











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Los estudios sobre biología reproductiva que abordan aspectos como la temporalidad del desove y la talla de madurez sexual aportan información muy útil para propiciar un mejoramiento en el manejo pesquero (Stearns, 2000), además, son de los aspectos de mayor importancia para profundizar en el conocimiento de la dinámica poblacional (Jennings et al., 1999).
En general, los ciclos reproductivos de los invertebrados marinos responden a cambios en el medio ambiente; entre los más importantes, están la temperatura y la disponibilidad de alimento (Darriba et al., 2004; Dridi et al., 2007; Enríquez-Díaz et al., 2009). Sin embargo, el proceso reproductivo también puede verse afectado como consecuencia de cambios en un medio marino contaminado (Vaschenko et al., 2013). En la región costera del golfo de California el ambiente sedimentario del puerto de Santa Rosalía es receptor y fuente de contaminantes inorgánicos; se han comprobado anomalías positivas de concentración de elementos no esenciales en los sedimentos de dicha zona, con potencial de toxicidad y bio-concentración (Shumilin et al., 2000; Rodríguez-Figueroa et al., 2009; Shumilin et al., 2011, Shumilin et al., 2012, Shumilin et al., 2013). Precisamente, en ese puerto minero se reportó recientemente una deteriorada condición de salud en las gónadas de la almeja chocolata Megapitaria squalida (Sowerby, 1835) (Yee-Duarte et al., 2018). Lo anterior demuestra que es posible llegar a inducir cambios patológicos en los bivalvos, los cuales, por ser organismos filtradores, suelen ser más susceptibles a los agentes ambientales, donde las anomalías en los tejidos como la presencia de granulocitomas o lesiones constituidas principalmente por granulocitos (Lowe & Moore, 1979) tienen relevancia diagnóstica como biomarcadores de exposición a la contaminación (Neff et al., 1987; Auffret, 1988; Villalba et al., 1997; Usheva et al., 2006; Kim & Powell, 2007; Garmendia et al., 2011). Algo parecido ocurre con la inflamación, pues a pesar de ser parte funcional de la respuesta inmune de un organismo (Muller, 2003), también se ha analizado su posible uso como biomarcador en exposiciones a los contaminantes, tanto agudas como subletales a largo plazo (Garmendia et al., 2011).
La almeja roñosa Chione undatella (Sowerby, 1835) es un recurso potencial pesquero y de cultivo que se ha explotado intensivamente durante mucho tiempo con poca o ninguna regulación, en la costa del Pacífico mexicano (Arizpe-Covarrubias, 1992). Esta almeja forma bancos en áreas someras y se entierra a poca profundidad en fondos blandos de sedimentos finos (Fisher et al., 1995), lo que la hace de fácil acceso. En este trabajo se estudió una población de la almeja roñosa y se determinó su ciclo reproductivo, su temporalidad de desove y su talla de primera madurez. Adicionalmente, se reporta una anomalía en el tejido gonadal que podría estar asociada con la exposición a los contaminantes presentes en el área costera de Santa Rosalía.
Materiales y métodos
Se realizaron muestreos mensuales en un ambiente costero adyacente al puerto minero de Santa Rosalía, localizado en la costa oriental del golfo de California (27° 20’ N y 112° 16’ O), durante el periodo de septiembre de 2014 a agosto 2015, excepto en marzo de 2015, cuando no fue posible obtener organismos. De cada organismo se registró la longitud de la concha (distancia en línea recta desde el margen anterior al margen posterior de la concha (LC; 0.1 mm de precisión), así como los pesos: total, sin concha y de la concha (0.1 g de precisión), y se calculó el índice de condición general (ICG), como la razón porcentual del peso sin concha y el peso total (Góngora-Gómez et al., 2016).
De cada organismo se extrajo tejido gonadal, el cual fue fijado en formol al 10% en agua de mar. Después de lavar el exceso de fijador, el tejido se procesó histológicamente (Humason, 1979), se deshidrató por medio de una serie de alcoholes de concentraciones crecientes (70-100%) y se aclaró con HemoDe® y con inclusión en Paraplast®. Se realizaron cortes de 5 µm de grosor y se tiñeron con la técnica de hematoxilina-eosina, con la que se obtiene un contraste adecuado de las estructuras del tejido.
