - |
| Chrysobalanaceae | ||||||||
| Hirtella racemosa Lam. | hra | 0 | 1 | 0 | 0 | 0 | LC | - |
| Clusiaceae | ||||||||
| Calophyllum brasiliense Cambess | cbr | 0 | 2 | 0 | 8 | 6 | LC | A |
| Clusia guatemalensis Hemsl. | cgu | 0 | 0 | 0 | 1 | 0 | LC | - |
| Combretaceae | ||||||||
| Terminalia amazonia (J. F. Gmel.) Excell | tam | 0 | 2 | 0 | 0 | 0 | LC | - |
| Ebenaceae | ||||||||
| Diospyros digyna Jacq. | ddi | 0 | 0 | 0 | 1 | 0 | LC | - |
| Euphorbiacea | ||||||||
| Acalypha skutchii I. M. Johnst | ask | 0 | 4 | 3 | 0 | 0 | LC | - |
| Fabaceae | ||||||||
| Ebenopsis ebano (Berland.) Barneby & J. W. Grimes | eeb | 0 | 0 | 0 | 6 | 0 | LC | - |
| Erythrina caribaea Krukoff & Barneby | eca | 1 | 0 | 0 | 0 | 0 | VU | - |
| Inga sp. | isp | 1 | 0 | 0 | 1 | 0 | - | - |
| Lonchocarpus guatemalensis Benth. | lgu | 0 | 2 | 1 | 3 | 3 | LC | - |
| Pterocarpus rohrii Vahl | pro | 0 | 1 | 2 | 0 | 0 | LC | - |
| Icacinaceae | ||||||||
| Oecopetalum mexicanum Grenm. & C. H. Thomps | ome | 7 | 7 | 14 | 4 | 3 | LC | - |
| Lauraceae | ||||||||
| Licaria capitata (Cham. & Schltdl.) Kosterm. | lca | 0 | 0 | 0 | 3 | 2 | LC | - |
| Nectandra ambigens (S. F. Blake) C. K. Allen | nam | 0 | 0 | 0 | 2 | 2 | LC | - |
| Nectandra angustifolia (Schrad.) Nees & Mart. | nan | 1 | 3 | 2 | 5 | 3 | - | - |
| Malvaceae | ||||||||
| Hibiscus elatus S. W. | hel | 0 | 0 | 2 | 2 | 1 | - | - |
| Ochroma pyramidale (Cav. ex Lam.) Urb. | opy | 0 | 1 | 1 | 0 | 0 | LC | - |
| Pseudobombax ellipticum (Kunth) Dugand | pel | 0 | 0 | 0 | 5 | 3 | LC | - |
| Quararibea funebris (La llave) Vischer | qfu | 1 | 0 | 1 | 0 | 0 | - | - |
| Sterculia mexicanaR. Br. | sme | 2 | 2 | 4 | 2 | 1 | - | - |
| Theobroma bicolor Humb. & Bonpl. | tbi | 0 | 0 | 0 | 1 | 0 | - | - |
| Melastomataceae | ||||||||
| Miconia sp | msp | 0 | 0 | 0 | 1 | 0 | - | - |
| Meliaceae | ||||||||
| Guarea donnell-smithii C. DC. | gdo | 0 | 1 | 0 | 1 | 0 | - | - |
| Guarea glabra Vahl | ggl | 0 | 1 | 0 | 1 | 1 | VU | - |
| Guarea grandifolia A. DC. | ggr | 1 | 1 | 2 | 3 | 2 | - | - |
| Trichilia japurensis C. DC. | tja | 2 | 1 | 2 | 1 | 2 | - | - |
| Moraceae | ||||||||
| Brosimum alicastrum Sw. | bal | 0 | 16 | 9 | 7 | 5 | LC | - |
| Ficus maxima Mill. | fma | 0 | 0 | 0 | 1 | 1 | LC | - |
| Ficus padifolia Kunth | fpa | 0 | 1 | 2 | 0 | 0 | LC | - |
| Pseudolmedia glabrata (Liebm.) C. C. Berg | pgl | 0 | 0 | 1 | 0 | 0 | LC | - |
| Pseudolmedia oxiphyllaria Donn. Sm. | pox | 1 | 0 | 0 | 0 | 0 | LC | - |
| Myrtaceae | ||||||||
| Eugenia acapulcensis Steud. | eac | 0 | 0 | 0 | 1 | 0 | LC | - |
| Polygonaceae | ||||||||
| Coccoloba barbadensis Jacq. | cba | 0 | 0 | 1 | 1 | 1 | LC | - |
| Primulaceae | ||||||||
| Ardisia fimbrillifera Lundell | afi | 0 | 1 | 0 | 2 | 2 | LC | - |
| Rubiaceae | ||||||||
| Alibertia edulis (Rich.) A. Rich. ex DC. | aed | 2 | 1 | 1 | 4 | 3 | LC | - |
