










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo. Serie horticultura
versión On-line ISSN 2007-4034versión impresa ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.14 no.1 Chapingo ene./abr. 2008
Efecto de la nutrición nítrica y sistemas de riego en el sabor de la fresa (Fragaria x ananassa Duch.)
Nitric nutrition, and irrigation systems effects on strawberry (Fragaria x ananassa Duch.) flavor
L. A. Ojeda–Real1, R. Cárdenas–Navarro2, P. Lobit2, O. Grageda–Cabrera3, E. Valencia–Cantero1 y L. Macías–Rodríguez1*
1 Instituto de Investigaciones Químico–Biológicas, Universidad Michoacana de San Nicolás de Hidalgo. Francisco J. Mújica s/n, Ciudad Universitaria. Morelia, Michoacán. México. Correo–e: lmacias@zeus.umich.mx (*Autor responsable).
2 Instituto de Investigaciones Agropecuarias y Forestales. Universidad Michoacana de San Nicolás de Hidalgo. Francisco J. Mújica s/n, Ciudad Universitaria. Morelia, Michoacán. México.
3 Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Celaya, Guanajuato. México.
Recibido: 14 de febrero, 2006
Aceptado: 30 de agosto, 2007
Resumen
El sabor de la fresa está dado por el contenido de azúcares, aminoácidos y ácidos orgánicos en el fruto. En la actualidad se desconoce el efecto de la nutrición nítrica y el sistema de riego en cada uno de los compuestos saborizantes de la fresa. El objetivo de este trabajo fue identificar y cuantificar el contenido de carbohidratos, aminoácidos y ácidos orgánicos en la fresa sometida a distintos niveles de fertilización nítrica y a dos sistemas de riego. El N fue aplicado como Ca(NO3)2 a una dosis de 0, 3, 9 y 20 mM y se emplearon dos sistemas de riego: goteo y gravedad. El contenido de carbohidratos y aminoácidos se incrementó en los niveles de 3 mM para riego por goteo y 9 mM en gravedad, ambos respecto al tratamiento no fertilizado y fertilizado con 20 mM de N. El contenido de los ácidos orgánicos disminuyó su concentración al aumentar el nivel de fertilización nitrogenada. Así que el uso excesivo de fertilizante y agua no se justifica para aumentar el contenido de los compuestos relacionados con la calidad sensorial de la fresa.
Palabras clave: carbohidratos, aminoácidos, ácidos orgánicos, cromatografía de gases.
Abstract
The strawberry fruit flavor depends on its sugar, amino acid, and organic acid content. Up to now, the effect of nitric nutrition and irrigation systems on these compounds is not well characterized. The objective of this work was to identify and to quantify the carbohydrates, amino acids, and organic acids content in strawberries cultivated at different levels of nitric fertilization, under two irrigation systems. Nitrogen was applied as Ca(NO3)2 at concentrations of 0, 3, 9 and 20 mM in the irrigation water and drip and furrow irrigation systems were used. The carbohydrate and the amino acid content increased at 3 mM in drip system and 9 mM in the furrow system, compared with the unfertilized treatment, but did not increase further at 20 mM nitrogen. The organic acids content decreased as the nitrogen fertilization increased. Therefore, an excessive use of fertilizer and irrigation water does not enhance the content of the compounds related with the sensorial quality of the strawberry.
Key words: carbohydrates, amino acids, organic acids, gas chromatography.
INTRODUCCIÓN
La fresa es una de las frutas más populares y con mayor demanda a nivel mundial. La calidad sensorial de la fresa está basada principalmente en su apariencia (tamaño, forma, color, entre otros) y el balance apropiado de compuestos volátiles y no volátiles presentes en el fruto; de estos últimos los carbohidratos, aminoácidos y los ácidos orgánicos son considerados como los compuestos responsables del sabor y además son precursores de los compuestos que denotan el aroma en la fresa (Shamaila et al., 1992; Zabetakis y Holden, 1997).
En el caso de los carbohidratos, la glucosa, fructosa y sacarosa están relacionados directamente con la percepción del sabor dulce (Macías–Rodríguez et al., 2002) y la glucosa da origen a diversos compuestos como los pertenecientes a la familia de las furanonas que forman parte del aroma de la fresa.
Los aminoácidos, aunque constituyen uno de los pilares fundamentales del funcionamiento bioquímico de las plantas, son también donadores de nitrógeno (N) orgánico para la formación de diversas moléculas biológicas; tal es el caso de los compuestos volátiles. Pérez et al. (1992), afirmaron que la diversidad en el tipo de ésteres encontrados en el perfil aromático de la fresa se debía a la composición de aminoácidos y eso explica la diferencia tan marcada en el sabor y aroma aún entre variedades.
Cordenunsi et al. (2002) mencionaron que el ácido cítrico contribuye con un 92% a la acidez de la fresa, por lo que ésta se establece en términos de porcentaje de ácido cítrico. Otro ácido orgánico importante para establecer la calidad en la fresa es el ácido ascórbico también conocido como vitamina C y que además está asociado con la salud de los consumidores (Pérez et al., 1997).
