











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La región de la Mixteca Alta, localizada al noroeste de Oaxaca, ha sido caracterizada como de alta biodiversidad, con un alto nivel de endemismo (García-Mendoza, Tenorio y Reyes-Santiago, 1994) y una compleja heterogeneidad ambiental y cultural (Palacio-Prieto et al., 2016). No obstante, la región registra una marcada degradación histórica de la cobertura forestal y una intensa erosión del suelo (Guerrero-Arenas, Jiménez-Hidalgo y Santiago-Romero, 2010; Fernández de Castro, Vásquez, Palacio, Peralta y García, 2018) que han modificado la composición de especies y la pérdida de endemismos a escala local (Asbjornsen, Ashton, Vogt y Palacios, 2004a; Aguilar-Santelises y del Castillo, 2013). A la fecha existen pocos estudios ecológicos sobre la estructura, composición y diversidad de los principales tipos de vegetación arbórea que allí se desarrollan (ej. Aguilar-Santelises y del Castillo, 2013; Silva-Aparicio, Castro y Castillo-Campos, 2018), en comparación con otras áreas aledañas como la Reserva de Tehuacán-Cuicatlán (Secretaría de Medio Ambiente y Recursos Naturales [semarnat], 2013), los trabajos en la Mixteca Alta se han centrado en listados florísticos (García-Mendoza et al., 1994).
La confluencia de las provincias florístico-fisiográficas de la Mixteca Alta y la Depresión del Balsas (Solano, 1997) propició que la Mixteca Alta albergue más de 1550 taxa nativos, 163 endémicas y 922 exclusivas de la región (García-Mendoza et al., 1994). Sin embargo, la cobertura forestal original de la región no ha estado exenta de cambios y modificaciones, debido a actividades productivas como la agricultura y la ganadería, principalmente. Un estudio retrospectivo de Smith (1976) propone que ciertas zonas del Valle de Nochixtlán se componían de bosques de pino y encino muy cerrados, que no permitían el desarrollo de herbáceas y arbustos en el sotobosque, así como bosques de galería sobre los cauces de ríos.
Para el geoparque Mundial Unesco Mixteca Alta (GMA), se han descrito diversos tipos de vegetación reconocidos de manera general para toda la región Mixteca Alta, conformados por bosques de pino y encino, matorrales xerófilos (básicamente esclerófilos), palmares y pequeñas áreas con bosque tropical caducifolio y bosque mesófilo de montaña (García-Mendoza et al., 1994). De manera particular para el GMA, se han identificado hasta 13 tipos de vegetación tales como, bosque de encino, bosque de encino y pino, vegetación secundaria arbórea y arbustiva de bosque de encino, matorral secundario, matorral espinoso, pastizal inducido y vegetación ribereña para la cuenca de Yanhuitlán (Oropeza, Vences, Cram, Ortiz y Hermann, 2016), así como bosque de enebro, bosque de encino, bosque tropical caducifolio, matorral xerófilo, matorral subtropical y bosque de galería para una localidad del distrito de Huajuapan (Solano, 1997).
Dentro del GMA, en un estudio sobre transición forestal, se ha observado en los últimos 30 años una recuperación de 20% para la vegetación de pino y encino, y encino, y de 80% para vegetación de matorral debido a la migración, abandono parcial de la agricultura, disminución del pastoreo y regulación interna de las comunidades locales sobre el uso de los recursos (Lorenzen, Orozco-Ramírez, Ramírez-Santiago, y Garza, 2020). Sin embargo, desde el punto de vista ecológico y de diversidad, los estudios que se han llevado a cabo en el área son escasos, sobre todo considerando los diversos ambientes y tipos de vegetación que integran al GMA.
Objetivos
Este estudio tuvo como objetivo identificar los tipos de coberturas del suelo, así como caracterizar la estructura, composición y diversidad de los tipos de vegetación leñosa del GMA, desde un punto de vista florístico y fisonómico, con la finalidad de generar información que pueda ser utilizada en los futuros proyectos de restauración y conservación de las comunidades locales que integran el geoparque.
