










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.26 no.2 Ciudad de México ago. 2009
Taxonomic revision and evolutionary classification of the biserial Cretaceous planktic foraminiferal genus Laeviheterohelix Nederbragt, 1991
Revisión taxonómica y clasificación evolutiva del género de foraminífero planctónico biserial del Cretácico Laeviheterohelix Nederbragt, 1991
Marius Dan Georgescu
Department of Geoscience, University of Calgary, 2500 University Drive N.W., Calgary, Alberta, T2N 1N4, Canada. dgeorge@ucalgary.ca
Manuscript recieved: March 3, 2008
Corrected manuscript received: February 20, 2009
Manuscript accepted: March 9, 2009
ABSTRACT
Taxonomic re–evaluation of the Upper Cretaceous serial planktic foraminiferal genus Laeviheterohelix Nederbragt, 1991 revealed that the genus is polyphyletic. Three genera are recognized, two new, Steineckia n. gen. and Fleisherites n. gen., and one emended, namely Laeviheterohelix. Steineckia n. gen. is the oldest serial planktic foraminifer with pore mounds and it appears restricted to the upper Turonian sediments of the South Atlantic (Falkland Plateau). It includes only the type species, Steineckia steinecki n. sp. The genus Laeviheterohelix is revised and according to the emendation proposed here consists only of species from the Upper Coniacian–Campanian stratigraphic interval: L. pulchra (Brotzen, 1936) – emended and L. reniformis (Marie, 1941) – emended. The species with smooth tests and occasionally with incipient costae of the Maastrichtian are included within Fleisherites n. gen. Its type species is Fleisherites glabrans (Cushman, 1938). This newly proposed taxonomic framework is based on detailed, scanning electron microscope–based observations on the test ornamentation and ultrastructure. The taxonomic significance of the presence/absence of the pore mounds, periapertural pustulose area and pore mound size is of paramount importance in documenting the iterative evolution pattern in the serial planktic foraminifera with smooth test wall or ornamented with pore mounds.
Key words: planktic foraminifera, taxonomy, new genera, new species, Upper Cretaceous.
RESUMEN
La re–evaluación taxonómica de la serie foraminífera planctónica del género Laeviheterohelix Nederbragt, 1991 del Cretáceo Tardío, demuestra que el género es polifiletico. Tres géneros fueron reconocidos, dos son nuevos: Steineckia n. gen. y Fleisherites n. gen., y uno renombrado: Laeviheterohelix. Steineckia n. gen., el cual es la serie foraminífera planctónica más antigua con montículos porosos y aparece restringida a sedimentos del Turoniano superior del Atlántico Sur (Plateau de las Malvinas). Este incluye solamente la especie tipo, Steineckia steinecki n. sp. El género Laeviheterohelix es revisado y de acuerdo al renombramiento propuesto en este trabajo, consiste únicamente de especies del intervalo estratigráfico del Coniaciano Tardío–Campaniano: L. pulchra (Brotzen, 1936) – renombrado y L. reniformis (Marie, 1941) – renombrado. Las especies con conchas suaves y ocasionalmente con costillas incipientes del Maastrichtiano son incluidas dentro del género Fleisherites n. gen. Su especie tipo es Fleisherites glabrans (Cushman, 1938). Esta nueva propuesta taxonómica se basa en observaciones detalladas de la ornamentación de las conchas y la ultraestructura utilizando un microscopio electrónico de barrido. El significado taxonómico de la presencia/ausencia de montículos porosos, área periapertural postulosa y el tamaño de los monticulos es de suprema importancia al documentar el patrón iterativo de evolución en la serie foraminífera planctónica con conchas suaves o con ornamentación de montículos porosos.
Palabras clave: foraminíferos planctónicos, taxonomía, género nuevo, especie nueva, Cretácico Superior.
INTRODUCTION
Biserial planktic foraminifera commenced their evolution in the upper Albian. The earlier representatives of the group, namely those of the upper Albian–lower Santonian stratigraphical interval generally present small tests. Larger tests are achieved especially with the development of adult multiserial chamber stages (upper Santonian–Maastrichtian). Most of the taxa included among the heterohelicids have a costate test surface (e.g., Pseudotextularia, Ventilabrella, Planoglobulina, Racemiguembelina). There are relatively fewer biserial taxa with smooth chamber surface or with pores situated in the center of a pore mound (e.g., Laeviheterohelix) and this kind of test ornamentation occurs in Turonian–Campanian.
Detailed observations on the heterohelicid tests are difficult to make with the classical optical microscope due to the small size of the various structures (e.g., periapertural structures, test ornamentation, pore size, etc). This impediment can be overcome by making observations with the aid of a large number of scanning electron microscope photographs. High detail SEM–based observations on tests, which were traditionally assigned to various species of Laeviheterohelix, resulted in a better understanding of the test morphology and variability of certain structures whose taxonomic significance was neglected in the past. The size of the pore mounds, presence/absence of a periapertural pustulose area and the presence/absence of vestigial costae are among these. The last mentioned is a feature that can be used to document the origins of certain smooth species or with pore mounds among the costate ones.
These observations demonstrate that the species with smooth tests or ornamented with pore mounds do not form a single plexus as considered since the definition of Laeviheterohelix (Nederbragt, 1991). Three different origins can be inferred for the genus Laeviheterohelix based on detailed test ultrastructure and ornamentation. Accordingly, three genera are now recognized: Steineckia n. gen. (upper Turonian), Laeviheterohelix – emended (upper Coniacian–Campanian) and Fleisherites n. gen. (Maastrichtian).
BRIEF HISTORY OF CONCEPTS IN HETEROHELICID CLASSIFICATION
Test ultrastructure and ornamentation was not considered a major morphological feature in the biserial planktic foraminiferal taxonomy at the genus level until the description of Laeviheterohelix in the last major taxonomic revision of the group (Nederbragt, 1991). This genus was originally described based on specimens with a smooth chamber surface and pore mounds in some of its species. Laeviheterohelix pulchra (Brotzen, 1936) was selected its type species. The description of Laeviheterohelix formalized the difference between the taxa with smooth test or with pore mounds and the rest of the biserial or biserial–multiserial Cretaceous planktic foraminifera, which were considered costate. This point of view received wide recognition in the subsequent studies on the Cretaceous representatives of the serial planktics (Huber, 1991; Premoli Silva and Sliter, 1995; Zapeda, 1998; Petrizzo, 2001; Abramovich et al., 2003; Georgescu, 2006).
