











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los encinos, también conocidos como robles, pertenecen al género Quercus y se caracterizan por estar presentes en las zonas montañosas del territorio mexicano (Martínez et al., 2022), en los bosques templados y matorrales. Estas especies son clave para el funcionamiento de los ecosistemas (Valencia, 2004), además, desempeñan un papel fundamental en la economía de las comunidades locales (Corrado et al., 2024). Estas especies tienen un rango ecológico amplio y una morfología diversa, extendiéndose prácticamente por todo el territorio nacional, con la excepción del estado de Yucatán (Encina y Villarreal, 2002).
En México, los bosques de encinos han sufrido degradación intensiva debido a diversas actividades antropogénicas, como el cambio de uso del suelo y la extracción de madera en forma de leña y carbón, realizadas para satisfacer necesidades básicas de la población local (Corrado et al., 2024). Por ejemplo, la madera de los robles ha sido el segundo recurso más utilizado a nivel nacional (Luna et al., 2003), sólo después de la madera de pinos (Moctezuma y Flores, 2020). También se ha reportado que el efecto del cambio climático global podría afectar la distribución espacial de los robles (Mirhashemi et al., 2023), generando un desequilibrio en los ecosistemas que ocupan, lo cual afecta la biodiversidad que depende de esta vegetación para su funcionamiento (Vaca et al., 2024). Por otro lado, existe una escasez de información sobre la distribución geográfica y ecológica de las especies locales, lo que dificulta el desarrollo de estrategias efectivas de conservación.
Durante los últimos años, los modelos de distribución de especies (MDE) han adquirido un papel fundamental en la ecología predictiva, permitiendo comprender la distribución potencial geográfica y ecológica pasada, presente y futura de los organismos en este planeta (Maciel et al., 2015; Marsh et al., 2023; Belfiore et al., 2024; Wei et al., 2024). Estos modelos se elaboran mediante la aplicación de técnicas avanzadas de programación y el uso de sistemas de información geográfica (SIG) (Drake et al., 2006). Para su desarrollo, se necesitan dos tipos de datos: los primarios, que incluyen la ubicación geográfica de las observaciones o recolecciones conocidas de la especie de interés, y los secundarios, que comprenden información ambiental como el clima, la topografía y la vegetación (Menon et al., 2010). Con esta información, se modela la idoneidad geográfica y ambiental de una especie utilizando diversas herramientas, entre las cuales se destaca MaxEnt, reconocida por tener capacidad de predicción y por ser ampliamente utilizada en este campo (Peterson y Robins, 2003).
En la cuenca de Cuitzeo, los encinos han sido muy explotados para elaborar leña, carbón y madera para escuadría, como lo han señalado Aguilar et al. (2012). Sin embargo, el cambio de uso del suelo ha provocado la fragmentación de su hábitat y la disminución de las poblaciones (López et al., 2006), lo que supone una alteración en su estructura y composición. Estas modificaciones han causado una intensa degradación de los bosques y matorrales donde habitan, lo que podría tener un impacto negativo en especies amenazadas o en peligro. A nivel local, se ha prestado escasa atención al estudio de los encinos desde perspectivas ecológicas y geográficas.
Este estudio tuvo por objetivo determinar las áreas geográficas y las condiciones ambientales de encinos en la cuenca de Cuitzeo, utilizando modelos de distribución potencial. Las especies que se estudiaron se encuentran en la categoría de amenazadas (A) y en peligro de extinción (P). De acuerdo con la NOM-059-SEMARNAT-2010 (SEMARNAT, 2010), la madera de Quercus crassipes (A) es explotada por la población local, mientras que la distribución de Quercus frutex (P) y Quercus glaucoides (A) es restringida en la región. Los hallazgos de esta investigación permitirían ubicar poblaciones de encinos y señalar áreas potenciales para su manejo y conservación en la zona de estudio.