| Blepharidium mexicanum Standl | bme | 0 | 0 | 0 | 1 | 0 | LC | - |
| Faramea occidentalis (L.) A. Rich. | foc | 3 | 2 | 0 | 0 | 1 | LC | - |
| Genipa americana L. | gam | 0 | 0 | 0 | 1 | 0 | LC | - |
| Posoqueria sp. | psp | 1 | 0 | 0 | 0 | 0 | - | - |
| Psychotria chiapanensis Standl. | pch | 2 | 0 | 0 | 0 | 0 | LC | - |
| Psychotria elata (Sw.) Hammel | pse | 0 | 1 | 1 | 0 | 0 | LC | - |
| Sapindaceae | ||||||||
| Cupania dentata Moc. & Sessé ex DC | cde | 2 | 0 | 0 | 1 | 0 | LC | - |
| Sapotaceae | - | - | ||||||
| Pouteria campechiana (Kunth) Baehmi | pca | 1 | 0 | 0 | 0 | 0 | LC | - |
| Pouteria multiflora (A. DC.) Eyma | pmu | 0 | 1 | 1 | 1 | 1 | - | - |
| Siparunaceae | ||||||||
| Siparuna andina (Tul.) A. DC. | san | 0 | 0 | 0 | 1 | 1 | LC | - |
| Tiliaceae | ||||||||
| Trichospermum mexicanum (DC.) Baill. | tme | 0 | 0 | 0 | 1 | 0 | LC | - |
| Urticaceae | ||||||||
| Urera caracasana (Jacq.) Gaudich. ex Griseb. | uca | 0 | 3 | 6 | 1 | 1 | LC | - |
| Violaceae | ||||||||
| Rinorea guatemalensis (S. Watson) Bartlett | rgu | 1 | 0 | 1 | 5 | 4 | LC | - |
| Totales (individuos) | 58 | 80 | 83 | 109 | 70 |
En cuanto al análisis de cada una de las unidades, en la UM 1 se registró un total de 58 individuos de 23 especies, pertenecientes a 23 géneros de 15 familias. La familia con mayor número de especies fue Rubiaceae con cuatro especies, las familias Anacardiaceae, Araliaceae, Fabaceae, Malvaceae y Meliaceae con dos especies cada una, las familias restantes estuvieron representadas por una. En la UM 2 se registraron 80 individuos de 27 especies, pertenecientes a 25 géneros de 19 familias. Las familias más representativas fueron Meliaceae con cuatro especies y Rubiaceae con tres, las familias Fabaceae, Malvaceae y Moraceae con dos cada una, y las demás familias con una especie respectivamente. En la UM 3 se encontró un total de 83 individuos de 24 especies, pertenecientes a 24 géneros de 15 familias. Las familias con más riqueza fueron Malvaceae con cuatro especies y Moraceae con tres. En la UM 4 se registraron 109 individuos de 39 especies, pertenecientes a 36 géneros de 24 familias. Las familias Malvaceae y Meliaceae estuvieron representadas por cuatro especies cada una, seguidas de Fabaceae, Lauraceae y Rubiaceae con tres especies, Arecaceae, Clusiaceae y Moraceae con dos y las familias restantes por una especie. Los géneros mejor representados fueron Guarea con tres especies y Nectandra con dos. En la UM 5 se registraron un total de 70 individuos de 26 especies, pertenecientes a 24 géneros de 18 familias. Las familias con mayor riqueza fueron: Lauraceae, Malvaceae y Maliaceae con tres especies cada una, seguidas de Moraceae con dos especies, las familias restantes estuvieron representadas con una. Los géneros Guarea y Nectandra estuvieron presentes con dos especies cada una, los géneros restantes se registraron con una especie cada uno (Tabla1).