Monroy et al. (2002), revelaron la importancia del N en el cultivo de la fresa, ya que es esencial para el buen crecimiento y desarrollo de la planta, principalmente en lo correspondiente a las etapas vegetativa y de floración. Las dosis de fertilización nitrogenada recomendada para este cultivo, oscilan entre 110 y 450 kg·ha–1 (Hancock, 1999). En Michoacán se llegan a aplicar dosis superiores a 600 kg de N·ha–1 con la finalidad de incrementar el rendimiento, lo que genera un impacto ambiental negativo, ya que la eficiencia de absorción de N por el cultivo de fresa es menor al 30% de las dosis aplicadas y el nitrógeno no absorbido contamina la atmósfera y el agua.
La fresa es una planta con una alta demanda de agua (Mass, 1987) lo que la hace sensible al estrés hídrico. En estas condiciones, los productores prefieren aplicar el riego en exceso para evitar periodos de deficiencia hídrica, pero la alta humedad puede ocasionar principalmente la lixiviación y pérdidas gaseosas de nutrimentos. La forma tradicional de riego para este cultivo en el estado de Michoacán es por gravedad, lo que ocasiona el desperdicio de agua y una reducción de fertilidad en el suelo. Estos problemas han impulsado a la búsqueda de nuevas formas de suministro, tiempos adecuados de aplicación y concentración de fertilizantes, además de sustituir el sistema de riego tradicional por sistemas de goteo, con la finalidad de incrementar la eficacia en la producción y elevar la calidad de este fruto.
El objetivo del presente trabajo fue determinar el efecto de la fertilización nítrica y sistemas de riego en el cultivo de la fresa sobre el contenido de carbohidratos, aminoácidos y ácidos orgánicos en el fruto.
MATERIALES Y MÉTODOS
Se trabajó con frutos de fresa (Fragaria x ananassa cv. Aromas) cosechados bajo un diseño experimental de parcelas divididas en un arreglo de bloques completos al azar con tres repeticiones. Los tratamientos de la parcela mayor fueron: riego por goteo y riego por gravedad. Los tratamientos de las subparcelas se aplicaron a diferentes dosis de nitrato de calcio en el agua de riego; de tal manera que tomando en cuenta la cantidad total de agua aplicada a lo largo del ciclo de cultivo se aplicaron las concentraciones totales siguientes: 0 mM, 3 mM (261 kg N·ha–1), 9 mM (783 kg N·ha–1) y 20 mM (1,740 kg N·ha–1). La aplicación de los tratamientos evaluados en el riego por goteo fue de acuerdo a un calendario preestablecido de demanda de agua por la planta y para el riego por gravedad se realizó un fraccionamiento de la dosis total de N en diez aplicaciones. A cada tratamiento se le asignó un área de 18 m2 (tres surcos) con una separación entre surcos de 0.85 m y 7.0 m de largo. Los datos experimentales se tomaron del surco central. El fruto seleccionado para análisis se cosechó en dos fechas diferentes: el 28 de noviembre de 2003 y 5 de marzo de 2004, fechas correspondientes a los picos de producción de la planta. El material se liofilizó, se homogenizó y se almacenó a –20 °C hasta su análisis. Los carbohidratos y aminoácidos se extrajeron con etanol al 80% v/v. El extracto crudo se dividió en dos, en la primer alícuota se derivaron los carbohidratos para formar aldonitrilos per–acetilados con cloruro de hidroxilamina en piridina y anhídrido acético; los aminoácidos y ácidos orgánicos se purificaron a partir de la segunda alícuota, utilizando una columna de intercambio catiónico de tamaño de malla de 200–400; posteriormente se derivaron con cloruro de acilo en metanol y anhídrido acético, para así ser analizados por cromatografía de gases acoplado a un detector de ionización de flama (CG–FID). Para el análisis por CG–FID se empleó una columna HP5 de 30m x 25mm con las condiciones siguientes: para carbohidratos la temperatura inicial del horno fue de 150 °C por 3 min, con incrementos de 6 °C·min–1, hasta una temperatura final de 270 °C que se mantuvo por 25 min; las temperaturas del inyector y detector fueron 230 y 250 °C, respectivamente; la presión fue de 5 psi y el flujo de la columna de 1 ml·min–1; para separar los aminoácidos y ácidos orgánicos la temperatura inicial del horno fue de 80 °C por 3 min con incrementos de 6 °C·min–1, hasta una temperatura final de 230 °C que se mantuvo por 40 min, las temperaturas del inyector y detector fueron 180 y 250 °C, respectivamente; la presión fue de 10 psi y el flujo de la columna de 4 ml·min–1. Tanto los carbohidratos como los aminoácidos y ácidos orgánicos fueron identificados por comparación con los tiempos de retención de estándares puros y cuantificados utilizando persitol y norvalina como estándares internos, respectivamente. Los datos se analizaron de manera independiente para cada fecha siguiendo un procedimiento estándar de análisis de varianza y la diferencia significativa entre medias se determinó a P≤0.05 por la prueba de la diferencia mínima significativa (DMS) (SAS Institute, 1988).