Materiales y métodos
Área de estudio
La cuenca del lago de Cuitzeo se localiza al norte del estado de Michoacán y al Sur de Guanajuato, entre los 19° 30’ y 20° 05’ de latitud norte y los 100° 30’ y 101° 35’ de longitud oeste, dentro de la región fisiográfica conocida como Sistema Volcánico Transversal, y tiene una superficie aproximada de 4000 km2 (Figura 1). En la cuenca predomina un clima templado con lluvias en verano; la temperatura media anual es de 17 °C, con una precipitación media anual de 800 mm y una altitud aproximada de 1830 m. s. n. m. (Morales et al., 2010). Esta área geográfica es de gran importancia ecológica para la región debido a su cobertura natural, compuesta principalmente por matorrales y bosques (López et al., 2006; Mendoza et al., 2011). En ella se alberga el lago de Cuitzeo, el segundo más grande de México, con una superficie de aproximadamente 300 km², el cual desempeña un papel crucial en la regulación del clima local y regional; además, es vital para el sustento de numerosas especies de flora y fauna (Morales et al., 2010).

Distribución Geográfica
Localidades de presencia de las especies
Uno de los insumos más importantes que se requieren para la construcción de los modelos de distribución, son las localidades de presencia (Liu et al., 2024), las cuales se obtuvieron de la revisión de las siguientes colecciones científicas: Herbario IEB del Instituto de Ecología, A. C., México, y Herbario de la Facultad de Biología de la UMSNH (EBUM). También se consultaron las bases de datos de colecciones en línea de la Global Biodiversity Information Facility (GBIF) y la Red Mundial de Información sobre Biodiversidad (REMIB) (Peterson y Nakazawa, 2008). Posteriormente, se llevó a cabo una depuración de los datos para evitar localidades repetidas, mediante el uso de la herramienta Niche Toolbox, donde se eliminaron las coordenadas duplicadas y aquellas que estaban separadas por una distancia menor a 1 km entre cada registro (Monterrubio et al., 2016). Como resultado, se obtuvo una única coordenada para cada celda de 1 km². Con este procedimiento se evitó la sobreestimación de los modelos debido a los efectos de la autocorrelación espacial (Manzanilla et al., 2019).
Se identificaron 40 localidades para Quercus crassipes Humb. & Bonpl, una especie que se encuentra en estado de conservación amenazada, su hábitat ha sido afectado por la tala inmoderada para el establecimiento de cultivos (Arizaga et al., 2009). Para Q. frutex Trel, se registraron 10 localidades; esta especie se encuentra en peligro de extinción debido a su distribución restringida y a la transformación de su hábitat causada por incendios y la expansión de cultivos (Arizaga et al., 2009); se identificaron nueve localidades para Q. glaucoides M. Martens & Galeottiy, una especie amenazada (SEMARNAT, 2010), cuyo hábitat ha sido transformado en asentamientos humanos (Arizaga et al. (2009).
Variables bioclimáticas y topográficas
Las variables climáticas utilizadas fueron extraídas de la base de datos de WORDCLIM versión 2.0 (disponible en http://www.worldclim.org). Comprende un total de 19 variables bioclimáticas con una resolución espacial de 1 km². Con el fin de prevenir la multicolinealidad y evitar el sobreajuste de los modelos (Loiselle, 2008; Manzanilla et al., 2019), se realizó un análisis de correlación utilizando el paquete estadístico de Statistica 7.0. Este análisis permitió seleccionar un subconjunto de variables climáticas con correlaciones menores entre sí (r < 0.5), resultando en la elección de nueve variables relevantes (Tabla 1) (Peterson y Nakazawa, 2008). Dado que la cuenca de Cuitzeo tiene una superficie de aproximadamente 4000 km², en términos hidrológicos es una unidad natural pequeña, siendo estas variables de baja resolución (1 km²). Por esta razón, se ajustó la resolución espacial a 6” de arco (~200 m x 200 m), siguiendo enfoques metodológicos implementados por (Hutchinson, 1998) y (Fick y Hijmans, 2017). Este ajuste permitió reflejar una mayor precisión en las características ambientales de la cuenca, tales como los gradientes altitudinales y climáticos, mejorando así la precisión de los modelos de distribución de las especies. Asimismo, se utilizó un modelo digital de elevación, también ajustado a una resolución de 200 metros, para asegurar la coherencia en la representación espacial de las variables (Téllez et al., 2010).