Materiales y métodos
Área de estudio
Este estudio se llevó a cabo en el territorio del GMA (Fig. 1). El cual se encuentra al noroeste de la ciudad de Oaxaca, en la región de la Mixteca Alta, posee una extensión de 415.4 km2 y fue declarado como tal en el año 2017 por la Organización de las Naciones Unidas para la Educación, la Ciencia y la Cultura [Unesco]. Lo conforman nueve municipios con dieciséis comunidades agrarias; los municipios que abarca son San Andrés Sinaxtla, San Bartolo Soyaltepec, San Juan Teposcolula, San Juan Yucuita, San Pedro Topiltepec, Santa María Chachoapám, Santiago Tillo, Santo Domingo Tonaltepec y Santo Domingo Yanhuitlán, Topográficamente se considera uno de los territorios más complejos de México, por el contacto entre la Sierra Madre del Sur y la Sierra Madre Oriental (Rosado, 2016), con un intervalo altitudinal de 2050 m a 2892 m s.n.m. El clima del área presenta tres subtipos de climas templado subhúmedo (C): el Cw0, Cw1 y el Cw2, así como un cuarto subtipo perteneciente al clima semiseco templado BS1Kw (García, 2004; Instituto Nacional de Estadística, Geografía e Informática [Inegi], 2008). La precipitación anual oscila entre 480 mm y 869 mm en el periodo 1927 a 2016 (Comisión Nacional del Agua [Conagua], 2019). Los suelos dominantes en la región corresponden a los tipos Feozem Cambisol, Castañozem, Litosol, Luvisol, Regosol y Vertisol (Inegi, 1998). El Feozem calcárico, es el dominante, se caracteriza por presentar un color pardo rojizo, de textura arcillosa, con altos contenidos de carbonato de calcio (16.9 %) (Contreras et al., 2003).
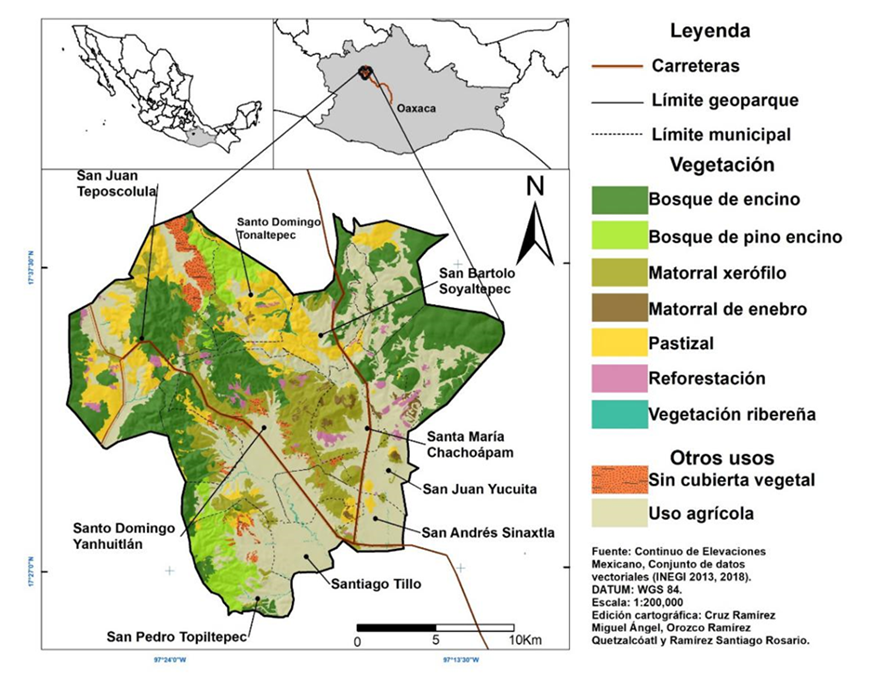
Tipos de coberturas del suelo del GMA
El mapa de uso de suelo y vegetación se construyó a partir de la interpretación visual (Lillesand, Kiefer y Chipman, 2015) realizada por expertos en la zona, utilizando como base la imagen de satélite de alta resolución (Maxar Technologies, del 19 de febrero de 2016), disponible en Google Earth. La digitalización de los tipos de cobertura se hizo en Qgis 2.18, que permite importar directamente la imagen de Google Earth georeferenciada. A cada tipo de cobertura se le asignó un tipo de vegetación con base en la clasificación de Rzedowski (2006) y en concordancia con los tipos de vegetación señalados por Smith (1976), García-Mendoza et al. (1994) y Oropeza et al. (2016). Para verificar la correcta asignación de los tipos de vegetación, se hicieron recorridos de campo. Con la información de estos recorridos, se corrigieron algunos polígonos, para asignarles el tipo de vegetación encontrada en campo.