RESULTADOS
Carbohidratos
La glucosa y fructosa fueron los carbohidratos más abundantes en los frutos analizados. Además, se identificó la presencia de ribosa, arabinosa, xilosa, manosa galactosa y myo–inositol.
En la Figura 1 se muestra el contenido de carbohidratos libres totales en los dos picos de producción. Los resultados de la primera fecha analizada mostraron que la mayor acumulación de carbohidratos libres fue en los niveles de 3 y 9 mM (135.88 y 131.42 mg·g–1 p.s., en riego por goteo; 127.33 y 129.32 mg·g–1 p.s., en riego por gravedad) siendo significativo sólo para el riego por gravedad con respecto al nivel de 0 mM de nitrógeno. En lo que respecta a la segunda fecha, en el riego por goteo se observó la máxima acumulación en el nivel de 9 mM (116.98 mg·g–1 p.s.) sin ser significativa, mientras que para el riego por gravedad se observó lo contrario ya que en los niveles de 3 y 9 mM la concentración de carbohidratos libres totales tiende a ser menor que en los niveles de 0 y 20 mM.

En la Figura 2 se muestran los datos obtenidos de la cuantificación de la fructosa. El análisis estadístico de la cosecha del 28 de noviembre mostró que hubo una acumulación significativa de fructosa en 3 mM en riego por goteo y en 9 mM en riego por gravedad (94.16 y 90.49 mg·g–1 p.s., respectivamente) con respecto a los niveles 0 y 20 mM. En la fecha del 5 de marzo en riego por goteo el contenido de fructosa se incrementó de manera no significativa en la dosis 9 mM (79.04 mg·g–1 p.s.), que también tendió a disminuir en el nivel de 20 mM de N. En el caso de riego por gravedad se observó que la fructosa tendió a acumularse en los niveles 0 y 20 mM.

En lo que respecta a la glucosa, en la fecha del 28 de noviembre, en ambos sistemas de riego hubo diferencias mínimas significativas en el contenido de glucosa en las dosis de 3 y 9 mM respecto a 0 mM de nitrato. En la fecha del 5 de marzo se observó que el contenido de glucosa fue similar en ambos sistemas de riego y sólo en riego por gravedad se observó una tendencia a incrementarse con el nivel de fertilización de 20 mM de N (Figura 3).

En el contenido de pentosas totales, como componentes importantes durante el desarrollo de la planta, en los frutos cosechados el 28 de noviembre se observó una acumulación en el nivel de 3 mM en ambos sistemas de riego, siendo significativa en el caso de riego por gravedad, con respecto a 9 y 20 mM de nitrato (Cuadro 1). La ribosa presentó diferencias mínimas significativas en los frutos cultivados en ambos sistemas de riego y en la primera fecha evaluada, alcanzó la mayor concentración en el nivel de 3 mM. Para la arabinosa se observó una tendencia no significativa a alcanzar su mayor concentración en los niveles de 9 o 3 mM. La concentración de xilosa en riego por goteo alcanzó su máximo en 20 mM, siendo significativa sólo para el 28 de noviembre; mientras que para riego por gravedad la acumulación tendió a ser mayor en 9 y 3 mM para el primer y segundo muestreo, respectivamente.

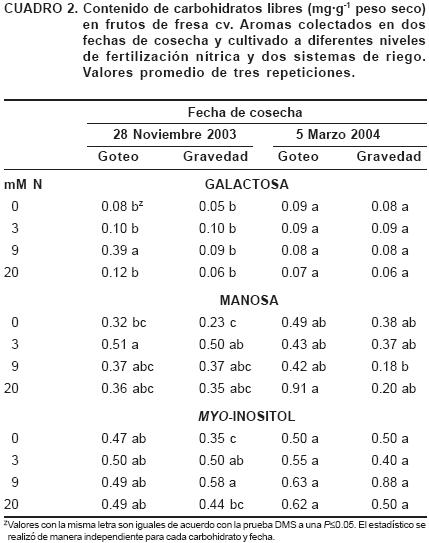
En el Cuadro 2 se muestran los resultados obtenidos en la cuantificación del resto de los carbohidratos identificados en los frutos cosechados en las dos fechas seleccionadas. En la primera fecha analizada, la manosa se acumuló significativamente en 3 mM respecto a 0 mM en ambos sistemas de riego; la concentración de galactosa aumentó significativamente en 9 mM, respecto a 0, 3 y 20 mM en riego por goteo y en riego por gravedad la concentración sólo tendió a aumentar en 3 mM; mientras que en el caso de myo–inositol sólo hubo una acumulación mínima significativa en 9 mM respecto a 0 mM en riego por gravedad. Para el muestreo del 5 de marzo, la concentración de galactosa sólo tendió a ser menor en el nivel de 20 mM. La concentración de manosa en riego por goteo fue mayor en 20 mM, pero no así en riego por gravedad; donde se acumuló en 0 mM; en el caso del myo–inositol la concentración tendió a ser mayor en el nivel de 9 mM en ambos sistemas de riego.