La especie más abundante en las cinco UM fue A. mexicanum con el 22% del total de los individuos registrados. En la UM 1, la segunda especie más abundante fue O. mexicanum con un 8.75%. En las UM 2, 3 y 5 B. alicastrum representó el 9.25%, D. arboreus con un 5.25% en la UM 5, y en las UM 4 y 5 C. brasiliense con el 4%. De acuerdo con el estado de conservación de la IUCN, de las 59 especies, Erythrina caribaea y G. glabra se encuentran listadas como Vulnerable (VU) y 44 en Preocupación Menor (LU). En la NOM-059-SEMARNAT-2010 las especies A. graveolens, G. anómala, T. radiata y C. brasiliense con categoría de Amenazada (A), las restantes no tienen una categoría asignada.
De acuerdo con el estimador no paramétrico de Bootstrap se estimó una riqueza de especies similar a lo observado en las cinco UM, con una completitud del muestreo de hasta el 84.51% (Tabla 2). En la UM 4 se observaron los valores más altos de diversidad de acuerdo con el índice de diversidad verdadera de orden 1 (1D) y 2 (2D) con 24.28 y 16.47 respectivamente, a diferencia de la UM 1 donde se observaron los más bajos con 12.62 y 6.67. Con respecto al índice de Pielou, las cinco UM mostraron una distribución equitativa de las especies, el valor más alto se registró en la UM 5 con J’=0.90 y el más bajo en la UM 1 con J’=0.80. Al calcular el índice de similitud entre las cinco UM, se determinó que los sitios que comparten un mayor número de especies fueron la UM 4 y la UM 5 al compartir una riqueza de 25 especies con un valor de similitud del 76%, en cambio la UM 1 y la UM 4 fueron las menos similares al compartir una riqueza de 12 especies con un valor de similitud del 38% (Tabla 3).
Tabla 2 Índices de diversidad verdadera de Orden 1 (1D) y de Orden 2 (2D), estimador de Bootstrap.Table 2. True diversity indices of Order 1 (1D) and Order 2 (2D), Bootstrap estimator.
| Cuadrante | Riqueza | 1D | 2D | Equidad | Bootstrap | Completitud |
|---|---|---|---|---|---|---|
| UM 1 | 23.00 | 12.62 | 6.67 | 0.80 | 28.49 | 80.73 |
| UM 2 | 27.00 | 14.56 | 8.60 | 0.81 | 32.81 | 82.29 |
| UM 3 | 24.00 | 13.40 | 8.39 | 0.81 | 28.40 | 84.51 |
| UM 4 | 37.00 | 24.28 | 16.47 | 0.88 | 44.92 | 82.37 |
| UM 5 | 26.00 | 19.20 | 14.67 | 0.90 | 30.91 | 84.12 |
| TOTAL | 59.00 | 25.62 | 12.9 | 0.80 | 67.34 | 87.61 |
Tabla 3 Valores de similitud en las cinco Unidades Muestrales (UM).Table 3. Similarity values in the five Sampling Units (UM).
| UM 1 | UM 2 | UM 3 | UM 4 | UM 5 | |
|---|---|---|---|---|---|
| UM 1 | - | 0.44 | 0.46 | 0.38 | 0.44 |
| UM 2 | 11 | - | 0.66 | 0.54 | 0.64 |
| UM 3 | 11 | 17 | - | 0.47 | 0.60 |
| UM 4 | 12 | 18 | 15 | - | 0.76 |
| UM 5 | 11 | 17 | 15 | 25 | - |
En la UM 1 la curva de rango abundancia muestra que A. mexicanum es la especie dominante en el sitio, lo que representa el 30.48% del total. En la UM 2 las especies más dominantes fueron A. mexicanum con el 23.75% y B. alicastrum con el 20%, representaron el 43.75% del total de la abundancia. En la UM 3 A. mexicanum con el 25.30% y Oecopetalum mexicanum con 16.86%, sumando el 42.16%. En la UM 4 y la UM 5 la especie A. mexicanum fue la más dominante con el 16.51% y 14.28%, respectivamente (Figura 1).
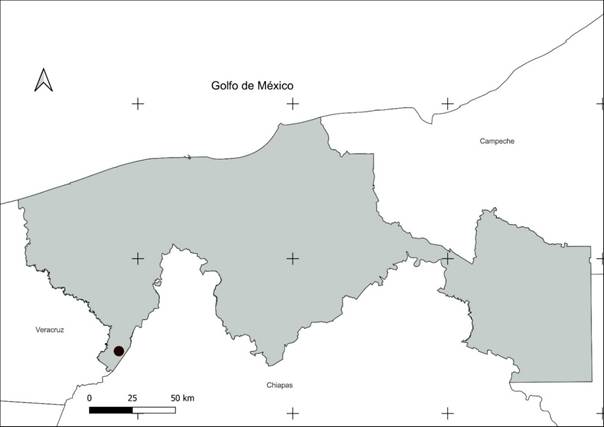
Figura 1 Mapa de ubicación del sitio de muestreo de una selva mediana, dentro del Complejo Ecoturístico “Agua Selva”, Huimanguillo, Tab. Méx.