Although formalization at the genus level is relatively recent, another heterohelicid species with a smooth test wall was described from the upper Albian–Cenomanian of Texas by Tappan (1940), namely Guembelina washitensis. The genus Guembelina Egger, 1902 is a junior synonym of Heterohelix Ehrenberg, 1843 as demonstrated by Hofker (1957), Montanaro Gallitelli (1957) and Loeblich and Tappan (1961). However, Heterohelix washitensis was subsequently considered a junior synonym of Heterohelix moremani (Cushman, 1938) by Pessagno (1967) and Nederbragt (1991).
The first observations on the development of the costate ornamentation were made by Thomas (1927) and Thomas and Rice (1927) who noted that the earlier heterohelicids from the Cenomanian–Turonian) are smooth and the later ones gradually acquire costate ornamentation. This aspect of the heterohelicid evolution was reconsidered by Brown (1969). In his excellent review of the group, this author concluded that it is possible to confer certain taxonomic significance to the test ornamentation. However, the general small size of the earlier heterohelicid taxa can impede the systematic use of this feature for taxonomical purposes.
A major step forward in the study of the Cretaceous heterohelicids happened with the introduction of scanning electron microscopy in micropaleontological practice in the early 70s. The group was thoroughly reviewed in the following two decades by Masters (1977), Weiss (1983) and Nederbragt (1989, 1991). Moreover, it received a thorough additional review at the genus level by Loeblich and Tappan (1987) in 'Foraminiferal Genera and Their Classification'. More recent and detailed studies on Cretaceous serial planktic foraminifera demonstrated the importance of test ornamentation and ultrastructure in the taxonomy of the group (Georgescu, 2007a, 2007b; Georgescu and Abramovich 2008a, 2008b; Georgescu and Huber, 2008; Georgescu et al., 2009).
MATERIAL AND METHODS
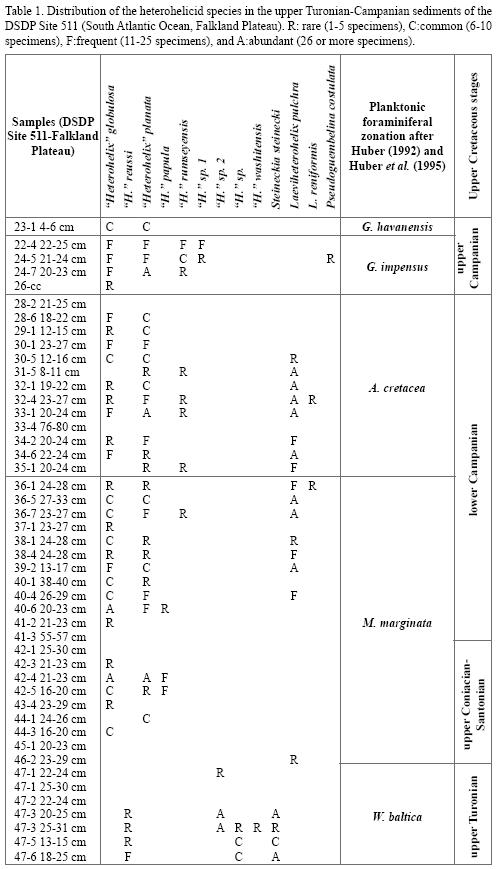
Most of the material analyzed in this study was collected from 15 DSDP/ODP wells: DSDP Hole 111A (Orphan Basin, North Atlantic), DSDP Site 305 (Shatsky Rise, Equatorial Pacific), DSDP Site 356 (São Paolo Plateau, western South Atlantic), DSDP Site 357 (Rio Grande Rise, western South Atlantic), DSDP Site 384 (Grand Banks, North Atlantic), DSDP Site 463 (Mid–Pacific Mountains, Equatorial Pacific), DSDP Site 511 (Falkland Plateau, South Atlantic Ocean), ODP Leg 174AX (New Jersey coastal plain), ODP Holes 689B and 690C (Maud Rise, South Atlantic), ODP Holes 761B and 762C (Exmouth Plateau, Indian Ocean) and ODP Holes 1049B, 1050C and 1052E (western North Atlantic, Blake Plateau) (Figure 1). The most informative section proved that from DSDP Site 511 and species distribution is given in Table 1. The samples analyzed from the New Jersey coastal plain subsurface are deposited in the Willi Karl Braun Micropaleontological Collection (WKB) at the University of Calgary. All the others are deposited in the Ocean Micropaleontology Collection at the National Museum of Natural History (NMNH), Smithsonian Institution, Washington, D.C. Samples of provenance for the various specimens are labeled according to the DSDP/ODP standards: leg number–site or hole number–core number–core section and the interval from which the sample was collected (given in centimeters). The only exception is represented by the specimens from ODP Leg 174AX at the Bass River Site (New Jersey coastal plain) in which, according to the original labeling, the leg number is followed by the depth in metres.
Type specimens were analyzed from the Cushman Collection, NMNH. The holotype of Guembelina glabrans Cushman, 1938 was recently figured in the Taxonomic Dictionary of Mesozoic Planktic Foraminifera (Huber, 2006) and is re–illustrated herein. A paratype of Guembelina pulchra Brotzen, 1936 deposited in the Cushman Collection (NMNH) is figured for the first time (Figure 8.2), becoming thus the third refigured paratype of the species, after the two illustrated by Weiss (1983). The very rich and diverse material from the Gulf of Mexico Santonian of the Van Morkhoven Collection (NMNH) was also investigated.