Composición y diversidad de la vegetación leñosa del GMA
En forma paralela a los recorridos de verificación del mapa de vegetación, se llevó a cabo el muestreo de la vegetación leñosa. Para ello se utilizaron sitios temporales en forma de transectos de 10 m × 50 m (500m2) (Mostacedo y Fredericksen, 2000). Se aplicó un diseño de muestreo aleatorio simple, el número de cuadrantes varió en función de la superficie del tipo de vegetación. Todos los árboles y arbustos mayores a un metro de altura fueron medidos. Para cada individuo se tomó registro de especie, altura (m), cobertura (mediante la medición de los diámetros mayor y menor en metros) y, solo para los árboles, el diámetro a la altura del pecho (cm). En el centro de cada sitio, se registró localización geográfica, altitud, topografía, orientación, exposición, pendiente del terreno y tipo de vegetación. En total se muestrearon 70 sitios, distribuidos de la siguiente manera: 24 sitios para el bosque de encino; 22 sitios para el bosque de pino y encino; 19 sitios para la vegetación de matorral xerófilo y 5 sitios para el matorral de enebro.
Se colectaron muestras botánicas de las especies registradas en cada sitio, las cuales fueron identificadas mediante claves taxonómicas y se hicieron consultas al portal en línea del Herbario Nacional de México [MEXU], de la Universidad Nacional Autónoma de México [UNAM]. Los ejemplares se encuentran depositados en el herbario del GMA. En el caso de los ejemplares de encinos, estos fueron identificados por la Dra. Susana Valencia Ávalos de la Facultad de Ciencias de la UNAM, por lo quedaron depositados en el herbario de la Facultad de Ciencias, UNAM.
Análisis de datos
Los individuos de árboles y arbustos medidos fueron registrados en una base de datos de Excel, agrupándose por tipo de vegetación. Posteriormente, se obtuvo el índice de valor de importancia (IVI) para cada especie y tipo de vegetación para lo cual se calcularon tres variables: dominancia relativa (en este caso se utilizó la cobertura), densidad relativa y frecuencia relativa (Mostacedo y Fredericksen, 2000). Las dos mediciones de cobertura se utilizaron para obtener el diámetro de copa y posteriormente la superficie de la copa, esta última se utilizó para el cálculo de la dominancia.
Se estimó la completitud del muestreo para cada tipo de vegetación, mediante un análisis de la cobertura de la muestra. La completitud considera la proporción del número total de individuos de las especies representadas en el inventario, por lo que es considerada una medida de la integridad del muestreo (Chao y Jost, 2012). Los valores de completitud varían de 0 (mínima completitud) a 1(máxima completitud) (Chao y Jost, 2012; González-Tokman, Cultid, Díaz, Ocampo y Martínez, 2018), donde valores mayores a 0.5 son considerados dentro de un inventario aceptable (Hsieh, Ma y Chao, 2016; López-Segoviano, Díaz-Verdugo, Arenas-Navarro y Arizmendi, 2019). Ambos análisis se realizaron con el paquete “iNEXT” 2.0.5 (Hsieh et al., 2016) y “vegan” v. 2.3.-2 (Oksanen, et al., 2015) en el software R versión 3.0.3 (R Core Team, 2014), con intervalos de confianza de 95%.
La diversidad alfa de cada tipo de vegetación se estimó como el número de especies efectivas (Hill, 1973; Jost, 2006; Moreno, Barragán, Pineda y Pavón, 2011). Donde la diversidad de orden cero (q = 0, Riqueza de especies) es igual a la riqueza de especies, la diversidad del orden 1 (q = 1, Shannon) pesa proporcionalmente todas las especies de acuerdo con su abundancia en la comunidad y la diversidad de orden 2 (q = 2, Simpson) da más peso a las especies dominantes.