Aminoácidos
Al igual que en la mayoría de las plantas, el glutamato/ glutamina (glu/gln) y aspartato/asparagina (asp/asn), fueron los compuestos más abundantes en los materiales analizados. Otros aminoácidos como glicina, alanina, valina, leucina, isoleucina, treonina, serina, fenilalanina, tirosina y lisina también fueron identificados.
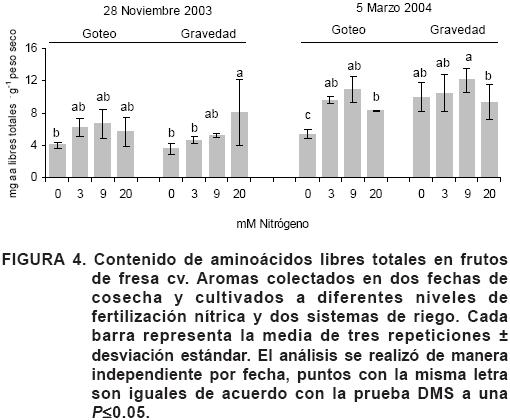
En la Figura 4 se muestra el contenido de aminoácidos libres totales identificados en los frutos analizados. En el primer pico de producción (28 de noviembre) la mayor acumulación de aminoácidos libres totales en el fruto fue significativa en riego por gravedad en el nivel de fertilización de 20 mM (8.06 mg·g–1 p.s.) respecto a 0 y 3 mM, pero no así para el sistema de riego por goteo, que sólo tendió a ser en el nivel de fertilización de 9 mM (6.68 mg·g–1 p.s.). En el segundo pico de producción, el contenido de aminoácidos libres aumentó al incrementar el nivel de fertilización, en donde en el sistema de riego por goteo la mayor acumulación fue en 9 mM respecto a 0 mM (10.93 y 5.40 mg·g–1 p.s., respectivamente) y en gravedad fue también en 9 mM pero con respecto a 20 mM (12.09 y 9.40 mg·g–1 p.s., respectivamente).
En la Figura 5 se muestra el contenido de aminoácidos esenciales libres de los frutos cosechados. En la primera fecha y en ambos sistemas de riego se observó que la concentración aumentó al incrementar el nivel de fertilización y la mayor acumulación significativa fue en el nivel de 20 mM respecto a 0 mM (3.93 mg·g–1 p.s. en goteo y 4.11 mg·g–1 p.s. en gravedad). En el caso de la segunda fecha de cosecha y en el sistema de riego por goteo, la acumulación de aminoácidos esenciales fue significativa a partir de 3 mM hasta alcanzar un máximo en el de 9 mM (5.10 mg·g–1 p.s.), mientras que el riego por gravedad fue en los niveles 0 y 3 mM (4.03 y 4.87 mg·g–1 p.s., respectivamente).

De los aminoácidos esenciales identificados en los frutos analizados, se observó que la fenilalanina en ambas fechas de cosecha se acumuló en el nivel de 9 mM en riego por goteo, mientras que en riego por gravedad fue en 20 mM. En lo que respecta a la acumulación de treonina en la primera fecha de cosecha, en ambos sistemas de riego sólo tendió a aumentar en 20 mM; no así para la segunda fecha donde en riego por goteo la mayor acumulación fue significativa en 9 mM respecto de 0 y 3 mM y en gravedad fue en 3 mM respecto a los niveles de 0, 9 y 20 mM. Otro aminoácido esencial identificado fue la lisina, cuya concentración evaluada en los frutos cosechados el 28 de noviembre y en ambos sistemas de riego fue significativamente mayor en el nivel de 3 mM respecto a 0 y 9 mM; mientras que en los frutos pertenecientes al 5 de marzo en riego por goteo la mayor acumulación fue en 9 mM y en gravedad fue en 20 mM (Cuadro 3).

Los aminoácidos que se sabe que son precursores del aroma de la fresa, como la alanina, isoleucina, leucina y valina se pudo observar que la concentración de la alanina en los frutos del 28 de noviembre y en ambos sistemas de riego fue significativamente mayor en 9 mM; mientras que en los frutos del 5 de marzo en riego por goteo que fue en 20 mM y en gravedad fue en 9 mM. La concentración de isoleucina en los frutos pertenecientes a la cosecha del 28 de noviembre, se observó que la mayor acumulación tendió a ser en 20 y 3 mM para riego por goteo y gravedad, respectivamente; mientras que en la segunda fecha evaluada la acumulación fue significativa en 9 y 3 mM para riego por goteo y gravedad, respectivamente. En el caso de la leucina se observó que en la primera fecha la concentración mayor fue en los niveles 20 mM para goteo y 9 mM para gravedad; no siendo así para la segunda fecha evaluada donde en ambos riegos la acumulación significativa fue en 0 mM. En la valina se observó que la acumulación en el 28 de noviembre, tendió a ser mayor en 0 mM para riego por goteo y en 20 mM para riego por gravedad; mientras que el 5 de marzo se acumuló significativamente en 9 y 3 mM para riego por goteo y gravedad, respectivamente (Cuadro 4).