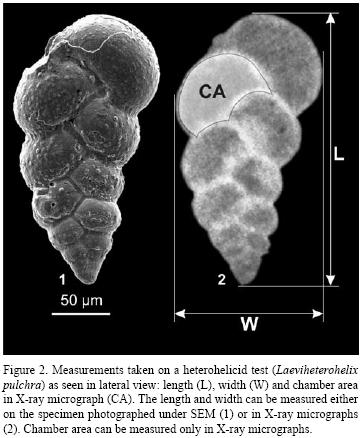
The specimens were studied using both the binocular optical microscope and the scanning electron microscope. Approximately 450 SEM micrographs were taken in the course of this study. Internal structure of the specimens of Laeviheterohelix and phylogenetically related or morphologically similar taxa was studied using X–ray micrographs (Figure 2). The X–ray micrographs of 15 specimens were taken on SO–253 High Speed Holographic film (exposure time 2.5 minutes at 20 kilovolts); they were developed with D–19 developer and fixed in F–5 fixing bath. The X–ray micrographs were then used for biometrical analysis and constructing the ontogenetic trajectories based on chamber increase in size for the specimens. Measurements were made using the Optimas, version 6.5 software.
All these data converged in extremely detailed observations and descriptions of the studied species and genera. Emphasis was put on understanding species variability and evolutionary relationships between the various species. As a result, all the species could be defined using the paleontological species concept given by Georgescu and Huber (2007, p. 158): "A well–documented paleontological species is the basic unit with taxonomic significance in the fossil record, and has the following characteristics: (i) it is monophyletic; (ii) it has a distinct range of morphological variability, showing relative stability over a definable period of time and presenting relatively discrete evolutionary changes; (iii) it is a morphologically heterogeneous and discontinuous entity, consisting of one or (mostly) more morphological and/or paleoecological varieties; (iv) it has its own developmental history traceable in space and time; and (v) its existence and integrity can be tested not only by comparative morphological distinctiveness, but also by its response to paleoenvironmental and geological factors (e.g., paleoclimatic changes, sea–level fluctuations, etc.), as inferred from paleontology and related geological disciplines."
REVISED MORPHOLOGICAL FEATURES
The use of the scanning electron microscope (SEM) and environmental scanning electron microscope (ESEM) in acquiring morphological data on Laeviheterohelix and similar genera revealed the necessity to re–evaluate the taxonomic role of certain test features. The morphological features are presented in detail here: test ornamentation, periapertural pustulose area and size of the pore mound. Notably, one of them is newly described, namely the periapertural pustulose area.
The dominant test ornamentation among the heterohelicids is the costate one. Apparently it is the result of a longer evolutionary process as the earliest tests of the upper Albian and most of the Cenomanian are smooth (Thomas, 1927; Thomas and Rice, 1927; Brown, 1969). Test ornamentation was considered relatively uniform for a longer period of time, all the heterohelicid taxa being described as either smooth, or costate, or with pore mounds (Cushman, 1938; Pessagno, 1967; Masters, 1977; Nederbragt, 1991). It was demonstrated, only recently, that the test ornamentation changes within a lineage, namely in the case of the genus Braunella (Georgescu, 2007a).
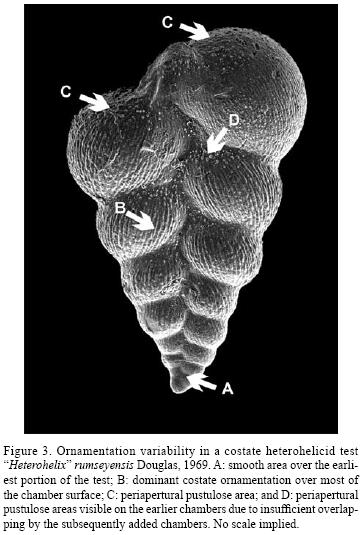
Extensive observations on the tests of the whole Cretaceous heterohelicid group show that the test ornamentation in not uniformly developed over the test surface and certain areas with distinct ornamentation patterns can be defined. These areas can be best seen in the case of a costate test. Costate ornamentation is the dominant one and is developed over most or all of the chambers of the test. A smooth area can be present over the earliest portion of the test. A distinct pustulose area can be recognized at the top of each chamber in certain species. This feature is named here "periapertural pustulose area". Parts of the periapertural pustulose areas can remain visible along the sutures through insufficient overlapping of successive chambers (Figure 3). It is noteworthy that the periapertural pustulose area proved highly significant for taxonomical purposes and in evolutionary reconstructions (Figure 4).
The presence of the pore mounds on the surface of some Laeviheterohelix species was considered a major taxonomic criterion and was used by Nederbragt (1991) in the definition of the genus. Smaller pore mounds (D = 1.7–2.8 µm) are characteristic for a group of upper Coniacian–Campanian species, which includes the biserial species Laeviheterohelix pulchra (Brotzen, 1936). Large–sized pore mounds occur in the upper Turonian species Steineckia steinecki n. sp. (D = 4.0–6.4 µm) and the Campanian L. reniformis, the latter being a descendant from L. pulchra (D = 3.2–6.9 µm) (Figure 5).
Pore size terminology is refined when compared to that adopted by the Mesozoic Planktonic Foraminifera Working Group: nannoperforate (Ø < 0.5 µm), microperforate (Ø = 0.5–1.0 µm), finely perforate ((Ø = 1.0–2.5 µm) and macroperforate (Ø> 2.5 µm). The biserial planktic foraminifera with smooth test surface or ornamented with pore mounds often present nannoperforate to microperforate test wall. Finely perforate tests are known only in Laeviheterohelix reniformis, a species with incipient chamber proliferation in the adult portion of the test.
SYSTEMATIC PALEONTOLOGY
Suprageneric classification is after Loeblich and Tappan (1987). Species concept is after Georgescu and Huber (2007).
Order Foraminiferida Eichwald, 1830
Suborder Rotaliina Delage and Hérouard, 1896
Superfamily Heterohelicacea Cushman, 1927
Family Heterohelicidae Cushman, 1927
Genus Laeviheterohelix Nederbragt, 1991 – emended
Type species. Guembelina pulchra Brotzen, 1936.
Laeviheterohelix Nederbragt, 1991, p. 350.
Emended diagnosis. Test wall is nannoperforate to microperforate, with small sized pore mounds and periapertural pustulose area.