El sexo (macho o hembra) se identificó mediante el análisis microscópico. Para evaluar la proporción de sexos en la muestra se aplicó una prueba de ji-cuadrada con la corrección de continuidad de Yates (χc2) bajo la hipótesis nula de que existe una proporción 1:1 (Zar, 1996). La regla de decisión se realizó con una confianza del 95%, sin aceptar la hipótesis cuando el valor de χc2 calculado fuera mayor a 3.86 (Sokal & Rohlf, 1979).
La descripción del ciclo reproductivo se realizó a través de la estimación de las frecuencias relativas mensuales de cada fase de desarrollo gonádico (Tabla 1). Durante el análisis microscópico se detectaron infiltraciones hemocíticas en el tejido gonadal y, al considerar el origen contaminado de las muestras, se aplicó el índice cualitativo propuesto por Bignell et al. (2011). Primero, con una estimación visual, se ubicaron las muestras en etapas (ausente, presente, intermedio o abundante) con base en la proporción de cobertura de células eosinófilas presentes en el tejido gonadal, de acuerdo con la clasificación que se presenta en la Tabla 2. Posteriormente, se clasificaron los organismos en cada etapa agrupándolos por sexo, mes y fase de desarrollo gonadal, para calcular los porcentajes y llevar a cabo las comparaciones entre ellas.
Tabla 1 Fases de desarrollo de las gónadas y los correspondientes valores del índice de condición general (media ± error estándar) de Chione undatella (Sowerby, 1835).
| Hembras | Machos | ||||
|---|---|---|---|---|---|
| Estadio de desarrollo gonádico | ICG | Observación microscópica | Estadio de desarrollo gonádico | ICG | Observación microscópica |
| Indiferenciado | 15.40 ± 0.13 | Presencia de acinos vacíos, no se observan gametos. No se puede diferenciar el sexo. El tejido conjuntivo es abundante. | |||
| Desarrollo inicial | 19.89 ± 1.31 | Ovocitos pedunculados unidos a las paredes de los folículos. En algunos casos se observan ovocitos residuales de un desove anterior (remaduración). | Desarrollo | 20.90 ± 0.62 | Una capa gruesa de espermatogonias y espermatocitos en los túbulos seminíferos, separados por tejido conectivo. |
| Desarrollo final | 20.30 ± 0.81 | Folículos con ovocitos en diferentes fases (vitelogénicos y pedunculados). Los ovocitos se encuentran unidos a la pared del folículo y unos pocos libres en el lumen. Una marcada reducción en la cantidad de tejido conectivo entre los folículos. | |||
| Madurez | 20.03 ± 0.71 | Folículos llenos de ovocitos libres en el lumen. Los núcleos y nucléolos de los ovocitos son visibles. Sin tejido conectivo entre folículos. | Madurez | 19.95 ± 0.56 | Crecimiento de los túbulos seminíferos, sin espacio entre ellos. Reducción en la cantidad de espermatogonias y espermatocitos con un aumento en la cantidad de espermatozoides visibles en el centro de los túbulos. |
| Desove | 20.52 ± 0.88 | Folículos parcialmente vacíos, con ovocitos libres en el lumen y presencia de fagocitos. | Espermiación | 19.99 ± 0.61 | Algunos túbulos con espermatozoides abundantes y otros parcialmente vacíos con las paredes rotas. |
| Recuperación | 19.77 ± 0.67 | Folículos vacíos, con algunos ovocitos residuales y actividad fagocitaria. | Recuperación | 19.34 ± 0.94 | Los túbulos presentan gametos residuales, actividad fagocitaria y abundante tejido conectivo entre ellos. |
Tabla 2 Gradiente cualitativo utilizado para evaluar la intensidad de la infiltración de células eosinófilas en el tejido gonadal de Chione undatella (Sowerby, 1835) (modificado de Bignell et al., 2011).