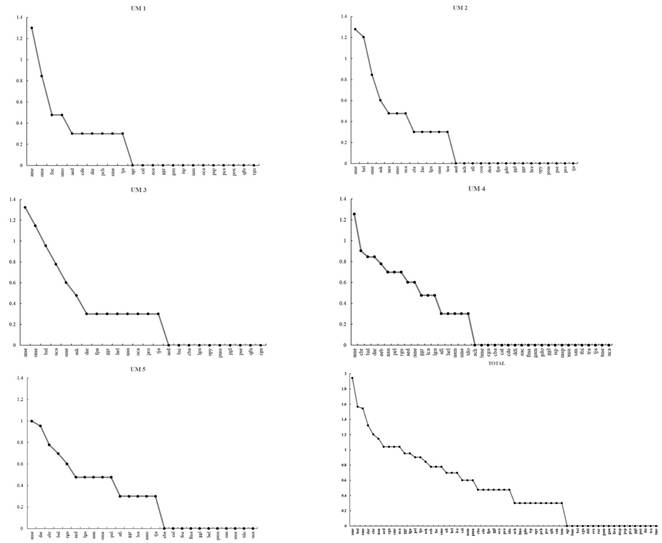
Las especies Terminalia amazonia y A. mexicanum presentaron los mayores valores de IVI con 31.55% y 27.16%, respectivamente. Por su parte, la especie O. mexicanum presentó el IVF más alto con 29.69%, seguida de B. alicastrum y A. mexicanum con 28.25% y 26.80% respectivamente. En el cálculo del IVF se tomaron en cuenta los valores relativos del diámetro, altura y cobertura de las especies. El mayor diámetro fue para O. mexicanum con 9.08%, seguido de Brosimum alicastrum, Astrocaryum mexicanum, Dendropanax arboreus con 9.41%, 8.93% y 8.83% respectivamente. La mayor altura la presentó A. mexicanum con 10.64%, seguida de B. alicastrum y O. mexicanum con 9.96% y 9.80%, respectivamente. Las especies con mayor cobertura de copa fueron O. mexicanum, B. alicastrum y Sterculia mexicana ocupando un 28.86% del área del sitio (Tabla 4).
Tabla 4 Parámetros estructurales estimados para las especies registradas en el sitio de muestreo.Table 4. Estimated structural parameters for the species recorded at the sampling site.
| Especie | Diámetro (%) |
Altura (%) |
Cobertura (%) |
IVF | Abundancia relativa (%) |
Dominancia relativa (%) |
Frecuencia relativa (%) |
IVI |
|---|---|---|---|---|---|---|---|---|
| Oecopetalum mexicanum Grenm. & C. H. Thomps | 9.08 | 9.80 | 10.81 | 29.69 | 8.75 | 4.33 | 3.92 | 17.00 |
| Brosimum alicastrum Sw. | 9.41 | 9.96 | 8.89 | 28.25 | 9.25 | 5.40 | 3.92 | 18.57 |
| Astrocaryum mexicanum Liebm. ex Mart | 8.93 | 10.64 | 7.23 | 26.80 | 22 | 1.24 | 3.92 | 27.16 |
| Dendropanax arboreus (L.) Decne. & Planch. | 8.83 | 7.79 | 7.30 | 23.92 | 5.25 | 10.03 | 3.27 | 18.55 |
| Sterculia mexicana R. Br. | 6.05 | 5.23 | 9.17 | 20.44 | 2.75 | 9.78 | 2.61 | 15.15 |
| Calophyllum brasiliense Cambess | 4.32 | 4.40 | 5.94 | 14.66 | 4 | 5.25 | 3.92 | 13.17 |
| Terminalia amazonia (J. F. Gmel.) Excell | 4.81 | 1.43 | 3.89 | 10.13 | 0.5 | 29.74 | 1.31 | 31.55 |
| Alibertia edulis (Rich.) A. Rich. ex DC. | 3.57 | 2.99 | 2.99 | 9.55 | 2.75 | 2.27 | 3.92 | 8.95 |
| Spondias mombin L. | 2.98 | 2.74 | 3.23 | 8.95 | 1.5 | 3.79 | 1.96 | 7.25 |
| Guarea grandifolia A. DC. | 3.21 | 3.15 | 2.52 | 8.88 | 2.25 | 2.58 | 3.27 | 8.09 |
IVF= Índice de Valor Forestal. IVI= Índice de Valor de Importancia. Las especies están ordenadas en orden descendente según su IVF.