Description. Test is biserial throughout or with incipient multiserial stage. Earlier chambers are globular to subglobular, the last–formed ones reniform or, more rarely, tubular. Periphery is broadly rounded, without peripheral structures. Aperture is situated at the base of the last–formed chamber and is bordered by two well–developed, symmetrical flanges. Chamber surface bears small sized pore mounds in the primitive biserial species and with larger diameter in the species with incipient multiserial stage and, more rarely, vestiges of the ancestral ornamentation. Well–developed periapertural pustulose area at the upper part of the chambers. Test wall is calcitic, hyaline, nannoperforate to finely perforate.
Species included.
Guembelina pulchra Brotzen, 1936, p. 121, pl. 9, fig. 3 (only).
Ventilabrella reniformis Marie, 1941, p. 264, pl. 28, fig. 277.
Remarks. Laeviheterohelix is redefined now in a narrower sense when compared to the original diagnosis (Nederbragt, 1991). It includes only taxa with small sized pore mounds and apertural pustulose area at the top of the chambers. Vestigial costae are observed only on the last–formed chambers in Laeviheterohelix pulchra (approximately in 1% of the specimens). The origins of the oldest species of the genus, namely L. pulchra, is within the costate taxa of the "Heterohelix" planata group of species as suggested by the general resemblances in the test architecture between the two species. A significant argument to support this evolutionary relationship is the direct observation of pore mounds aligned to the vestigial costae on the last chambers of rare specimens of L. pulchra (Figures 8.1c, 8.3c). The ontogenetic development of the two species shows gradual decrease in chamber size increase rate with the change in ornamentation (Figure 6); a similar process was previously reported in the Praegublerina pseudotessera–Gublerina rajagopalani lineage (Georgescu et al., 2009). Laeviheterohelix flabelliformis Nederbragt, 1991, which was described from the Turonian of Tunisia, lacks pore mounds on the chamber surface and for this reason is not considered a Laeviheterohelix species. The systematic position of this species will be analyzed in a forthcoming study; most likely L. flabelliformis is a junior synonym of Gublerina prima De Klasz, Le Calvez and Rerat, 1968 of the Turonian of Gabon.
Stratigraphic range. Upper Coniacian–Campanian (from the upper part of Whiteinella baltica Biozone throughout the Globigerinelloides impensus Biozone, the latter being the biostratigraphical equivalent of Radotruncana calcarata Biozone).
Geographic distribution. North America (United States, Canada), South Atlantic Ocean (Falkland Plateau), Western Europe (Sweden, Denmark, Germany, France, Romania) and North Africa (Tunisia).
Laeviheterohelix pulchra (Brotzen, 1936) – emended
(Figures 7.1–7.5, Figures 8.1–8.3)
Guembelina pulchra Brotzen, 1936, p. 121, pl. 9, fig. 3 (only).
Heterohelix pulchra (Brotzen). Pessagno, 1967, p. 262, pl. 87, fig. 4.
Heterohelix pulchra (Brotzen). Douglas and Rankin, 1969, p. 190, fig. 4.
Heterohelix pulchra (Brotzen). McNeil and Caldwell, 1981, p. 239, pl. 19, fig. 3.
Heterohelix pulchra (Brotzen). Weiss, 1983, pl. 2, figs. 10–13 (only).
Heterohelix pulchra (Brotzen). Nederbragt, 1989, p. 198, pl. 3, figs. 3–4.
Laeviheterohelix pulchra (Brotzen). Nederbragt, 1991, p. 352, pl. 5, fig. 7, pl. 6, fig. 1.
Laeviheterohelix turgida Nederbragt, 1991, p. 354, pl. 6, figs. 2–4.
Laeviheterohelix pulchra (Brotzen). Georgescu, 2006, fig. 4.17–19.
Emended description. Test is biserial throughout consisting of 15 to 18 chambers (proloculus included). Proloculus is small (6.4–10.4 µm in diameter). Earlier chambers are globular to subglobular, the last–formed ones reniform, tilted to the test axis of growth. Sutures are straight to slightly curved, depressed and oblique. Periphery is broadly rounded, without peripheral structures. Aperture is arch–shaped, situated at the base of the last–formed chamber and bordered by two well–developed, symmetrical periapertural flanges. Chamber surface is ornamented with small pore mounds (D = 1.7–2.8 µm, and rarely larger, up to 3.9 µm in less than 1% of the total number of measured structures; average value: 1.91 µm). Vestigial costae, which are collinear with the pore mounds, can be occasionally present on the last–formed chambers. Periapertural pustulose area is well–developed. Test wall is calcitic, hyaline, nannoperforate to microperforate, with pore diameter between 0.38–0.84 µm.
Remarks. Laeviheterohelix pulchra differs from Heterohelix planata (Cushman, 1938) by the test surface ornamented with pore mounds rather than fine costae.
Stratigraphic range. Upper Coniacian–Campanian (from the upper part of Whiteinella baltica Biozone throughout the Globigerinelloides impensus Biozone, the latter being the biostratigraphical equivalent of Radotruncana calcarata Biozone). The reviewed stratigraphic range confirms the observation by Douglas and Rankin (1969, p. 191) that this species occurs in "...Coniacian and younger strata".
Geographic distribution. North America (United States, Canada), South Atlantic Ocean (Falkland Plateau), Western Europe (Sweden, Denmark, Germany, France, Romania) and North Africa (Tunisia).
Laeviheterohelix reniformis (Marie, 1941) – emended
(Figure 8.4)
Ventilabrella reniformis Marie, 1941, p. 264, pl. 28, fig. 277.
Laeviheterohelix reniformis (Marie). Georgescu et al., 2009, pl. 2: D1–2.
Emended description. Test with early biserial stage and with incipient multiserial stage when adult. Earlier chambers are globular to subglobular, those of the multiserial stage reniform or tubular. Proloculus is small (10.7–12.8 µm). Sutures are straight to slightly curved, depressed and oblique to the test axis of growth. Periphery is broadly rounded, without peripheral structures. Aperture is multiple in the adult incipient multiserial stage; the shape is that of a low arch bordered by symmetrically developed periapertural flanges. Chamber surface is ornamented with pore mounds (D = 3.2–6.0 µm). There are well developed periapertural pustulose areas in the upper (anterior) part of the chambers. Test wall is calcitic, hyaline, microperforate to finely perforate, with pore diameter between 0.59–1.19 µm.