Tabla 1 Variables climáticas utilizadas.
| Número | Variables |
| 1 | Oscilación diurna de la temperatura media mensual (tº máx.-tº min.) (BIO2) |
| 2 | Isotermalidad (B2/B7) (*100) (BIO3) |
| 3 | Estacionalidad de la temperatura (desviación estándar *100) (BIO4) |
| 4 | Rango de Temperatura Anual (B5 - B6) |
| 5 | Precipitación anual (mm) (BIO12) |
| 6 | Precipitación del mes más húmedo (BIO13) |
| 7 | Precipitación del trimestre más húmedo (BIO16) |
| 8 | Precipitación del trimestre más cálido (mm) (BIO18) |
| 9 | Precipitación del trimestre más frío (mm) (BIO19) |
Nota. se muestran las nueve variables que se utilizaron en el desarrollo de los modelos de distribución potencial.
Fuente: Elaboración propia a partir de los datos de Worldclim versión 2.0.
Modelado de la distribución de las especies de encino
Para generar los modelos de distribución potencial de cada especie, se delimitó el área de modelamiento en el espacio geográfico mediante un recorte a nivel de cuenca. Este enfoque permitió ajustar las variables a un mismo tamaño, evitando así la sobreestimación de las áreas de distribución (Monterrubio et al., 2016; Pan et al., 2023). El procedimiento se realizó utilizando el software MaxEnt versión 3.3 (Phillips y Dudík, 2008). MaxEnt estima la distribución potencial mediante el principio de máxima entropía, es decir, calcula la probabilidad de áreas aptas o idóneas de una especie a partir de datos de presencia conocida de las especies. Este método divide la información en dos conjuntos: al azar con el 50% de prueba y el 50% para validar los modelos. Además, utiliza un conjunto de variables ambientales anteriormente mencionadas (Phillips et al., 2006). Bajo esta premisa, el valor de cada característica analizada en la distribución estimada debería coincidir con su valor promedio empírico.
Los modelos generados por MaxEnt producen valores en un rango de 0 a 1, donde un valor alto indica condiciones muy favorables para la presencia de la especie, mientras que un valor bajo sugiere lo contrario (Peterson y Nakazawa, 2008). Los ajustes utilizados en MaxEnt fueron los siguientes: Se desactivó Extrapolate y Do clamping para evitar el sobreajuste de los modelos, y el tipo de archivo de salida logística fue de 500 iteraciones y un umbral de convergencia 0.00001 (Manzanilla et al., 2019).
Validación estadística de los modelos
El desempeño de los modelos fue evaluado a través de las métricas obtenidas con la curva ROC (Receiver Operating Characteristic), la cual describe la tasa de identificación correcta de presencias (sensitivity) en comparación con la tasa de falsas presencias (specificity). Sin embargo, se ha reportado que este método no es adecuado para evaluar modelos de nicho ecológico (Lobo y Hortal, 2003). En esta investigación, se optó por utilizar la Partial ROC, una modificación sugerida por Peterson et al. (2008). Para el análisis se utilizó un umbral predefinido de E = 5%, lo que permite establecer la sensibilidad mínima aceptable y evaluar el rendimiento del modelo por encima de este umbral. Se llevaron a cabo 1000 iteraciones para calcular una versión parcial del Área Bajo la Curva (AUC) con valores que oscilan de 0 a 1 (valores entre 0.7 y 0.9 indican que el rendimiento de los modelos es bueno, y valores mayores a 0.9 indican un rendimiento excelente y un umbral de omisión de 95%) (Peterson, 2011; Peterson et al., 2008; Wu et al., 2024; Yang et al., 2024). Los resultados se expresaron como la razón entre la curva ROC observada y la esperada al azar, con una gama de puntuaciones entre 1 y 2. Valores inferiores a 1 indican un modelo obtenido completamente al azar, mientras que valores cercanos a 2 indican una correlación significativa y no aleatoria. Además, se realizó una prueba de hipótesis Z con los valores de las proporciones AUC, el cual complementó la fiabilidad de este análisis (Manzanilla et al., 2023). Este análisis se realizó utilizando el software Tool Partial ROC versión 1.0 (Peterson, 2011).