| Etapa | Proporción de células eosinófilas |
|---|---|
| 0 | |
| Ausente | Sin presencia de células eosinófilas en el tejido (0 %) |
| 1 | |
| Presente | Presencia de algunas células eosinófilas en el tejido (> 0 % ≤ 10 %) |
| 2 | |
| Intermedio | Presencia notable de células eosinófilas en el tejido (> 10 % ≤ 50 %) |
| 3 | |
| Abundante | Presencia abundante de células eosinófilas en el tejido (> 50 %) |
Mediante el modelo logístico y = a/(1+be-c*L), se obtuvo la talla de madurez sexual (L50%) al medir la longitud de la concha a la cual el 50% de los individuos son sexualmente maduros. Para esto se utilizó solamente a los organismos con actividad reproductiva, es decir, organismos en fase de madurez, desove/espermiación y recuperación.
Se obtuvieron datos de la temperatura superficial del mar (TSM) y la concentración superficial de clorofila-a desde el navegador en línea de la National Oceanic and Armospheric Administration (NOAA) “Coast Watch Browser”. Para ambas variables se calcularon los promedios mensuales de la zona aledaña a la dársena del puerto de Santa Rosalía, Baja California Sur, en el periodo de 2014 a 2015 en las coordenadas 27° 20’ 45.76’’ N, -112° 15’ 49.28’’ O.
Análisis estadísticos. Al aplicar una prueba de normalidad de Kolmogorov-Smirnov se encontró que los datos de tallas (KS = 0.14, p = 0.01), peso total (KS = 0.11, p = 0.02), peso sin concha (KS = 0.10, p = 0.04) e índice de condición (KS = 0.12, p = 0.04) no presentan una distribución normal, por lo que para detectar diferencias significativas de estas variables entre sexos se utilizó la prueba de Kruskal-Wallis. Debido a que los valores del índice de condición son porcentajes, se aplicó una transformación arcoseno (Zar, 1996) para normalizar la distribución de los datos y poder aplicar los análisis de correlación y cumplir con el supuesto de normalidad. Se utilizaron los promedios mensuales para realizar correlaciones de Spearman (análisis no paramétrico) entre las variables ambientales (temperatura superficial del mar y concentración superficial de clorofila a y el ICG, así como también con la fase de desarrollo de mayor frecuencia. Los análisis estadísticos se realizaron con un nivel de significancia de p < 0.05 con el programa Minitab 18.
Resultados
Se analizaron 176 organismos, de los cuales, 2 fueron indiferenciados (1%), 82 fueron machos (47%) y 92 fueron hembras (52%). La proporción sexual total fue 1.1H:1M, y sólo fue significativamente diferente de la proporción 1:1 en julio (χc 2 = 1.14; p < 0.05), donde hubo más hembras que machos (6H:1M).
Los valores medios, error estándar y rango de las variables morfométricas y del índice de condición general por sexo, así como los resultados de las pruebas de Kruskal-Wallis se presentan en la Tabla 3. Las hembras fueron significativamente más grandes que los machos, tanto en longitud de la concha (LC) como en peso total; sin embargo, al comparar el peso sin concha, esta diferencia desapareció y, de forma similar, en el ICG se equilibró la diferencia entre sexos. Al analizar la variabilidad mensual del ICG se observa la misma tendencia en ambos sexos, a pesar de que no hay una correlación significativa (Rho = 0.77; p = 0.005; N = 11; Fig. 1), ya que se registró un promedio alto en abril, tanto en hembras (23.4 ± 0.38) como en machos (21.45 ± 0.83), y otro en agosto en machos (23.37 ± 0.63) y en septiembre en hembras (24.55±2.62), hasta alcanzar los menores valores en octubre (14.45 ± 0.94 y 15.29 ± 1.29, hembras y machos respectivamente). Se encontró una correlación inversamente proporcional (Rho = -0.633; p = 0.010; N = 11) entre los promedios mensuales del ICG y la fase de desarrollo de mayor frecuencia en hembras, pero no en machos.