Resultados
Tipos de coberturas del suelo del GMA
Siguiendo la clasificación de Rzedoswki (2006) para la vegetación, se identificaron nueve tipos de vegetación y dos tipos de coberturas en el GMA (Fig. 1). De estos, el de mayor extensión es el uso agrícola, que cubre 154 km2 y representa 37% de la superficie del geoparque; el bosque de encino es el segundo tipo de cobertura más extenso y cubre una superficie de 101 km2 (24%); el matorral xerófilo ocupa un superficie de 77 km2 (18.6 %), siendo el tercero en extensión; le sigue el pastizal que ocupa una superficie de 39 km2 (9%); el bosque de pino y encino que cubre 20 km2 (5%); y el matorral de enebro que ocupa una superficie reducida, de apenas 4 km2 (1%).
Se identificaron otros tipos de vegetación y coberturas, como la vegetación ribereña, reforestaciones de Pinus sp. y áreas sin cubierta vegetal, que ocupan superficies reducidas y no superan, en conjunto, 6% de la superficie total del geoparque. Cabe destacar que la superficie sin cubierta vegetal solo representa 3% del área de GMA.
Composición y diversidad de la vegetación leñosa del GMA
En relación con el muestreo efectuado, los valores de completitud para cada tipo de vegetación fueron satisfactorios, ya que se obtuvieron valores por arriba de 0.88 en bosque de encino, pino-encino, matorral xerófilo y matorral de enebro; obteniendo el valor más bajo el bosque de encino con 0.88 y el más alto el matorral xerófilo con 0.96, mientras la cobertura de la muestra total para el área fue de 97% de las especies posibles (Fig. 2). Un listado completo de las especies registradas en este estudio se incluye en el suplemento 1.
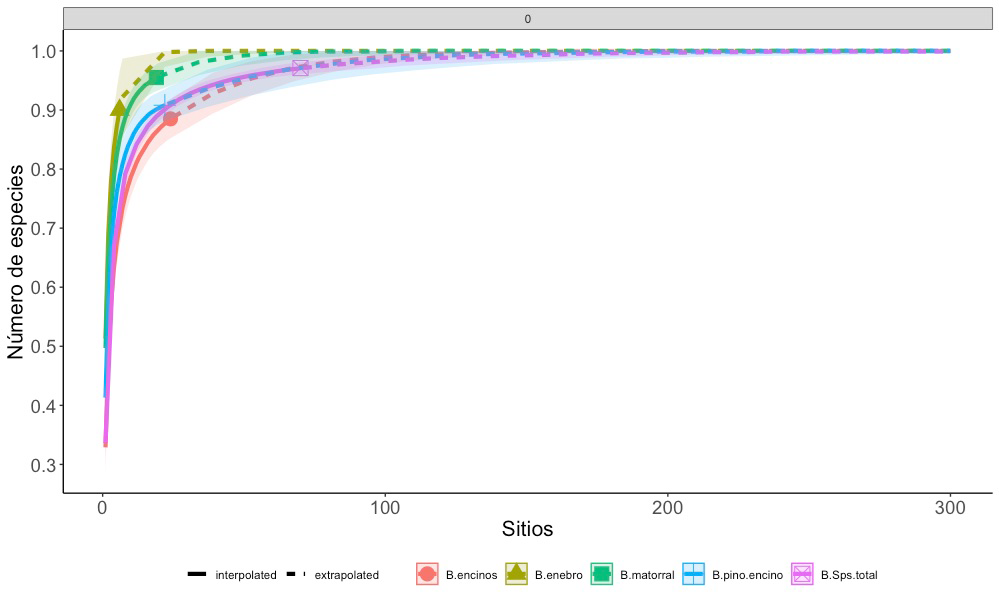
Las bandas corresponden al intervalo de confianza a 95%.
Figura 2 Análisis de la cobertura de la muestra basado en el número de individuos recolectados en cuatro tipos de vegetación del geoparque Mixteca Alta (encino, pino y encino, matorral y enebro), así como el total del inventario (B.Sptotal).