Análisis de la distribución en el espacio geográfico y ecológico
En esta investigación, los modelos de distribución se ajustaron a un mapa de vegetación y uso del suelo Serie V (continuo nacional), del Instituto Nacional de Estadística y Geografía (INEGI, 2013), a una escala 1:250 000. La elección de esta versión se basó en la necesidad de mantener la coherencia temporal con los datos de presencia seleccionados de cada una de las especies, que corresponden a años anteriores al 2015, en su mayoría. Posteriormente, se determinó el área de aptitud potencial en kilómetros cuadrados. Además, se delimitaron las áreas de aptitud ambiental, donde se superpusieron los modelos de distribución potencial obtenidos de MaxEnt y las variables de temperatura y precipitación media anual. Este análisis permitió identificar el rango de ambas variables para cada una de las especies estudiadas. Adicionalmente, se generó un mapa de orientación de laderas en grados a partir del modelo digital de elevación, que permitió identificar si las especies presentan alguna preferencia por ciertas laderas. La evaluación se realizó mediante un análisis de estadística circular utilizando el software Oriana (Kovach, 2004). Este análisis incluyó la prueba de uniformidad de Rayleigh para determinar si existe una orientación preferente hacia algún cuadrante (norte, sur, este u oeste). El vector medio (μ) indicó la dirección preferida, el parámetro de concentración k reveló la tendencia direccional de los datos y el valor de Z proporcionó información sobre la desviación de los datos con respecto a la homogeneidad. Estas condiciones ambientales reflejan tanto el nicho fundamental (NF), definido como las áreas potencialmente aptas para la especie, como el nicho realizado (NR), que representa las áreas donde la especie realmente habita, más acorde con la realidad (Soberón y Miller, 2009).
Resultados
El análisis estadístico realizado sobre los tres modelos desarrollados con MaxEnt reveló que los valores de AUC para el conjunto de entrenamiento se situaron en un rango comprendido entre 0.85 y 0.95, mientras que para el conjunto de validación los valores oscilaron entre 0.80 y 0.96. Además, los valores obtenidos mediante el análisis de Partial ROC varían entre 1.11 y 1.40, respaldados por un valor de Z estadísticamente significativo (p < 0.001), como se muestra en la Tabla 2. Estos valores son superiores a 1, lo cual confirma que los modelos tienen un rendimiento mejor que el azar y proporcionan una medida adicional de la precisión predictiva.
Tabla 2 Valores estadísticos de AUC y del análisis parcial de la curva ROC.
| Especie | Valor de AUC | Valor de Roc Partial | Z | P | |
| Entrenamiento | Validación | ||||
| Q. crassipes | 0.95 | 0.96 | 1.20 | 88.54 | <0.0001 |
| Q. frutex | 0.90 | 0.93 | 1.40 | 142.30 | <0.0001 |
| Q. glaucoides | 0.85 | 0.80 | 1.11 | 183.00 | <0.0001 |
Nota. Los valores presentados son la media de la razón del AUC que oscila entre 0 y 1, obtenido de las 1000 iteraciones que se realizaron en el análisis con MaxEnt y los valores de partial ROC con valores <1 = azar y 2 = prueba perfecta.
Fuente: Elaboración propia.
Distribución geográfica potencial
Los modelos de distribución geográfica potencial de las tres especies de encinos se ubican principalmente al sur de la cuenca de Cuitzeo, una región donde predominan los bosques de pino y encino-pino (Figura 2). Para Q. crassipes, el modelo identificó aproximadamente 2285 km² de áreas con aptitud para su distribución geográfica. Estas áreas se encuentran dispersas al sur, abarcando Cerro de la Nieve, Cerro Burro y Cerro del Frijol, donde predominan los bosques de Abies, pino y matorral subtropical. Hacia el suroeste se encuentran en Cerro del Quinceo, Las Joyitas y Cuto de la Esperanza, en bosques de pino-encino. Al sureste se extienden por la cañada del Río Chiquito, San Miguel del Monte y Jesús del Monte, en áreas de bosques de pino-encino. Finalmente, al norte, abarcan Cerro de las Cruces, Cerro las Ventanas, La Joya y Laguna Verde, donde predomina el matorral subtropical.