En el Cuadro 5 se muestran los aminoácidos no esenciales identificados en los frutos cosechados en las dos fechas seleccionadas. Los resultados obtenidos de glu/gln indicaron que la concentración de estos compuestos en los frutos cosechados en el primer pico de producción fue similar para ambos sistemas de riego; siendo que en el sistema por goteo el nivel de fertilización nítrica de 3 mM produjo una concentración significativamente más alta que la de 20 mM. En la fecha del segundo pico de producción en el sistema de riego por gravedad el nivel de fertilización de 9 mM produjo una concentración de glu/gln significativamente más alta que la del resto de los niveles de fertilización. Esta misma tendencia se observó en el riego por goteo. En lo que respecta al contenido de asp/asn, los frutos de fresa cosechados en la fecha del 28 de noviembre en riego por gravedad se observó que tendió a ser mayor en el nivel de 20 mM, con respecto a los demás niveles de fertilización. En el caso del riego por goteo, no hubo diferencias significativas en el contenido de asp/asn, solo se observó la tendencia a acumularse en el nivel de 9 mM. Los frutos colectados en la fecha del 5 de marzo bajo el sistema de riego por gravedad se obtuvo el máximo contenido de asp/asn en las plantas no fertilizadas, que fue significativamente mayor al encontrado en el nivel de fertilización de 20 mM, mientras que en riego por goteo se acumuló significativamente en 9 y 20 mM con respecto a 0 mM. De manera general en la primer fecha la concentración de los aminoácidos glicina y serina tendió a incrementarse significativamente en el nivel de 9 mM en riego por gravedad mientras que en goteo fue en 20 y 3 mM, respectivamente; para la segunda fecha de cosecha se observó que la glicina se acumuló en 20 mM en goteo y en 9 mM en gravedad; mientras que la mayor acumulación de serina fue en 0 mM en ambos sistemas de riego. Para tirosina, se observó que el 28 de noviembre se acumuló significativamente en 9 mM y 3 mM, en goteo y gravedad respectivamente; mientras que el 5 de marzo en riego por goteo tendió a acumularse en 3 y 9 mM pero para gravedad se acumuló significativamente en 0 y 20 mM respecto al nivel de 9 mM.
Ácidos orgánicos
En el Cuadro 6 se muestra el contenido de los ácidos ascórbico y cítrico en los frutos cosechados en ambas fechas de muestreo. Como se puede observar, el contenido del ácido ascórbico fue el doble y en algunos casos superior al contenido del ácido cítrico. De forma general, el contenido de ácido ascórbico tendió a disminuir al aumentar la dosis de fertilización aplicada. Este comportamiento también se observó en el caso del ácido cítrico cuantificado en los frutos del 28 de noviembre, pero no así para el 5 de marzo, donde se observó que la mayor acumulación de este ácido fue en 9 y 20 mM para riego por goteo y gravedad, respectivamente.

DISCUSIÓN
Los estudios que se han realizado en nutrición vegetal están enfocados hacia determinar los niveles de N requeridos por la planta para incrementar su rendimiento; sin ser prioritaria la calidad organoléptica del fruto, que se sabe está fuertemente influenciada por las prácticas de cultivo (Prange y DeEll, 1997).
En este trabajo se pudo observar que la acumulación de los carbohidratos, aminoácidos y ácidos orgánicos fue afectada, tanto por la aplicación de diferentes niveles de nitrato, como por la forma de riego. Esto sugiere que la calidad sensorial y nutrimental del fruto ha sido afectada también.
En el caso específico del análisis de carbohidratos en los frutos cosechados en dos fechas representativas, sometidos a diferentes niveles de fertilización nítrica y a dos sistemas de riego indicó que los compuestos más abundantes fueron fructosa y glucosa, lo que coincide con lo reportado por Pérez et al. (1997). Otro compuesto importante reportado por estos mismos autores en fresa es la sacarosa, que en este estudio no se identificó como uno de los azúcares más abundantes, probablemente porque la técnica de identificación por medio del detector de ionización de flama (FID) no es la adecuada para efectuar su análisis y cuantificación.