Remarks. Laeviheterohelix reniformis is a rare species, which differs from L. pulchra by the adult stage with chamber proliferation, chamber surface with larger pore mounds (3.2–6.0 µm rather than 1.7–2.8 µm) and larger pores (0.59–1.19 µm rather than 0.38–0.84 µm).
Stratigraphic range. Campanian (from the uppermost part of Marginotruncana marginata Biozone throughout Radotruncana calcarata Biozone).
Geographic distribution. South Atlantic Ocean (Falkland Plateau) and Western Europe (France).
Genus Steineckia n. gen.
Type species. Steineckia steinecki n. sp.
Diagnosis. Test is nannoperforate to microperforate, with large pore mounds and without periapertural pustulose area.
Description. Test is biserial throughout. Earlier chambers are globular, the last–formed ones reniform. Periphery is broadly rounded, without peripheral structures. Aperture is an arch at the base of the last–formed chamber and is bordered by two well–developed symmetrical flanges. Chamber surface ornamented is with large pore mounds. Test wall is calcitic, hyaline, nannoperforate to microperforate.
Species included.
Steineckia steinecki n. sp.
Remarks. Steineckia differs from Laeviheterohelix – emended by the absence of a periapertural pustulose area; it also lacks the incipient multiserial stage developed in the evolved species of Laeviheterohelix (i.e., L. reniformis). The origins of Steineckia are unknown. The absence of vestigial costae suggests that its ancestor is a taxon with smooth test surface.
Etymology. Genus named in honor of Dr. P. Lewis Steineck (Purchase, State University of New York) for his contributions in the development of the evolutionary classification of the Cenozoic planktic foraminifera.
Stratigraphic range. Upper Turonian (lower part of the Whiteinella baltica Biozone).
Geographic distribution. South Atlantic (Falkland Plateau).
Steineckia steinecki n. sp.
(Figures 9.1–9.5)
Holotype. Specimen WKB–F00501.
Dimensions of the holotype. Length: L = 0.169 mm; width: W = 0.116 mm; W/L = 0.686; thickness: T = 0.058 mm; T/L = 0.343.
Paratypes. Seven specimens WKB–000502.
Dimensions. L = 0.139–0.176 mm; W = 0.083–0.133 mm; W/L = 0.597–0.769; T = 0.058–0.065 mm; T/L = 0.343–0.387. Ranges are based on measurement of 14 specimens (paratypes and topotypes).
Material. Over 300 specimens.
Type locality. DSDP Site 511, Falkland Plateau. Geographic coordinates: 51° 00' S, 47°58' W.
Type level. Upper Turonian, Sample 79–511–47–5, 13.5–14.5 cm.
Etymology. As for the genus.
Description. Test is biserial throughout, consisting of 10 to 11 chambers that increase gradually in size; earlier chambers are globular, the last formed ones reniform. Proloculus is small (9.8–16.3 µm). Sutures are straight to slightly curved, depressed and oblique to the test axis of growth. Periphery is broadly rounded, without peripheral structures. Aperture is a medium–high arch situated at the base of the last–formed chamber and is bordered by two well–developed, symmetrical flanges. Chamber surface is ornamented with large pore mounds (D = 4.0–6.4 µm, rarely as small as 3.3 µm in juvenile specimens in less than 4% of the measured structures). No periapertural pustulose area is present. Test wall is calcitic, hyaline, nannoperforate to microperforate, pore diameter ranging between 0.43 and 0.76 µm.
Remarks. Ontogenetic trajectories based on chamber increase in size apparently indicate that the ancestor of Steineckia steinecki is among the smooth small–sized heterohelicid species of the Cenomanian and Turonian (Figure 10). The diameters of the pore mounds in S. steinecki (D = 4.0–6.4 µm, rarely as small as 3.3 µm) are much larger than those of the biserial L. pulchra (D = 1.7–2.8 µm, occasionally as large as 3.9 µm). However, there is an overlap area between the two taxa, which might suggest the existence of an evolutionary relationship between them. The absence of costae in S. steinecki and presence of vestigial costae in L. pulchra indicate different evolutionary origins for the two taxa.
Stratigraphic range. Upper Turonian (lower part of the Whiteinella baltica Biozone).
Geographic distribution. South Atlantic (Falkland Plateau).
Genus Fleisherites n. gen.
Type species. Guembelina glabrans Cushman, 1938.
Diagnosis. Test is nannoperforate to microperforate, smooth or with weak costae and with periapertural pustulose area.
Description. Test is biserial throughout. Earlier chambers are globular, then subrectangular, the last–formed reniform to subcircular. Test is laterally compressed and with subacute periphery as seen in edge view. Aperture is arch–shaped at the base of the last–formed chamber and is bordered by two well–developed, often rimmed, symmetrical flanges. Test is smooth or presents weak costae, often on the earlier portion and with well–developed periapertural pustulose area. Test wall is calcitic, hyaline, nannoperforate to microperforate.
Species included.
Guembelina glabrans Cushman, 1938, p. 15, pl. 3, figs. 1–2.
Remarks. Fleisherites n. gen. is restricted to the Late Cretaceous [uppermost Campanian (?) to Maastrichtian] and includes only taxa with a smooth test wall or, rarely, weak incipient costae and without pore mounds. Periapertural pustulose area is well–developed in all the studied specimens. Additional pustulose zones can be developed along the sutures due to imperfect overlapping of the periapertural pustulose areas of the previously formed chambers by the newly added ones (Figure 11). Fleisherites differs from Steineckia and Laeviheterohelix – emended by the absence of the pore mounds on the chamber surface. The origins of Fleisherites are unknown. Incipient costate ornamentation is often observed on the earlier part of the test (Figure 12.6a) and demonstrates that its ancestor is a smooth taxon (Figure 13), but no costate species with a laterally compressed test has been previously described. The incipient costate ornamentation in Fleisherites presents major differences when compared to those in Laeviheterohelix pulchra. Vestigial costae are present only on the last–formed chambers in the latter species and moreover, the test ultrastructure shows that the costae are the ancestral structures of the pore mounds. These further documents that L. pulchra and F. glabrans have different evolutionary origins and belong to different lineages and therefore, should be included in distinct genera.