Discusión
La composición de especies arbóreas en el área de estudio se agrupó mayormente en las familias Rubiaceae, Malvaceae, Fabaceae y Moraceae, dichas familias son frecuentemente reportadas en las selvas medianas perennifolias y subperennifolias debido a que están ampliamente distribuidas en la región tropical (Bremer, 2009; Juárez-Fragoso et al., 2017; Alarcón-Alarcón et al., 2019; Moreno-Jiménez et al., 2019; Gómez Velázquez et al., 2020; García-Cox et al., 2023). Este hallazgo es consistente con estudios previos en selvas tropicales, donde se ha observado que ciertas especies dominan en términos de abundancia debido a su adaptabilidad a las condiciones locales (Hubbell, 2001; Challenger y Soberón, 2008), a su plasticidad ecológica y estrategias adaptativas. En particular Fabaceae, es una familia conocida por su capacidad de fijación de nitrógeno, lo que mejora los suelos escasos en nutrientes y le da ventaja para adaptarse, en cambio hay especies de Moraceae que suelen estar asociados con interacciones mutualistas con fauna dispersora, favoreciendo su establecimiento y regeneración (Domínguez-Domínguez et al., 2006; Díaz-Prieto et al., 2017). Además, la dominancia de estas familias podría estar influenciada por factores ambientales locales, como la disponibilidad de agua y luz y la competencia interespecífica, también habría que tomar en cuenta la historia de perturbaciones en la zona. De acuerdo con Chazdon (2017), ciertas especies en ecosistemas perturbados, tienden a establecerse con mayor éxito debido a su regeneración rápida o tolerancia a condiciones de estrés.
La riqueza y la abundancia que se registró en nuestro estudio fue similar a lo reportado por Maldonado-Sánchez y Maldonado-Mares (2010) con 51 especies y una abundancia de 549 individuos en una selva tropical de Tacotalpa, Tabasco; no obstante, fueron mayores a lo registrado por Alarcón-Alarcón et al. (2019) quienes reportan una riqueza cinco veces menor en una selva de Veracruz; así mismo, Zamora-Crescencio et al. (2018, 2020) reportan una riqueza y abundancia mayor a lo registrado en nuestro estudio con 80 especies y 2 367 individuos y 79 especies con 1 528 individuos, ambos estudios en Campeche. Como se puede apreciar en los diferentes estudios el número de especies y sus abundancias puede variar notablemente, esto solo es el reflejo de la riqueza florística de las selvas mexicanas. Esto podría deberse al resultado de la combinación de factores como la interacción entre especies, la competencia por recursos, la fitofagia por parte de otros grupos taxonómicos, el tipo de suelo e incluso las restricciones fisiológicas propias de éstas. Comprender el grado y el porqué de estas diferencias es importante para interpretar los patrones de distribución de la biodiversidad, ya sea animal o vegetal.
En los muestreos realizados en el área de estudio se registró a Astrocaryum mexicanum como la especie más abundante. De acuerdo con Piñero et al. (1977), esta especie es comúnmente dominante en el estrato inferior de las selvas, alcanzando alturas considerables. La densidad y cobertura de esta especie contribuyen significativamente en la composición de la comunidad vegetal, ya que pueden influir en la regulación de la luz solar que penetra desde el dosel, la humedad y la competencia por recursos. En áreas donde esta palma es menos abundante, se observa un incremento en la diversidad de especies vegetales, especialmente en el estrato inferior. Por otra parte, en áreas donde A. mexicanum presenta abundancias superiores al 80 % los valores de diversidad tienden a ser bajos. Estos datos coinciden con nuestros registros de A. mexicanum, donde se obtuvo una abundancia promedio del 22% en cada UM, con valores de diversidad medios-altos.