Los análisis mostraron que las concentraciones de carbohidratos libres totales, así como el contenido de fructosa y glucosa aumentaron conforme se incrementó el nivel de fertilización hasta alcanzar un máximo en los niveles 3 o 9 mM, para posteriormente disminuir en dosis de 20 mM de nitrato (Figuras 1, 2 y 3), efecto que fue más marcado en la fecha del 28 de noviembre. En la fecha del 5 de marzo sólo se presentaron tendencias en la acumulación de estos carbohidratos y puede deberse a que la planta concentra azúcares en este órgano para evitar la pérdida de agua por la alta demanda evaporativa que caracteriza a esta época del año en comparación con el mes de noviembre. Así, que los frutos de plantas cultivadas con deficiencia de N al estar más expuestos al sol, por el poco crecimiento vegetativo, tendieron a acumular carbohidratos y no se aprecian las diferencias entre niveles de fertilización.
En lo que respecta a las pentosas (ribosa, arabinosa y xilosa), se observó una tendencia no muy clara del nivel de fertilización sobre la acumulación de estos azúcares (Cuadro 1), lo que sugiere que la fertilización nitrogenada puede influenciar el metabolismo de las pentosas en las plantas de fresa, el cual es crucial para la biosíntesis de L–arabanos y D–xilanos, quienes estarían participando en la formación de rizomas, hojas, raíces y frutos (Luckner, 1990). En otras hexosas, como la manosa y galactosa, de manera general se volvió a encontrar la misma tendencia a acumularse principalmente en los niveles de 3 y 9 mM (Cuadro 2), al igual que el myo–inositol, que según Loewus y Loewus (1980) participa en la biosíntesis del ácido fítico, el cual es considerado como una fuente importante de fosfato en las plantas.
Con estos resultados se llegó a ver que a niveles altos de N no se obtiene una mayor concentración de azúcares en el fruto, sino una retardación en la maduración, pérdida de firmeza y disminución del tamaño del fruto (May y Pritts, 1990).
Otro pilar fundamental en el sabor y aroma de la fresa son los aminoácidos, que con sus estructuras hidrocarbonadas diversas, son precursores de metabolitos secundarios como es el caso de los compuestos volátiles que denotan el aroma en las frutas.
Los resultados mostraron que en el segundo pico de producción se produjo una mejor calidad de fresa en términos de aminoácidos libres totales (Figura 4), esto podría deberse a que en el segundo pico de producción la planta ha terminado ya su crecimiento vegetativo y concentra los aminoácidos disponibles en el fruto.
Darnell y Stutte (2001) observaron que al aplicar dosis crecientes de nitrato en plantas no se incrementó el contenido de NO3– en el fruto, sugiriendo que el fruto tiene un potencial de almacenamiento que al ser saturado existe una relocalización continua de N en función de la concentración de NO3– externo. El análisis del contenido de aminoácidos libres totales, el glu/gln y asp/asn mostró que se acumularon a partir de 3 mM (Figura 4 y Cuadro 5), lo cual era de esperarse, ya que estos compuestos distribuyen el N a toda la planta de acuerdo a la demanda metabólica y de la concentración de NO3– externo. También es de notarse que el incremento de la dosis de fertilización no aumenta de manera significativa la acumulación de los aminoácidos de transporte de N y que el potencial de almacén de estos compuestos es saturado en los niveles de 3 y 9 mM.
El valor nutritivo de los frutos analizados, en términos de acumulación de aminoácidos esenciales, se afectó por la fertilización nitrogenada y el sistema de riego aplicado, siendo en el nivel 9 mM de N en la segunda fecha analizada y en riego por goteo el que favoreció dicha acumulación (Figura 5).
Los aminoácidos alanina, isoleucina, leucina y valina son considerados como precursores de compuestos volátiles en fresa, ya que se ha observado que el contenido de éstos varía significativamente durante la maduración del fruto, por lo que se sugiere que pueden ser los responsables del alto contenido de ésteres en el perfil aromático de dicho fruto (Pérez et al., 1992). En nuestros resultados se observó que es precisamente en isoleucina y valina donde se notó una mayor acumulación en los niveles de 3 y 9 mM de nitrato (Cuadro 4), lo que muestra que la fertilización nitrogenada influye sobre la síntesis de los compuestos precursores del aroma en el fruto.
El fruto de fresa es también órgano de almacenamiento de compuestos carbonados, como los ácidos orgánicos y por tanto refleja el estado metabólico de la planta, el cual es altamente influenciado por la nutrición y disponibilidad de agua en el suelo.