Etymology. Genus named in the honor of Dr. Robert L. Fleisher (Micropaleontology Press) for his contributions to the development of the evolutionary classification of the Cenozoic planktic foraminifera.
Stratigraphic range. Upper Campanian (?), Maastrichtian (from the upper part of the Gansserina gansseri Biozone throughout the top of the Pseudoguembelina hariaensis Biozone).
Geographic distribution. Cosmopolitan.
Fleisherites glabrans (Cushman, 1938) – emended
Guembelina glabrans Cushman, 1938, p. 15, pl. 3, figs. 1–2.
Heterohelix glabrans (Cushman). Olsson, 1960, p. 26, pl. 4, fig. 4.
Heterohelix glabrans (Cushman). Pessagno, 1967, pl. 88, figs. 1–2, 10–11.
Heterohelix dentata Stenestad, 1968, p. 67, pl. 1, figs. 3–6, 8–9, pl. 2, figs. 1–3.
Heterohelix glabrans (Cushman). Weiss, 1983, pl. 1, figs. 6–7.
Laeviheterohelix glabrans (Cushman). Nederbragt, 1991, p. 352, pl. 5, fig. 6.
Emended description. Test with or without early incipient planispiral coil followed by the adult stage, which has biserial chamber arrangement. Proloculus is small (diameter, 12.8–16.7 µm). Chambers are subrectangular to subcircular increase gradually in size. Sutures are straight to slightly curved, depressed and oblique to the test axis of growth. Periphery is subangular and without peripheral structures. Aperture is in the shape of an arch at the base of the last–formed chamber and bordered by two symmetrical and well–developed periapertural flanges. Chamber surface is generally smooth, but scattered pustules and vestiges of the ancestral costate ornamentation can be occasionally observed on the earlier portion of the test. The only constantly ornamented zone of the test is the periapertural pustulose area. Test wall is calcitic, hyaline, nannoperforate to microperforate, pore diameter ranging between 0.37–0.76 µm; most of the pore diameters (over 80%) are concentrated within the 0.37–0.61 µm range.
Remarks. The ontogenetic trajectories of the type species of Laeviheterohelix and Fleisherites have different patterns in the adult stages, namely higher rate of increase in chamber size in F. glabrans (Figure 14). Fleisherites glabrans requires revision by means of detailed observations on the test ultrastructure. Apparently the species has wide morphological variability with respect to the chamber shape (i.e., subrectangular to subcircular) and sutures (i.e., straight to slightly curved). It is noteworthy that the holotype selected by Cushman (1938, pl. 3, fig. 1) and refigured here (Figures 11 and 12.3) is clearly at the middle between any of the end members of the chamber and suture shapes. Laeviheterohelix dentata (Stenestad 1968) is considered a junior synonym of F. glabrans, being described to accommodate the test variety with very well developed flanges resulting in a strongly dentate central suture; this interpretation is also supported by the similar stratigraphical ranges of the two species.
Stratigraphic range. Upper Campanian (?), Maastrichtian (from the upper part of the Gansserina gansseri Biozone throughout the top of the Pseudoguembelina hariaensis Biozone).
Geographic distribution. Cosmopolitan.
DISCUSSION AND CONCLUSIONS
The genus Laeviheterohelix was erected by Nederbragt (1991) to include the biserial planktic foraminifera with a smooth chamber surface or ornamented with pore mounds of the Turonian–Maastrichtian stratigraphic interval. A critical morphological review of the species originally included in Laeviheterohelix in the light of evolutionary classification shows that the genus is polyphyletic (Figure 15). Three distinct origins can be inferred for the taxa with smooth chamber surface or ornamented with pore mounds and consequently, three genera are recognized: Steineckia n. gen., Laeviheterohelix Nederbragt, 1991– emended and Fleisherites n. gen.
The earliest tests with pore mounds developed on the chamber surface are recorded in the upper Turonian of the southern hemisphere (Falkland Plateau). They are formalized as Steineckia steinecki n. gen., n. sp. This taxon exhibits large pore mounds (D = 4.0–6.4 µm, rarely as small as 3.3 µm) on the chamber surface and lacks periapertural pustulose area. Apparently it evolved from a smooth taxon. This is strongly supported by the absence of vestigial costae on the chamber surface (Figure 16).
Laeviheterohelix Nederbragt, 1991 is emended and it includes two species: L. pulchra (Brotzen, 1936) and L. reniformis (Marie, 1941). The stratigraphic range of the emended Laeviheterohelix is upper Coniacian–Campanian. Species included within Laeviheterohelix – emended present periapertural pustulose area. The earlier and therefore primitive species, L. pulchra has completely biserial tests; the pore mounds present small diameters, 1.7–2.8 µm, and rarely larger, up to 3.9 µm and the test wall is nannoperforate to microperforate (pore diameter ranges between 0.38–0.84 µm). Laeviheterohelix reniformis apparently evolved from L. pulchra and is the only species of the genus with incipient chamber proliferation in the adult stage (Figure 16); this species presents larger pore mounds (D = 3.2–6.0 µm) and microperforate to finely perforate test wall (pore diameter ranges between 0.59–1.19 µm), in contrast with its ancestor.
Maastrichtian serial planktic foraminiferal species, which were originally included in the genus Laeviheterohelix by Nederbragt (1991), do not have pore mounds on the chamber surface. Such tests were recorded from the upper part of Gansserina gansseri Biozone throughout the Cretaceous/Tertiary boundary and are formalized at the genus level as Fleisherites n. gen. This genus includes only F. glabrans (Cushman, 1938), a biserial species that occasionally has an early incipient planispiral coiling. These tests consistently develop a periapertural pustulose area. Incipient costate ornamentation could be observed on nearly half of the specimens examined; when present, the costae are situated mostly on the earlier portion of the test.
This new taxonomic framework reveals that the morphological differences between various heterohelicid taxa are much smaller than previously thought. Observations on the representatives of the genus Laeviheterohelix show that tests with relatively close morphological features were achieved through iterative evolutionary processes (Figure 17). This evolution pattern was originally observed in the case of Cenozoic planktic foraminifera: "Pervasive convergent and repetitive evolution precludes existing classifications based solely on morphologic similarity from reflecting evolutionary relationship." (Steineck and Fleisher, 1978, p. 618). Accordingly, it appears that a closer morphological analysis combined with biostratigraphic and paleobathymetric data is necessary in order to recognize the natural, monophyletic taxonomic units.