Tabla 3 Comparación de las variables morfométricas y del índice de condición general por sexo de Chione undatella (Sowerby, 1835) en Santa Rosalía, Baja California Sur, México. Se presentan media ± error estándar y el rango entre paréntesis.
| Machos | Hembras | Kruskal-Wallis | |
|---|---|---|---|
| N = 82 | N = 92 | g.l. 1 | |
| Longitud de la concha (mm) | 57.94 ± 0.50 | 58.85 ± 0.47 | H = 4.73 |
| (48-70) | (42-66) | p = 0.029 | |
| Peso total (g) | 62.81 ± 1.62 | 65.44 ± 1.44 | H = 5.57 |
| (38.5-112.5) | (23.9-94.4) | p = 0.018 | |
| Peso sin concha (g) | 12.59 ± 0.31 | 13.30 ± 0.35 | H = 3.47 |
| (7.6-20.3) | (4.1-20.3) | p = 0.063 | |
| Índice de condición general | 20.31 ± 0.33 | 20.47 ± 0.43 | H = 0.18 |
| (10.25-25.94) | (11.68-44.23) | p = 0.669 |
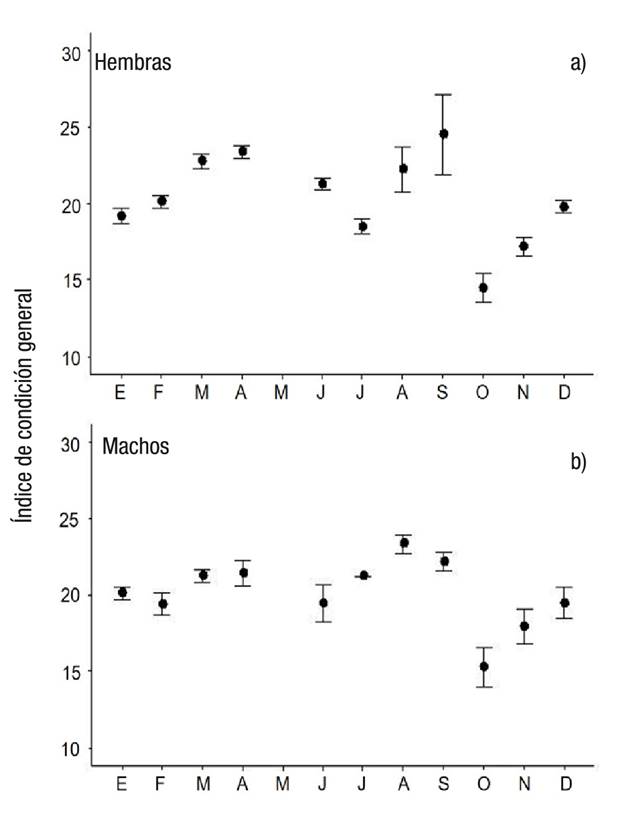
Figuras 1a-b Variación estacional del índice de condición general de hembras (a) y machos (b) de Chione undatella (Sowerby, 1835) en Santa Rosalía, Baja California Sur, México.
Se estimó la talla de madurez sexual (L50%) para machos en 57.01 mm LC (r2 = 0.98) y para hembras en 57.09 mm LC (r2 = 0.99), aunque la hembra más pequeña con actividad reproductiva fue de 42 mm LC y el macho de 50 mm LC (Fig. 2).
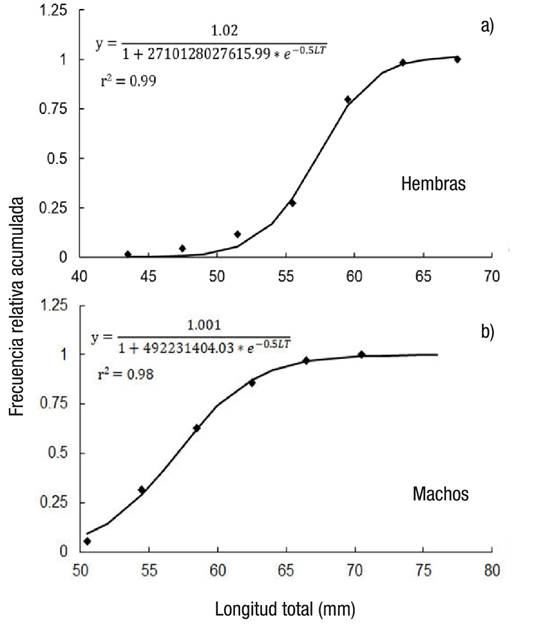
Figuras 2a-b Talla de madurez sexual (L50) para hembras (a) y machos (b) de Chione undatella (Sowerby, 1835) en Santa Rosalía, Baja California Sur, México. Se incluyen los modelos ajustados.