El ácido ascórbico es una g–lactona que puede ser sintetizado a partir de glucosa y galactosa, por lo que su contenido está en función de la concentración de estos azúcares en el fruto. De manera general, los azúcares presentaron la tendencia a incrementarse en los niveles 3 y 9 mM en ambas fechas de muestreo, por ambos sistemas de riego (Cuadro 2 y Figura 3), pero la acumulación de ácido ascórbico tendió a disminuir al aumentar el nivel de fertilización (Cuadro 6), esto puede deberse a que el ácido ascórbico puede estar utilizándose como antioxidante para contrarrestar alguna explosión oxidativa que pudiera estar desencadenando las altas dosis de fertilización nitrogenada. En el caso del ácido cítrico, se observó que la mayor acumulación fue en los niveles de 9 y 20 mM de N en la segunda fecha de cosecha y en los sistemas de riego por goteo y gravedad, respectivamente (Cuadro 6); esto podría deberse a un ajuste metabólico por parte de la planta, debido a que durante la asimilación de nitrato la síntesis de carbohidratos disminuye y más carbono entra al metabolismo de los ácidos orgánicos, el cual tiene distintas funciones dentro de la asimilación de nitrato; una de ellas para sintetizar malato, que actúa como contraanión para evitar la alcalinización y la otra es proveer 2–oxoglutarato que es el aceptor primario para amonio (Stitt et al., 2002). Lo anterior sugiere que la acidez de la fruta está influenciada también por las dosis de N aplicadas y el sistema de riego utilizado.
Basándose en los resultados obtenidos, se propone que el metabolismo de las plantas cultivadas, dado por la interacción de la asimilación del carbono y del nitrógeno, está en equilibrio en los niveles de 3 y 9 mM, dando como resultado una planta vigorosa que produjo frutos de buena calidad gustativa y nutrimental reflejado por la acumulación de los compuestos evaluados.
En el caso de las plantas no fertilizadas, se observó de manera general una concentración menor de los compuestos evaluados, respecto a los niveles 3 y 9 mM, que puede deberse a un desequilibrio metabólico originado por la baja asimilación de nitrogenados en la planta (dada por la baja concentración de nitrato disponible). De manera que, si la disponibilidad de los asimilados de N es menor a lo que demanda la planta para sostener el proceso de fotosíntesis, la composición de los tejidos y el crecimiento de la planta son dispares (Ulrich et al., 1980; Yoshida et al., 2002; Nestby et al., 2005), trayendo como consecuencia frutos de una calidad gustativa y nutrimental no satisfactorias.
Por otro lado, en las plantas fertilizadas con el nivel de 20 mM se observó una disminución en la concentración de la mayoría de los compuestos de calidad gustativa y nutrimental evaluados, respecto a los niveles de 3 y 9 mM, siendo el caso de la fructosa, el ácido cítrico y ascórbico. Esta respuesta puede deberse a un desajuste metabólico en la planta, debido a que la demanda de esqueletos carbonados aumenta a consecuencia de la alta tasa de asimilación de nitrato (Stitt et al., 2002). De esta manera, la disponibilidad de fotosintatos disminuye y menos compuestos carbonados son distribuidos a los órganos de almacén, lo que dió como resultado una planta con un desarrollo vegetativo excesivo que produjo frutos con una baja acumulación de los compuestos y por tanto, con una baja calidad sensorial y nutrimental.
Respecto a los sistemas de riego evaluados, se observó que en riego por goteo la acumulación de los compuestos evaluados fue más predecible y hubo menor variabilidad de los resultados dentro de cada tratamiento. En el caso del sistema de riego por gravedad no fue así, ya que la acumulación de metabolitos fue inconsistente y se encontró más variabilidad dentro de los tratamientos evaluados. Esto sugiere que la disponibilidad del nitrato se afecta cuando se utiliza el riego por gravedad y provoca que se acentúe el efecto que tienen otros factores, como los ambientales, sobre el metabolismo y el rendimiento de la fresa. Un ejemplo de lo anterior, se observó en los frutos cosechados el 5 de marzo, donde hubo una mayor acumulación de asp/asn en los frutos de plantas no fertilizadas que en los frutos de plantas fertilizadas cuando el sistema de riego fue por gravedad; pero en riego por goteo no fue así, pues la acumulación de este aminoácido fue aumentando al incrementar la concentración de nitrato externo (Cuadro 5); lo que indica que de alguna forma el sistema de riego está influyendo en la disponibilidad de N en el suelo y por tanto, afecta la acumulación de los compuestos en los órganos de la planta, como el fruto.
También se observó que en el riego por goteo con un nivel de fertilización de 3 mM nitrato se logró la acumulación de la mayoría de los compuestos evaluados; mientras que en riego por gravedad se necesitó el nivel de 9 mM de nitrato para lograr una acumulación similar de los compuestos. Así que al aplicar el riego en exceso para contrarrestar la sensibilidad de la fresa al estrés hídrico (Mass, 1987) disminuye la disponibilidad del fertilizante en el suelo por lixiviación y por pérdidas gaseosas de nitrógeno, afectando entonces la calidad sensorial de la fresa. Por lo anterior, es importante considerar que con un sistema de riego por goteo se tiene un menor impacto ambiental y económico (Monroy et al., 2002).