En total se registraron 31 familias, distribuidas en 55 géneros y 91 especies, de las cuales 62 son arbustos, 28 corresponden a especies de árboles y una especie de palma del género Brahea. Las familias mejor representadas en el GMA, en términos de abundancia relativa son Fagaceae (22%), Compositae (10%) y Ericaceae (9.8%), Rosaceae (8%), Sapindaceae (8%) y Pinaceae (7%). Las tres primeras, son las que registraron el mayor número de especies (Suplemento 1). Además, se registró que 20% de las especies están presentes en los cuatro tipos de vegetación. De estas especies las más representativas por su dominancia son: Juniperus fláccida, Pinus oaxacana, Comarostaphylis polifolia, Dodonaea viscosa, Cercocarpus fothergilloides, Rhus standleyi, Quercus liebmannii, Garrya laurifolia, Malacomeles denticulata, Ageratina tomentella y Archibaccharis asperifolia (Tabla 1; Suplemento 1).
Tabla 1 Índice de valor de importancia para las especies de árboles y arbustos (> 1 m de altura) de mayor dominancia en los tipos de vegetación leñosa del geoparque Mixteca Alta.
| Tipo de vegetación | Forma de vida | Especie | IVI % |
| Bosque de encino | Árbol | Quercus rugosa Née | 68.205 |
| Bosque de encino | Árbol | Quercus castanea Née | 31.506 |
| Bosque de encino | Árbol | Quercus liebmannii Oerst. ex Trel. | 21.440 |
| Bosque de encino | Árbol | Quercus deserticola Trel. | 19.197 |
| Bosque de encino | Árbol | Quercus crassifolia Bonpl. | 14.385 |
| Bosque de encino | Árbol | Arbutus xalapensis Kunth | 11.452 |
| Bosque de encino | Árbol | Comarostaphylis polifolia (Kunth) Zucc. ex Klotzsch | 11.407 |
| Bosque de encino | Árbol | Juniperus flaccida Schltdl. | 10.546 |
| Bosque de encino | Arbusto | Archibaccharis asperifolia (Benth.) S.F.Blake | 9.187 |
| Bosque de encino | Arbusto | Ageratina tomentella (Schrad.) R.M.King & H.Rob. | 8.928 |
| Bosque de pino y encino | Árbol | Pinus oaxacana Mirov | 72.658 |
| Bosque de pino y encino | Árbol | Juniperus flaccida Schltdl. | 22.471 |
| Bosque de pino y encino | Árbol | Quercus rugosa Née | 19.344 |
| Bosque de pino y encino | Arbusto | Dodonaea viscosa (L.) Jacq. | 16.145 |
| Bosque de pino y encino | Arbusto | Garrya laurifolia Benth. | 16.039 |
| Bosque de pino y encino | Árbol | Quercus liebmannii Oerst. ex Trel. | 15.268 |
| Bosque de pino y encino | Árbol | Quercus deserticola Trel. | 14.289 |
| Bosque de pino y encino | Árbol | Comarostaphylis polifolia (Kunth) Zucc. ex Klotzsch | 11.820 |
| Bosque de pino y encino | Arbusto | Rhus standleyi F.A. Barkley | 10.189 |
| Bosque de pino y encino | Árbol | Arbutus xalapensis Kunth | 8.840 |
| Matotorral xerófilo | Árbol | Comarostaphylis polifolia (Kunth) Zucc. ex Klotzsch | 35.438 |
| Matotorral xerófilo | Árbol | Cercocarpus fothergilloides Kunth | 23.958 |
| Matotorral xerófilo | Arbusto | Dodonaea viscosa (L.) Jacq. | 20.944 |
| Matotorral xerófilo | Árbol | Juniperus flaccida Schltdl. | 20.815 |
| Matotorral xerófilo | Arbusto | Rhus standleyi F.A. Barkley | 19.894 |
| Matotorral xerófilo | Arbusto | Garrya laurifolia Benth. | 17.100 |
| Matotorral xerófilo | Árbol | Quercus grahamii Benth. | 14.603 |
| Matotorral xerófilo | Arbusto | Malacomeles denticulata (Kunth) G.N.Jones | 12.775 |
| Matotorral xerófilo | Palma | Brahea sp. | 11.462 |
| Matotorral xerófilo | Árbol | Quercus liebmannii Oerst. ex Trel. | 11.046 |
| Matorral de enebro | Árbol | Juniperus flaccida Schltdl. | 58.715 |
| Matorral de enebro | Árbol | Cercocarpus fothergilloides Kunth | 31.680 |
| Matorral de enebro | Arbusto | Dodonaea viscosa (L.) Jacq. | 30.113 |
| Matorral de enebro | Arbusto | Rhus standleyi F.A. Barkley | 22.831 |
| Matorral de enebro | Árbol | Comarostaphylis polifolia (Kunth) Zucc. ex Klotzsch | 21.343 |
| Matorral de enebro | Arbusto | Malacomeles denticulata (Kunth) G.N.Jones | 18.385 |
| Matorral de enebro | Arbusto | Salvia semiatrata Zucc. | 17.128 |
| Matorral de enebro | Árbol | Pinus oaxacana Mirov | 15.369 |
| Matorral de enebro | Arbusto | Mimosa sp. | 12.073 |
| Matorral de enebro | Arbusto | Archibaccharis asperifolia (Benth.) S.F.Blake | 10.696 |
El orden de las especies es de acuerdo con su valor de IVI.