Análisis histológico. En el tejido gonadal se observó que al inicio del desarrollo la pared de los folículos estaba engrosada, las ovogonias y ovocitos se encontraban en crecimiento en las hembras, y los espermatocitos en los machos se hallaban embebidos en la pared de los folículos. Había tejido conectivo interfolicular muy evidente, el cual se reduce conforme se avanza en el proceso de maduración. De manera generalizada, se observan gametos en diferentes fases de desarrollo al mismo tiempo en un mismo individuo, lo que permite suponer que hay una producción continua de gametos y desoves parciales.
Se presenta actividad reproductiva durante todo el año, de manera más evidente en machos (Fig. 3). Las hembras maduras se presentan en mayor porcentaje de marzo a junio y en agosto (> 50%), y desovan en mayor porcentaje durante julio y septiembre (50 y 100%, respectivamente), pero presentan desove de menor intensidad durante enero y febrero (38 y 36%, respectivamente). Los machos maduros se observan prácticamente todo el año (excepto en septiembre y octubre) y registran un mayor porcentaje de espermiación entre octubre y enero, con mayor intensidad en octubre y noviembre (> 80%). Los organismos indiferenciados se presentaron en julio y en diciembre con tallas de 61 mm LC y 57 mm LC, respectivamente.
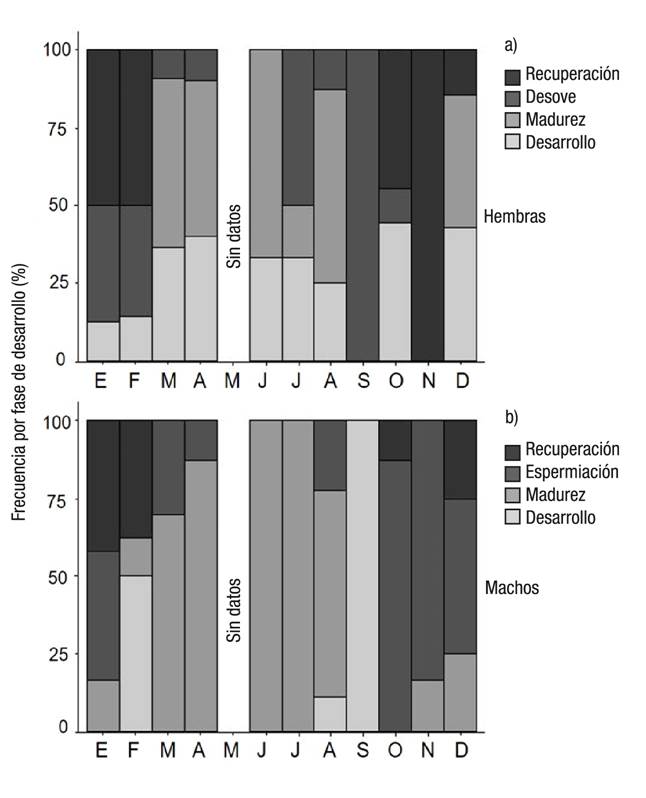
Figuras 3a-b Ciclo reproductivo de hembras (a) y machos (b) de Chione undatella (Sowerby, 1835) en Santa Rosalía, Baja California Sur, México.