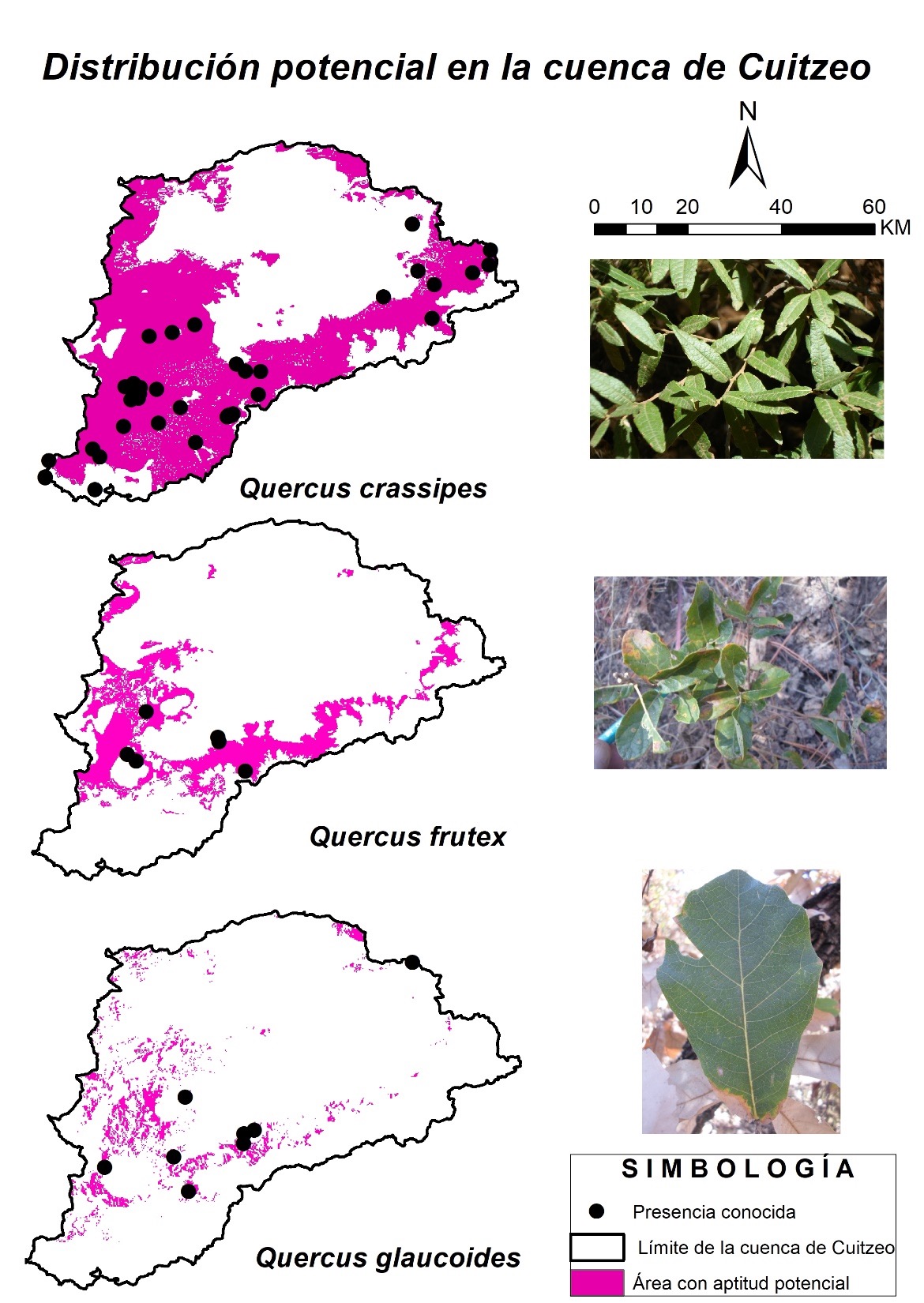
Fuente: Elaboración propia.
Figura 2 Distribución potencial para las tres especies de encinos en la cuenca de Cuitzeo.
Las especies con un número limitado de localidades de presencia muestran una distribución más restringida del área potencialmente apta para su distribución. De acuerdo con los modelos Q. frutex, tiene aproximadamente 742.44 km² de área con aptitud, concentradas al suroeste de la cuenca. Estas áreas incluyen las partes bajas del Cerro del Águila y el Cerro del Quinceo, así como Cuto de la Esperanza y Las Flores, donde se encuentran bosques de encino y zonas agrícolas. También se observan al sureste, en San Miguel del Monte, Jesús del Monte, la cañada del Río Chiquito, San José de las Torres y La Resinera. En esta zona predomina el bosque de pino y encino, mientras que al norte se localizan en el Cerro de las Cruces, en áreas de matorral subtropical.