A escala de especies, en los cuatro tipos de vegetación (bosque de encino, bosque de pino y encino, matorral de enebro y matorral xerófilo), el estrato arbustivo (48, 38, 18 y 26 especies, respectivamente) fue más rico que el estrato arbóreo (23, 19, 11 y 16 especies respectivamente). Sin embargo, a escala de IVI, los árboles contribuyeron más, que los arbustos en el bosque encino (71% y 29%, respectivamente) y bosque pino-encino (63.1% y 36.9%, respectivamente), aspecto contrario en el matorral de enebro (48.8% y 51.2%, respectivamente) y matorral xerófilo (46.7% y 53.3%, respectivamente) donde la contribución de los arbustos fue ligeramente mayor (Tabla 1).
En el bosque de encino, son precisamente las especies del género Quercus las que tuvieron la mayor contribución al IVI, siendo Quercus rugosa la de mayor aportación (68%), mientras que en arbustos fueron las especies A. asperifolia (9%) y A. tomentella (8%). En cambio, en el bosque de pino-encino, la aportación de Quercus al IVI fue mucho menor y P. oaxacana es la especie con mayor contribución (72%.), mientras que en los arbustos fue D. viscosa (16.3%). En el matorral de enebro sobresalen entre los árboles J. flaccida y C. fothergilloides, con una contribución al IVI de 58% y 31%, respectivamente, mientras que, a nivel arbustivo, sobresalieron D. viscosa y R. standleyi con un aporte de 30% y 22%, respectivamente. Finalmente, en el matorral xerófilo las especies arbóreas con mayor contribución al IVI, fueron C. polifolia (35%) y C. fothergilloides (23%), y en el arbustivo fueron las mismas especies que en el matorral de enebro, D. viscosa (20%) y R. standleyi (19%) (Tabla 1). Los valores de IVI para todas las especies registradas se incluyen en el suplemento 1.
Al comparar la riqueza efectiva de especies entre tipos de vegetación, con referencia a la abundancia total de especies de árboles y arbustos del área de estudio (91 especies), el número efectivo de especies (q=0) fue de 71 para el bosque de encino, 57 para el pino-encino, y significativamente diferente con respecto a matorral xerófilo (43) y al matorral de enebro (29) (Fig. 3.0). La diversidad de especies medianamente abundantes, que corresponde al exponente de la diversidad de Shannon (q=1), fue de 49 y 40 para bosque de encino y pino-encino, difiriendo significativamente del matorral xerófilo y el matorral de enebro, que presentaron 29 y 22, respectivamente (Fig. 3.1). De igual manera, los valores de q = 2 (el inverso del índice de Simpson) muestran que el matorral xerófilo y matorral de enebro presentaron significativamente menor número de especies dominantes (25), con respecto al bosque de encino (34) y al de pino-encino (29) (Fig. 3.2).
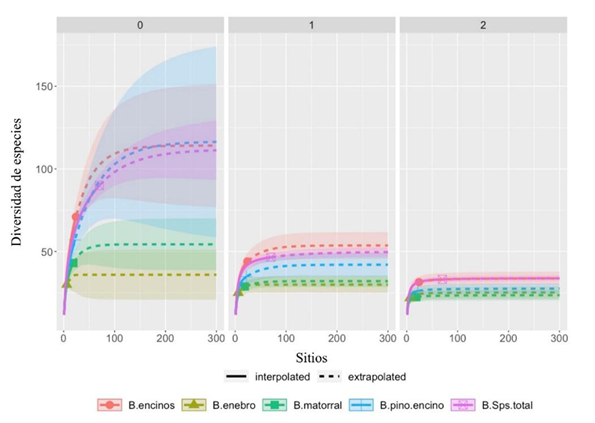
Las bandas corresponden al intervalo de confianza a 95%.