Infiltración hemocítica en el tejido gonadal. Esta condición se presentó tanto en hembras como en machos con intensidades similares (Rho = 1.90; p = 0.168; N = 174), por lo que se analizaron los datos en conjunto. Adicionalmente, esta condición estuvo acompañada de una reducción y desaparición del tejido conectivo. Durante la mayor parte del año se observó una infiltración de células eosinófilas en el tejido gonadal en más del 80% de los individuos analizados (Fig. 4). La variación mensual de la prevalencia de organismos con infiltración de células eosinófilas en el tejido gonadal se presenta en la Figura 5a. Los meses cuando se observó una menor frecuencia de organismos con infiltración (53 y 47%) fueron agosto y octubre, mientras que la prevalencia de estas infiltraciones fue notable en enero y julio, cuando más del 85 % de los individuos presentaban infiltración con una intensidad en etapa 3 (abundante); mientras que en enero, marzo, julio, septiembre y noviembre el 100% de los organismos presentaban infiltraciones. Por otro lado, en las cuatro fases de desarrollo gonádico se presentó una intensidad de intermedia (etapa 2) a abundante (etapa 3) de infiltración de células eosinófilas en más del 60% de los organismos (Fig. 5b).
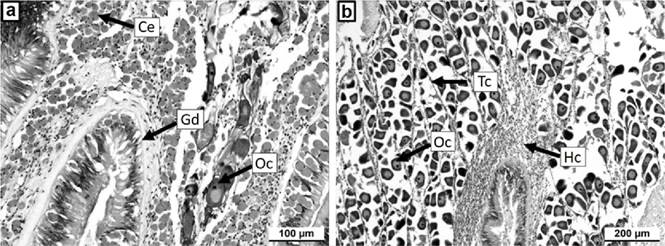
Figuras 4a-b Microfotografía de tejido gonadal de Chione undatella (Sowerby, 1835), teñido con hematoxilina-eosina. a) Tejido gonadal con infiltración de células eosinófilas (Ce), con algunos ovocitos (Oc), y parte de la glándula digestiva (Gd). b) Tejido gonadal en estado normal con abundantes ovocitos (Oc), con tejido conectivo (Tc) y células hemocíticas (Hc).
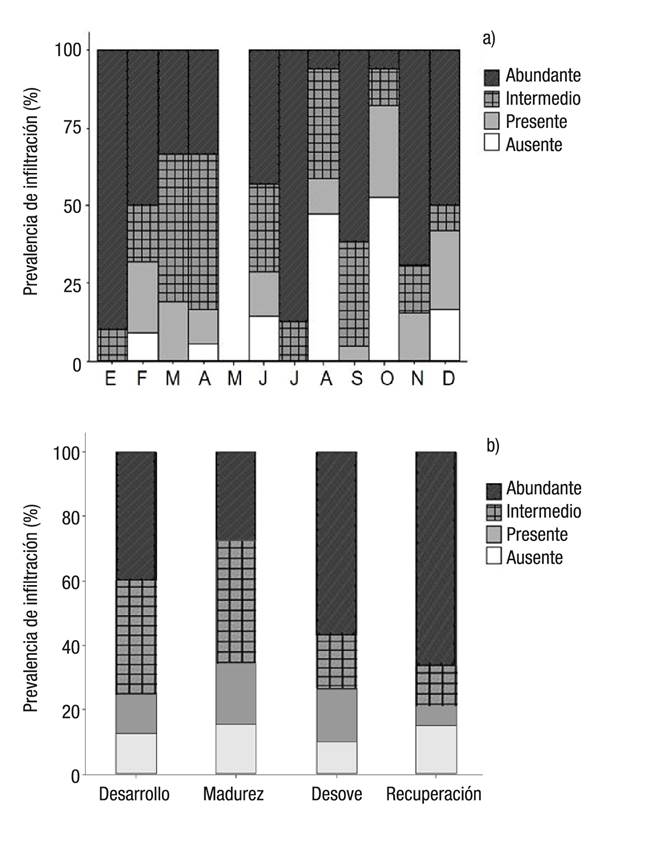
Figuras 5a-b Variación mensual (a) y por fase de desarrollo gonádico (b) de la prevalencia por etapa de intensidad de infiltración de células eosinófilas en el tejido gonadal de Chione undatella (Sowerby, 1835) (ambos sexos) en la zona costera de la región minera de Santa Rosalía. Baja California Sur, México.