CONCLUSIONES
Los resultados anteriormente descritos permitieron concluir que los frutos cosechados en campo en los niveles de fertilización comprendidos por las dosis de 3 mM en riego por goteo y 9 mM de nitrato en riego por gravedad, favorecieron la acumulación de los compuestos relacionados a calidad sensorial y nutrimental en fresa. Esta observación es importante, ya que mediante la optimización y el uso eficiente de fertilizantes y del agua se pueden obtener frutos de buena calidad organoléptica y nutrimental, además de que disminuye el impacto ambiental y los costos económicos en la producción de fresa. Así que una mayor fertilización nitrogenada y la aplicación excesiva de agua no se justifican en términos de contenido de estos compuestos, pudiéndose disminuir el consumo de estos insumos que además contribuirían a disminuir los costos de producción de fruto de buena calidad organoléptica.
LITERATURA CITADA
CORDENUNSI, B.; OLIVEIRA, J.; GENOVESE, M.; LAJOLO, F. 2002. Influence of cultivar on quality parameters and chemical composition of strawberry fruits grown in Brazil. Journal of Agricultural and Food Chemistry. 50: 2581–2586. [ Links ]
DARNELL, R.; STUTTE, G. 2001. Nitrate concentration effects on NO3–N uptake and reduction, growth, and fruit yield in strawberry. Journal of the American Society for Horticultural Science 125(5): 560–563. [ Links ]
HANCOCK J., F. 1999. Strawberry. CABI Publishing. London, United Kindom. 237 p. [ Links ]
LOEWUS, F.; LOEWUS, M. 1980. Myo–inositol: biosynthesis and metabolism. pp. 43–76. In: The Biochemistry of Plants A Comprehensive Treatise. Stumpf, P. & Conn, E. (Eds.). Academic Press: New York. [ Links ]
LUCKNER, M. 1990. Secondary metabolism in microorganisms, plants, and animals. Springer–Verlag: Berlin. pp. 102. [ Links ]
MACÍAS–RODRÍGUEZ, L.; QUERO E. LÓPEZ, M. G. 2002. Carbohydrate Differences in Strawberry Crowns and Fruit (Fragaria x ananassa) during Plant Development. Journal of Agricultural and Food Chemistry. 50, 3317–3321. [ Links ]
MASS, J. 1987. Compendium of strawberry diseases. APS Press, St–Paul, Minn. pp. 137. [ Links ]
MAY, G; PRITTS, M. 1990. Strawberry Nutrition. Advances in Strawberry Production, 9, 10–23. [ Links ]
MONROY, J.; VERA–NUÑEZ, J. A.; CARRERA, M. A.; GRAGEDA–CABRERA, O. A; PEÑA–CABRIALES, J. J. 2002. Absorción de nitrógeno (15N) y productividad del agua por el cultivo de fresa (Fragaria x ananassa) en "El Bajío", México. Terra, 20, 65–69. [ Links ]
NESTBY, R.; LIETEN, F.; PIVOT, D.; RAYNAL LACROIX, C.; TAGLIAVINI, M. 2005. Influence of mineral nutrients on strawberry fruit quality and their accumulation in plant organs: a review. International Journal of Fruit Sciences, 5(1): 141–158. [ Links ]
PÉREZ, A.; OLÍAS, R.; ESPADA, J.; OLÍAS, J; SANZ, C. 1997. Rapid determination of sugars, nonvolatile acids, and ascorbic acid in strawberry and other fruits. Journal of Agricultural and Food Chemistry. 45, 3545–3549. [ Links ]
PÉREZ, A.; RÍOS, J.; SANZ, C.; OLIAS, J. 1992. Aroma components and free amino acids in strawberry variety chandler durin ripening. Journal of Agricultural and Food Chemistry. 40, 2232–2235. [ Links ]
PRANGE, R.; DeELL, J. 1997. Preharvest factors affecting postharvest quality of berry crops– HortScience. 32(5): 824–829. [ Links ]
SAS INSTITUTE. 1988. SAS/STAT user's guide. Release 6.03 ed. SAS Inst. Cary, NC. [ Links ]
SHAMAILA, M.; BAUMANN, T.; EATON, G.; POWRIE, W.; SKURA, B. 1992. Quality attributes of strawberry cultivars grown in British Columbia. Journal of Food Science. 57, 696–699. [ Links ]
STITT, M.; MÜLLER, C.; MATT, P.; GIBON, Y.; CARILLO, P.; MORCUENDE, R.; SCHEIBLE, W.; KRAPP, A. 2002. Steps towards an integrated view of nitrogen metabolism. Journal of Experimental Botany. 53, 959–970. [ Links ]
ULRICH, A.; MOSTAFA, M.; ALLEN, W. 1980. Strawberry Deficiency Symptoms: a Visual and Plant Analysis Guide to Fertilization, University of California Agricultural Science Publication. 4098. [ Links ]
YOSHIDA, Y.; GOTO, T.; IRÍA, M.; MASUDA, M. 2002. Anthocyanin accumulation in strawberry fruits as affected by nitrogen nutrition. Acta Horticulturae, 567: 357–360. [ Links ]
ZABETAKIS, I.; HOLDEN, M. 1997. Strawberry flavour: analysis and biosynthesis. Journal of the Science and Food Agriculture. 74, 421–434. [ Links ]