Figura 3 Curvas de interpolación (rarefacción)/extrapolación de la diversidad de especies arbóreas y arbustivas en cuatro tipos de vegetación leñosa del geoparque Mixteca Alta (encino, pino y encino, matorral y enebro), con base en los números de Hill: a) q=0: riqueza (número efectivo de especies), b) q=1: exponente de la diversidad Shannon, c) q=2: inverso del índice de diversidad de Simpson. B.Sptotal es de referencia al acumulado total de especies que presentó la región.
Discusión
Tipos de coberturas del suelo del GMA
Los tipos de vegetación señalados en este estudio coinciden con los mencionados para la región Mixteca por García-Mendoza et al. (1994), por Torres-Colín (2004),) y por Oropeza et al. (2016). Su presencia y desarrollo están relacionados con las condiciones geológicas, topográficas y climáticas de la región, pero sobre todo han estado sujetos a cambio, por las actividades humanas, que se llevan a cabo por parte de las comunidades locales (García-Mendoza et al., 1994; Solano, 1997). Lo que provocó que, en décadas pasadas, gran parte de la vegetación fuera eliminada en la región de la Mixteca, incluida el área de estudio del GMA (Smith, 1976; García-Mendoza et al., 1994; Solano, 1997; Oropeza et al., 2016).
No obstante, en las últimas décadas, esta región ha tenido cambios en el uso del suelo, como consecuencia del abandono de la agricultura debido a la migración de la población, así como a la modificación en las actividades productivas en los medios de vida de los pobladores y a la regulación sobre el acceso y uso de los recursos naturales, que ha conducido a un incremento en la superficie que ocupa actualmente la vegetación leñosa (Lorenzen et al., 2020), lo cual es ratificado en este estudio.
Aunque la agricultura es la cobertura dominante del suelo, el incremento de la vegetación se ve reflejado en los bosques de encino y matorral xerófilo que, sumados, representan 42.6% de la cobertura vegetal dentro del GMA y, en menor medida, también contribuye la cobertura del bosque de pino y encino, así como la del matorral de enebro.
La sucesión secundaria ha sido el factor clave en el aumento de la cobertura leñosa, en comparación con otros mecanismos de restauración como las reforestaciones locales de Pinus sp., como lo confirman las características de composición y estructura registrados en este estudio para el GMA, y en concordancia con lo expuesto por Oropeza et al. (2016) y Lorenzen et al. (2020). Por lo que, de acuerdo con Almazán, Corcuera, Parra, Jiménez y Charre (2016) y Zakkak et al. (2018), el mecanismo de restauración mediante sucesión secundaria debería ser considerado de forma más amplia ya que, como Uriarte y Chazdon (2016) sugieren, este mecanismo presenta ventajas no solo ecológicas, sino también económicas y sociales para las poblaciones locales y bien puede aplicarse en combinación con la reforestación con especies nativas.
Composición y diversidad de la vegetación leñosa del GMA
Con respecto al muestreo efectuado, los valores de completitud obtenidos en los cuatro tipos de vegetación (> 0.8) sugieren un buen esfuerzo de muestreo (Hsieh et al., 2016; López-Segoviano et al. 2019), sin embargo, como indican López-Segoviano et al. (2019), ello no implica que no se puedan registrar nuevas especies, en estudios futuros fuera de los puntos de muestreo.
En cuanto a la composición florística, las familias de mayor abundancia relativa coinciden con las registradas para otras áreas de bosque templado de México (Vásquez, Clark, Manzano, González y Aguirre, 2018; Zacarías y del Castillo, 2010; Aguilar-Santelises y del Castillo, 2013; Almazán et al., 2016). En el caso particular del GMA, la composición florística está caracterizada por la dominancia de especies que han sido consideradas en otros estudios como pioneras o adaptadas a ambientes perturbados (P. ej. C. polifolia, C. fothergilloides, D. viscosa, J. fláccida, Rhus sp., P. Oaxacana, Q. rugosa, Q. liebmanii, entre otras) (Smith, 1976; García et al., 1994; Asbjornsen, Vogt y Ashton, 2004b; Acosta, González y Ruacho, 2014; Almazán et al., 2016; Oropeza et al., 2016), así como por la dominancia de formas de vida arbustiva. Rasgos que, de acuerdo con Bongers, Poorter, Hawthorne y Sheil (2009), son representativos de estadios sucesionales tempranos.