Variables ambientales. Las variaciones mensuales de la temperatura superficial del mar y clorofila a se presentan en la Figura 6. Se registraron temperaturas del agua mayores a 25 ºC en el periodo de junio a octubre, agosto fue el mes más cálido con 30.2 ºC, en noviembre la temperatura del agua empezó a disminuier hasta llegar al menor promedio mensual en enero con 18.4 ºC. La concentración de clorofila-a mostró notables variaciones: tres periodos con valores altos, el máximo en marzo (47.66 mg/m3) seguido de septiembre (41.44 mg/m3) y el menor en junio (28.39 mg/m3). Julio fue el mes con la menor concentración de clorofila-a, y a que sólo alcanzo los 0.63 mg/m3. No se encontró correlación estadísticamente significativa entre el ICG con la construcción de clorifila a (Rho = -0.08; p = 0.81; N = 11), ni con la TSM (Rho = -0.01; p = 0.98; N = 11).

Discusión
En la región costera del golfo de California, el ambiente sedimentario del puerto de Santa Rosalía es receptor y fuente de contaminantes inorgánicos. En este sentido, se han reportado anomalías de concentración de elementos no esenciales en los sedimentos, que evidencian una potencial toxicidad y su bioconcentración en la macroalga Padina durvillaei Bory Saint-Vincent, 1827 (Rodríguez-Figueroa et al., 2009). La almeja roñosa C. undatella, al ser un organismo filtrador, está propensa a bioacumular metales pesados, como se ha documentado para otros organismos filtradores (Gutiérrez-Galindo et al., 1999; Cadena-Cárdenas et al., 2009), lo cual podría traer consecuencias adversas para el metabolismo. Asimismo, se reportó recientemente que en esta región minera la almeja chocolata M. squalida presenta indicios de una salud general deteriorada, que se observa en su menor tamaño y su baja condición (ICG < 20%), en comparación con otras zonas no contaminadas o contaminadas en menor grado (Yee-Duarte et al., 2017). En C. undatella los valores del ICG parecen estar relacionados con la fase de madurez (ver Tabla 1). En las hembras, por su parte, los valores mensuales de ICG estuvieron entre 24.5 ± 2.6 en septiembre y 14.4 ± 0.9 en octubre, pero esto parece estar más relacionado con el desove que con una mala condición de salud general. Al considerar específicamente la condición histopatológica de la gónada, se reportaron, en la misma zona minera donde se realizó el presente estudio, algunas alteraciones en las gónadas de M. squalida (ovocitos atrésicos, células cafés, infiltración de hemocitos, granulocitomas y parásitos) y se evidenció una gran presencia de cobre (30%) y hierro (45%), que se midió como área de cobertura en el corte del tejido de la gónada (Yee-Duarte et al., 2018). Lo anterior guarda estrecha concordancia con la exposición crónica a altos niveles de metales pesados. En C. undatella se observó una infiltración anormal de células eosinófilas dentro del tejido gonadal, lo cual se discute más adelante.
Los resultados mostraron que C. undatella presenta actividad reproductiva (madurez y desove) durante la mayoría de los meses con una producción continua de gametos (evidenciada por la presencia de gametos en diferentes fases de desarrollo al mismo tiempo) y desoves parciales. En el caso de los machos, es más marcada la actividad reproductiva continua (madurez o espermiación durante todo el año). De la misma forma, se ha reportado una actividad reproductiva continua en dos poblaciones de la misma especie en Bahía Falsa y El Cardonal, ambos lugares dentro de la bahía de La Paz (Baqueiro-Cardenas & Masso-Rojas, 1988), lo cual indica que es parte de la estrategia reproductiva de la especie.