ACKNOWLEDGMENTS
Dr Brian T. Huber (National Museum of Natural History, Smithsonian Institution, Washington, D.C.) is thanked for the help and full access to collections and research facilities during my 2005–2006 stay at the NMNH. The author thanks Drs K. A. Ansdell and W. K. Braun (University of Saskatchewan at Saskatoon, Canada) for the access to research facilities during my short stay in 2007. Dr Leonard V. Hills (University of Calgary, Alberta, Canada) is kindly thanked for the presubmittal review of the manuscript. The manuscript was reviewed by Dr Danièle Grosheny (Université du Strasbourg, France) and an anonymous reviewer. Mr I. Levya (University of Calgary) is kindly thanked for preparing the Spanish abstract. Last but not least, the author warmly thanks Drs María del Carmen Perrilliat and Carlos M. González León (Universidad Nacional Autónoma de México) for accepting the publication of the manuscript in Revista Mexicana de Ciencias Geológicas.
REFERENCES
Abramovich, S., Keller, G., Stüben, D., Berner, Z., 2003, Characterization of late Campanian and Maastrichtian planktic foraminiferal depth habitats and vital activities based on stable isotopes: Palaeogeography, Palaeclimatology, Palaeoecology, 202, 1–29. [ Links ]
Brotzen, F., 1936, Foraminiferen aus dem schwedischen unterer Senon von Eriksdal in Schonen: Sveriges Geologiska Untersökning, ser. C, 396(30), 1–69. [ Links ]
Brown, N.K., Jr., 1969, Heterohelicidae Cushman, 1927, amended, a Cretaceous planktic foraminiferal family, in Brönnimann, P., Renz, H.H. (eds.), Proceedings of the First International Conference on Planktic Microfossils, Geneva 1967: Leiden, E. J. Brill, 2, 21–67. [ Links ]
Cushman, J.A., 1927, An outline of a re–classification of the Foraminifera: Contributions from the Cushman Foundation for Foraminiferal Research, 3, 1–105. [ Links ]
Cushman, J.A., 1938, Cretaceous species of Guembelina and related genera: Contributions from the Cushman Laboratory for Foraminiferal Research, 14, 2–27. [ Links ]
De Klasz, I., Le Calvez, Y., Rerat, D., 1968, Nouveaux foraminifères du basin sédimentaire du Gabon (Afrique Équatoriale), in Said, R., El Ansary, S., Viotti, C., Ghorab, M.R. (eds.), Proceedings of the Third African Micropaleontology Colloquium, Cairo, March 4–10, 1968: 269–287. [ Links ]
Delage, Y., Hérouard, E., 1896, Traité de zoologie concrete. Tome I: La cellule et les protozoaires: Paris, Schleicher Frères, 584 p. [ Links ]
Douglas, R.G., Upper Cretaceous planktonic foraminifera in northern California. Part 1 – Systematics: Micropaleontology, 15, 151–209. [ Links ]
Douglas, R.G., Rankin, C., 1969, Cretaceous planktic foraminifera from Bornholm and their zoogeographic significance: Lethaia, 2, 185–217. [ Links ]
Egger, J.G., 1902, Foraminiferen und Ostrakoden aus den Kreidemergeln der oberbayerischen Alpen: Abhandlungen der königlichen bayerischen Akademie der Wissenschaften zu München, Mathematische–Naturwissenschaftlische Klasse, 21, 3–230. [ Links ]
Ehrenberg, C.G., 1843, Verbreitung und Einfluss des mikrosckopischen Lebens in Süd und Nord America: Abhandlungen der königlichen preussischen Akademie der Wiessenschaften zu Berlin (1841), 1, 291–446. [ Links ]
Eichwald, C.E., von, 1830, Zoologia specialis II: Vilnae, D. E. Eichwaldus, 323 p. [ Links ]
Georgescu, M.D., 2006, Santonian–Campanian planktic foraminifera from the New Jersey coastal plain and their distribution related to the relative sea–level changes: Canadian Journal of Earth Sciences, 43, 101–120. [ Links ]
Georgescu, M.D., 2007a, Taxonomic re–evaluation of the Late Cretaceous serial planktic foraminifer Guembelina punctulata Cushman, 1938 and related species: Revista Española de Micropaleontologia, 39, 155–167. [ Links ]
Georgescu, M.D., 2007b, A new planktic heterohelicid foraminiferal genus from the Upper Cretaceous (Turonian): Micropaleontology, 53, 212–220. [ Links ]
Georgescu, M.D., Abramovich, S., 2008a, Taxonomic revision and phylogenetic classification of the Late Cretaceous (upper Santonian–Maastrichtian) serial planktic foraminifera (Family Heterohelicidae Cushman 1927) with peripheral test wall flexure: Revista Española de Micropaleontologia, 40, 97–114. [ Links ]
Georgescu, M.D., Abramovich, S., 2008b, A new serial Cretaceous planktic foraminifer (Family Heterohelicidae Cushman 1927) from the upper Maastrichtian of the equatorial Central Pacific: Journal of Micropaleontology, 27, 117–123. [ Links ]
Georgescu, M.D., Huber, B. T., 2007, Taxonomic revision of the late Campanian–Maastrichtian (Late Cretaceous) planktic foraminiferal genus Rugotruncana Brönnimann and Brown, 1956, and a new paleontological species concept for planktic foraminifera: Journal of Foraminiferal Research, 37, 150–159. [ Links ]
Georgescu, M.D., Huber, B.T., 2008, Taxonomic re–evaluation and phylogeny of the stellate planktic foraminiferal genus Hastigerinoides Brönnimann, 1952: Journal of Foraminiferal Research, 38, 52–58. [ Links ]
Georgescu, M.D., Saupe, E.E., Huber, B.T., 2009, Taxonomic revision of the Late Cretaceous (late Campanian–Maastrichtian) serial planktic foraminiferal genus Gublerina Kikoïne, 1948: Micropaleontology, 54, 397–424. [ Links ]