Para Q. glaucoides, el modelo predijo áreas con una aptitud de 254.0 km², siendo la especie con la menor área predicha para su presencia en la zona de estudio. Estas áreas potenciales se encuentran al suroeste de la cuenca, en las zonas más bajas del Cerro del Águila, ascendiendo por Iratzio, Las Pilas, Cuto de la Esperanza, Cerro del Quinceo, Buenavista y Arroyo Colorado, hasta San José de las Torres, donde se distribuyen bosques de encino, matorral subtropical y áreas agrícolas. Además, al norte, se localizan en las partes bajas del Cerro de las Cruces, en áreas de matorral subtropical.
Perfil bioclimático
El perfil bioclimático obtenido para las tres especies de encinos en la cuenca muestra que Q. crassipes presenta un rango de temperatura que va de 12 °C a 18 °C, con una media de 15 °C ± 1.1 °C y una precipitación anual que varía entre 763 mm y 1408 mm, con una media de 1408 mm ± 168 mm. Por su parte, Q. frutex tiene un rango de temperatura entre 13 °C y 17 °C, una media de 15 °C ± 0.8 °C y una precipitación que va de 819 mm a 1325 mm, con una media de 1059 mm ± 112 mm. Finalmente, Q. glaucoides presenta un rango de temperatura de 14 °C a 17 °C, una media de 15 °C ± 0.9 °C y el rango de precipitación que oscila entre 800 mm a 1255 mm, con una media de 989 mm ± 100 mm. Estos datos reflejan la similitud en las condiciones climáticas por estas especies, pero muestran una ligera diferenciación en los rangos de precipitación que caracterizan sus respectivos hábitats en la zona de estudio.
Perfil topográfico
Rango altitudinal
La cuenca de Cuitzeo presenta una altimetría que varía aproximadamente entre los 1800 m y los 4220 m. Se encontró que para Q. crassipes su rango altitudinal va desde los 1865 m hasta los 2810 m, con una altitud media de 2244 m ± 138 m. Por otro lado, Q. frutex oscila entre los 1933 m y los 2632 m, tiene una altitud media de 2239 m ± 102 m. En cuanto a la distribución de Q. glaucoides, se registró entre los 1885 m hasta los 2484 m y una media de 2178 m ± 101 m.
Pendiente
Se obtuvo la inclinación del terreno en la cuenca de Cuitzeo, la cual presenta una variedad de pendientes, que van desde 0° hasta 38°, con una media de 10° ± 6.8°. Los encinos estudiados muestran una distribución que abarca desde pendientes suaves hasta pronunciadas. Q. crassipes se encontró en pendientes que oscilan entre 0° y 32°, una media de 7° ± 4.8°. Para Q. frutex, su distribución abarca pendientes de 0° a 24° y una media que va de 9° ± 5.6°, mientras que para Q. glaucoides el rango de pendientes va desde 0° hasta 21°, con una media de 6° ± 4.6°.
Orientación de laderas
El análisis de orientación de laderas mostró para Q. crassipes un vector medio de µ = 323.62° ± 3.4°, con un valor de Z = 11.2 (p = 0.00001), indicando una clara preferencia por la orientación hacia la porción norte de la cuenca de Cuitzeo. En el caso de Q. frutex, se registró un vector medio de µ = 309.98° ± 2.0°, con un valor de Z = 29.0 (p = 0.0001), lo que señala una orientación preferencial en la porción noroeste de la cuenca. Por su parte, Q. glaucoides presentó un vector medio de µ = 255.67 ± 10.3, con un valor de Z = 11.2 (p = 0.0001), correspondiendo a una orientación preferencial hacia el sur de la zona de estudio (Figura 3).