Sin embargo, se detectan diferencias temporales en los periodos de mayor intensidad de desove entre las poblaciones. En este estudio, el mayor porcentaje de hembras en desove se presenta en julio (50%) y septiembre (100%), mientras que en Bahía Falsa es en abril (30%), junio (45%), enero y febrero (máximo de 37%), y finalmente, en El Cardonal su máximo se presenta en octubre (45%). En bivalvos se ha demostrado que la temperatura y disponibilidad de alimento son los factores más importantes que afectan su ciclo reproductivo (Cardoso et al., 2007; Freites et al., 2010) y, por lo tanto, las diferencias en la temporalidad de los desoves pueden ser atribuidas a las distintas condiciones ambientales que presenta cada zona (Laudien et al., 2001; Rodríguez-Jaramillo et al., 2001) o a variaciones interanuales en las condiciones ambientales. Es común que el desove en los bivalvos se lleve a cabo en concordancia con temperaturas cálidas y se dispare cuando excede de un nivel crítico característico (Maeda-Martínez, 2001). En C. fluctifraga (Sowerby, 1853) se detectó el desove principal durante julio y agosto, cuando la temperatura alcanzó sus máximos valores (29 y 30 °C, respectivamente) (Castillo-Durán et al., 2013); mientras que en C. californiensis (Broderip, 1835) el desove se presentó cuando la temperatura fue mayor a 24 °C (García-Domínguez et al. 1993). En contraste, también se ha reportado una reproducción continua independientemente de la variación de la temperatura para C. cancellata (Linnaeus, 1767) en Florida (Moore & López, 1969).
De acuerdo con este estudio y con lo reportado por Baqueiro & Masso (1988), en Baja California Sur C. undatella se reproduce continuamente sin un efecto aparente originado de la variación estacional de la temperatura del agua o de la concentración de clorofila-a. Sin embargo, en Santa Rosalía encontramos que los periodos máximos de desove y espermiación coinciden con los máximos valores en la temperatura superficial del mar. Por su parte, las mayores concentraciones de clorofila a podrían estar más relacionadas con los periodos de maduración gonádica y desove o con una mayor cantidad de alimento para la producción de gametos o para las larvas, respectivamente.
En C. undatella se encontraron células eosinófilas infiltradas en el tejido gonadal similares a las células hemocíticas relacionadas con la excreción de material extraño en Cerastoderma edule (Linnaeus, 1758) (Russell-Pinto et al., 1994). En moluscos, los hemocitos (células inmunoefectoras) y células cafés son de los principales mecanismos de defensa involucrados en el proceso de inflamación y en la acumulación y desintoxicación de contaminantes (Ottaviani et al., 2010; Ottaviani, 2011; Boscolo-Papo et al., 2014).
Cuando consideramos que el tejido gonadal de C. undatella muestra una infiltración de células eosinófilas con una prevalencia alta (intermedia y abundante) en todos los meses y fases de desarrollo gonádico, podemos descartar dos causas: presencia de alguna infección por parásitos (no se encontró evidencia de su presencia) o que se deba al proceso de recuperación natural de la gónada (se presentó en el desarrollo y la madurez con > 80% de prevalencia). Por lo tanto, se podría hipotetizar que, debido a que estos organismos están expuestos crónicamente a un ambiente contaminado por metales (Shumilin et al., 2013), estas células están relacionadas con la reacción celular a estrés abiótico, el cual resulta en un proceso de inflamación inmune innato (Bouallegui et al., 2017).
Distintos autores mencionan que después del transporte de hemocitos y células cafés, algunos tejidos se identifican como blancos de acumulación o desintoxicación de contaminantes en moluscos (Boscolo-Papo et al., 2014). Estas infiltraciones se presentan acompañadas de alteraciones morfológicas a nivel celular, como un decremento en el volumen de una célula o un órgano, así como una pérdida de grosor del epitelio o atrofia (Bouallegui et al., 2017). Esta pérdida de la arquitectura normal del tejido gonadal en C. undatella fue evidente con la reducción y desaparición del tejido conectivo. Dicho fenómeno ha sido descrito previamente por Villalba et al. (2001).
A pesar de que C. undatella presenta actividad reproductiva todo el año, la alta prevalencia de infiltraciones hemocíticas y la pérdida de la arquitectura normal del tejido gonadal sugieren una potencial deficiencia en su reproducción. Es importante hacer más estudios (gasto energético, fecundidad y viabilidad de los huevos) para confirmarlo y para determinar su relación con la contaminación presente en la zona.