Gradstein, F.M., Ogg, J.G., Smith, A.G. (eds.), 2004. A Geologic Time Scale 2004: Cambridge, Cambridge University Press, 589 p. [ Links ]
Hay, W.H., DeConto, R.M., Wold, C.N., Wilson, K.M., Voigt, S., Schulz, M., Wold, A.R., Dullo, W.C., Ronov, A.B., Balukhovsky, A.N., Söding, E., 1999, Alternative global Cretaceous paleogeography, in Barrera, E., Johnson, C.C. (eds.), Evolution of the Cretaceous Ocean–Climate System: The Geological Society of America, Special Paper 332, 1–47. [ Links ]
Hofker, J., 1957, Foraminiferen der Oberkreide von Nordwestdeutschland und Holland: Beihefte zum Geologischen Jahrbuch, 27, 1–464. [ Links ]
Huber, B.T., 1991, Planktic foraminifer biostratigraphy of Campanian–Maestrichtian sediments from Sites 698 and 700, southern South Atlantic, in Ciesielski, P.F., Kristoffersen. Y. et al. (eds.), Proceedings of the Ocean Drilling Program, Scientific Results: College Station, Texas, 114, 281–297. [ Links ]
Huber, B.T., 1992, Upper Cretaceous planktic foraminiferal biozonation for the Austral Realm: Marine Micropaleontology, 20, 107–128. [ Links ]
Huber, B.T. (coordinator)–Mesozoic Planktic Foraminiferal Working Group, 2006, Mesozoic Planktic Foraminiferal Taxonomic Dictionary, <http://portal.chronos.org/gridsphere/gridsphere?cid=resources>. [ Links ]
Huber, B.T., Hodell, D.A., Hamilton, C.P., 1995, Middle–Late Cretaceous climate of the southern high latitudes: stable isotopic evidence for minimal equator–to–pole thermal gradients: Geological Society of America Bulletin, 107, 1164–1191. [ Links ]
Loeblich, A.R.,Jr., Tappan, H., 1961, The genera Microaulopora Kuntz, 1895, and Guembelina Kuntz, 1895, and the status of Guembelina Egger, 1899: Journal of Paleontology, 35, 625–627. [ Links ]
Loeblich, A.R.,Jr., Tappan, H., 1987. Foraminiferal Genera and Their Classification: New York, van Nostrand Reinhold Company, 970 p. [ Links ]
Marie, P., 1941, Les foraminifères de la craie a Belemnitella mucronata du Bassin de Paris: Mémoires du Muséum National d'Histoire Naturelle, 12, 1–296. [ Links ]
Masters, B.A., 1977, Mesozoic Planktic Foraminifera: a world–wide review and analysis. in Ramsay, A.T.S. (ed.), Oceanic Micropaleontology: Academic Press, 1, 301–733. [ Links ]
McNeil, D.H., Caldwell, W.G.E., 1981, Cretaceous Rocks and Their Foraminifera in the Manitoba Escarpment: The Geological Association of Canada, Special Paper 21, 1–439. [ Links ]
Montanaro Gallitelli, E., 1957, A revision of the foraminiferal Family Heterohelicidae, in Loeblich, A.R.,Jr. (ed.), Studies in Foraminifera: United States National Museum Bulletin, 215, 133–154. [ Links ]
Nederbragt, A.J., 1989, Maastrichtian Heterohelicidae (Planktic Foraminifera) from the North West Atlantic: Journal of Micropaleontology, 8, 183–206. [ Links ]
Nederbragt, A.J., 1991, Late Cretaceous biostratigraphy and development of Heterohelicidae (planktic foraminifera): Micropaleontology, 37, 329–372. [ Links ]
Olsson, R.K., 1960, Foraminifera of latest Cretaceous and earliest Tertiary age in the New Jersey Coastal Plain: Journal of Paleontology, 34, 1–58. [ Links ]
Pessagno, E.A.,Jr., 1967, Upper Cretaceous planktic foraminifera from the Western Gulf Coastal Plain: Palaeontographica Americana, 5(37), 245–445. [ Links ]
Petrizzo, M.R., 2001, Late Cretaceous planktic foraminifera from Kerguelen Plateau (ODP Leg 183): new data to improve the southern Ocean biozonation: Cretaceous Research, 22, 829–855. [ Links ]
Premoli Silva, I., Sliter, V.W., 1995, Cretaceous planktic foraminiferal biostratigraphy and evolutionary trends from the Bottaccione section, Gubbio, Italy: Palaeontographia Italica, 82, 1–89. [ Links ]
Robaszynski, F., Caron, M., 1995, Foraminifères planctoniques du Crétacé: commentaire de la zonation Europe–Mèditerranée: Bulletin de la Société Géologique de France, 6, 681–692. [ Links ]
Steineck, P.L., Fleisher, R.L., 1978, Towards the classical evolutionary reclassification of Cenozoic Globigerinacea (Foraminiferida): Journal of Paleontology, 52, 618–635. [ Links ]
Stenestad, E., 1968, Three new species of Heterohelix Ehrenberg from the upper Senonian of Denmark: Meddelelser fra Dansk Geologisk Forening, 18, 64–70. [ Links ]
Tappan, H., 1940, Foraminifera from the Grayson Formation of northern Texas: Journal of Paleontology, 14, 93–126. [ Links ]
Thomas, N.L., 1927, The use of evolutionary changes in geologic correlation: Journal of Paleontology, 1, 135–140. [ Links ]
Thomas, N.L., Rice, E.M., 1927, Changing characters in some Texas species of Guembelina: Journal of Paleontology, 1, 141–145. [ Links ]
Weiss, W. 1983, Heterohelicidae (seriale planktonische Foraminiferen) der tethyalen Oberkreide (Santon bis Maastricht): Geologisches Jahrbuch, A72: 1–96. [ Links ]
Zapeda, M.A., 1998, Planktic foraminiferal diversity, equitability and biostratigraphy of the uppermost Campanian–Maastrichtian, ODP Leg 122, Hole 762C, Exmouth Plateau, NW Australia, eastern Indian Ocean: Cretaceous Research, 19, 117–152. [ Links ]