Al comparar las UM, se determinó que la UM 4 es más heterogénea, lo que fue corroborado con los índices de diversidad 1D y 2D, posiblemente por variaciones microambientales que favorecen la coexistencia de más especies (Grubb, 1977), ya que en esta UM se presenta una pendiente de aproximadamente 45 grados, está inclinación puede crear condiciones desfavorables para el crecimiento de árboles de tallas grandes debido a la estabilidad limitada, mayor erosión y menor profundidad para que las raíces logren fijarse. La ausencia de estos árboles permite mayor entrada de luz, lo que facilita que otras especies de menores tallas puedan desarrollarse. Así mismo, se identificó que la mayor equidad de las poblaciones vegetales se registró en la UM 5, debido a que las poblaciones vegetales presentan abundancias similares, con pocas poblaciones dominantes. En cuanto a la similitud, el valor más alto se registró entre la UM4 y UM5, con un valor del 76%, mientras que la menor similitud fue entre la UM1 y UM4 (38%). Esto podría deberse a diferencias en las condiciones ambientales y el manejo del uso del suelo entre las diferentes unidades (Sánchez-Gutiérrez et al., 2021).
Las primeras diez especies con mayor IVF e IVI nos permiten conocer los componentes más importantes de la vegetación en el sitio. Estos índices destacan los árboles que están mejor adaptados por su dominancia, codominancia, abundancia y distribución en cada UM. En este estudio las especies con mayores índices fueron Oecopetalum mexicanum y Brosimum alicastrum, esta última también fue reportada con el mayor IVI por Martínez-Sánchez (2016) y Tadeo-Noble et al. (2024) en selvas medianas subperennifolias en Tabasco y Quintana Roo, respectivamente. En otros estudios reportan a Terminalia buceras (Zamora-Crescencio et al., 2017) y Bursera simaruba (Gómez Velázquez et al., 2020) con los índices más altos. Aunque en los sitios donde se han llevado a cabo estos estudios son selvas medianas, se pueden notar las diferencias en la riqueza de especies arbóreas que se encuentran, esto se puede deber en parte a la biología y adaptabilidad que presentan para colonizar los sitios. En el caso de T. buceras se ha reportado como una especie donde el lugar tiene características de zonas inundables, en el caso de B. simaruba es una especie que se le conoce por ser una especie típica de selvas perturbadas, pero que en el sitio estudiado se registró con valores bajos de IVI e IVF, por lo que sugiere que aún es una selva conservada. En cambio, B. alicastrum fue la segunda con los mayores IVI e IVF, esto puede ser debido a que estos árboles llegan a adaptarse a sitios con pendientes pronunciadas y es una especie de amplia distribución por todo el Golfo de México y por el Pacífico, y es común en la selva alta y mediana perennifolia de México (Pennington y Sarukhán, 2005). También tiene un papel importante en la dinámica y estructura de las selvas, ya que se ha visto que bajo los árboles masculinos se presenta una alta diversidad de plántulas sin que llegue a dominar alguna especie en particular en las primeras etapas de desarrollo. Esto sugiere que B. alicastrum es una especie que favorece el repoblamiento de otras; en cambio, bajo la cobertura de los árboles femeninos se ha presentado una baja diversidad de plántulas, alrededor del 80% de esta especie. Por lo que facilita la coexistencia de diversos árboles en el dosel. Esta característica ha favorecido que se le encuentre asociado con especies como Lonchocarpus sp., Spondias sp., Terminalia amazonia, Calophyllum brasiliense y Dendropanax arboreus (Berg, 1972; Villanueva-Ávalos, 2018), mismas especies que también fueron registradas para este estudio.
Conclusiones
La diversidad y composición de especies arbóreas en el sitio de estudio reflejan la riqueza que caracteriza a las selvas medianas perennifolias. Las familias Rubiaceae, Malvaceae, Fabaceae y Moraceae fueron predominantes en esta investigación, y que coincidieron con hallazgos previos en selvas tropicales, resaltando la adaptabilidad a las condiciones locales de cada sitio muestreado. La riqueza y la abundancia muestran variaciones significativas en comparación con otros estudios realizados en el sureste, lo que resalta la influencia de los factores microambientales y la competencia por los recursos en la distribución de las poblaciones de árboles. La abundancia de A. mexicanum en el estrato inferior y su impacto en la diversidad de especies vegetales demuestra la complejidad de las interacciones ecológicas presentes. Además, la heterogeneidad y equidad en las UM podrían estar influenciadas por las condiciones microambientales y las variaciones del terreno, lo que posiblemente favorezca la coexistencia de diversas especies. Por último, los índices de valor de importancia resaltan las especies mejor adaptadas y su rol en la estructura y dinámica del ecosistema, evidenciando la importancia de árboles como B. alicastrum para favorecer el repoblamiento y coexistencia de otras especies arbóreas.