Aunque los tipos de vegetación comparten la composición florística caracterizada por especies pioneras, estos difieren entre sí, en cuanto a la especie y a la forma de vida dominante. Resultados que concuerdan con lo señalado por Almazán et al. (2016), quienes indican que las especies pioneras presentes en los estadios sucesionales tempranos también pueden estar presentes en los estadios subsecuentes y promover el establecimiento de especies sucesionales tardías. En el caso del GMA, la vegetación de matorral (matorral xerófilo y matorral de enebro) ha sido considerada una fase sucesional derivada de la eliminación del bosque de encino (Oropeza et al., 2016), lo que confirma los resultados de este estudio.
Por otro lado, la riqueza de especies de los tipos de vegetación leñosa, en general, es baja en comparación con otras áreas de bosque templado del país (Valenzuela-Núñez y Granados-Sánchez, 2009; Aguilar-Santelises y del Castillo, 2013; Almazán et al., 2016; Ramírez-Santiago et al., 2019). Lo anterior puede ser atribuido a factores ambientales y antrópicos. Como Torres (2004) señala, los bosques templados del interior del estado de Oaxaca, como en el caso de la Mixteca Alta, suelen presentar una menor riqueza de especies debido a una menor incidencia de humedad y precipitación (Torres, 2004), así lo señalan también Zacarías y del Castillo (2010) para bosques templados ubicados en regiones de sotavento de Oaxaca. Aunque también influye el uso histórico del suelo que la región ha tenido (tala, pastoreo, uso agrícola) y que ha propiciado la pérdida de diversidad (García-Mendoza et al., 1994; Aguilar-Santelises y del Castillo, 2013).
No obstante lo anterior, cabe destacar que en cuanto a la diversidad de especies (Shannon y Simpson) registrada en los tipos de vegetación estudiados existe una mayor diversidad en los bosques de encino y pino-encino, en comparación con el matorral xerófilo y con el matorral de enebro, lo que puede ser resultado de una mayor estabilidad de las especies, debido a un menor impacto de actividades humanas y a un estadio sucesional más avanzado (Almazán et al., 2016), lo que permite la presencia de un mayor número de especies arbóreas, sobre todo encinos y algunas coníferas, que contribuyen a aumentar la diversidad vegetal (Estrada et al., 2012). Esto se ve reflejado en los valores del IVI, donde las especies arbóreas tienen una mayor contribución, caso contrario al matorral xerófilo y al matorral de enebro, donde sobresalen especies arbustivas, lo cual es una característica en bosques templados sometidos a procesos de perturbación antrópica como la agricultura, el pastoreo y la tala (Encina et al., 2009; González et al., 2009; Aguilar-Santelises y del Castillo, 2013; Granados, Giné y García, 2014).
Conclusiones
La vegetación arbórea del GMA es de tipo secundario, está caracterizada por una composición florística dominada por especies pioneras y con una riqueza comparativamente baja en relación con otros bosques templados. Los tipos de vegetación del GMA revelan que existe una recuperación de la cobertura vegetal mediante el mecanismo de sucesión secundaria.
El conocimiento de las características de las áreas de vegetación leñosa del GMA puede contribuir con información que enriquezca las acciones de reforestación y proyectos de conservación locales. Por ejemplo, las características de la vegetación leñosa aportan información sobre las especies de árboles y arbustos que podrían utilizarse para llevar a cabo reforestaciones en las áreas más degradadas del GMA.
Las comunidades que integran el geoparque han llevado a cabo acciones que han contribuido al mantenimiento de la cobertura vegetal. No obstante, aún se requiere de más investigaciones sobre las características de la vegetación, que incluya las especies de herbáceas y los procesos de sucesión secundaria que tienen lugar en el GMA, así como el papel de los factores ambientales y de las actividades humanas, principalmente del pastoreo, sobre estas áreas.