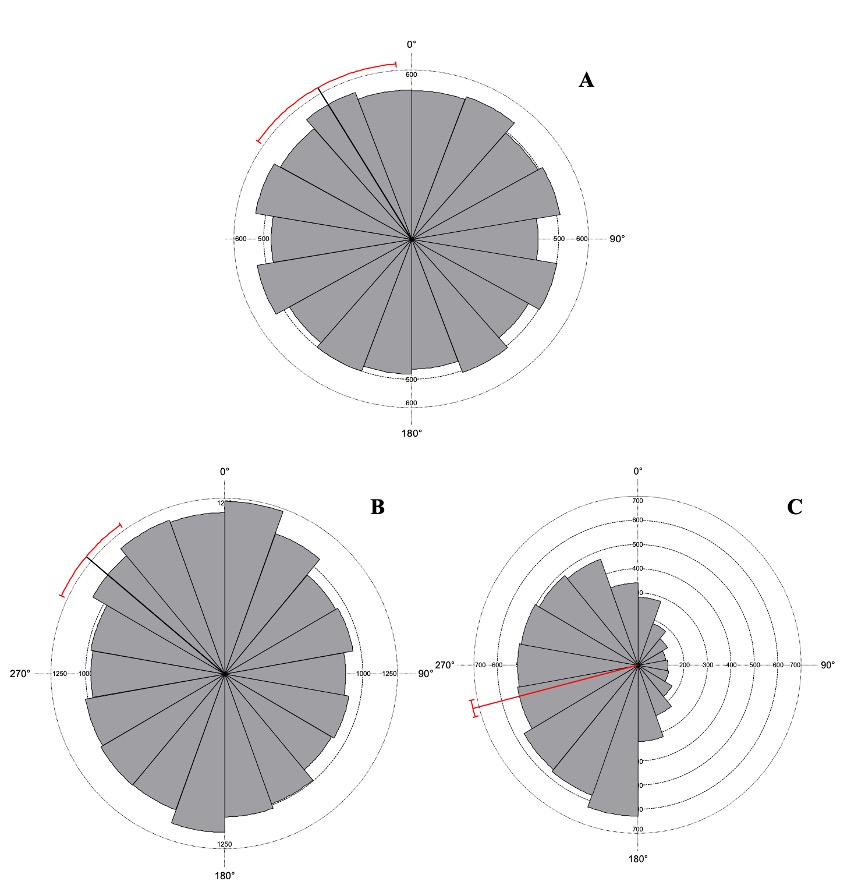
Fuente: Elaboración propia a parir de la intersección de los modelos de distribución y el mapa de orientación de laderas.
Figura 3 Orientación de ladera (a) Quercus crassipes, (b) Quercus frutex (c) Quercus glaucoides en la cuenca de Cuitzeo, donde el valor medio es la línea punteada que parte del centro del diagrama hacia la periferia y la línea de color rojo indica el límite de confianza al 95%.
Discusión
La representación espacial obtenida mediante los modelos con MaxEnt para cada especie de estudio han mostrado un ajuste adecuado entre los conjuntos de datos de entrenamiento y prueba, como lo confirman los resultados de la prueba de Partial ROC, donde se siguieron las recomendaciones de Peterson et al. (2008). Además, los valores de AUC obtenidos oscilan entre 0.85 y 0.95, lo cual indica que los modelos poseen una alta capacidad de discriminación, que sustenta la confiabilidad de las predicciones registradas. Este enfoque ha sido ampliamente respaldado por el éxito que ha mostrado en diversas investigaciones, debido a la aproximación sólida del nicho ecológico y geográfico, tal como se ha reportado en otros estudios (Manzanilla et al., 2023; Wu et al., 2024; Yang et al., 2024). Asimismo, al considerar únicamente áreas donde los modelos predicen presencia, se facilita la identificación de zonas con alta probabilidad de idoneidad, lo que puede optimizar el monitoreo, manejo y planificación para la conservación de estas especies en la cuenca. Sin embargo, es importante considerar que la implementación de este enfoque presenta limitaciones, como la posible subestimación del rango ecológico completo de la especie y la sobreestimación de la idoneidad del hábitat en ciertas áreas (Campos et al., 2023; Kaur et al., 2024). Para mejorar la precisión y aplicabilidad de estos modelos, se sugiere incluir datos de ausencia, incorporar el conocimiento experto sobre la ecología y comportamiento de la especie, y realizar trabajo de campo para validar la información obtenida.
Análisis en el ámbito geográfico y ecológico
Los requerimientos de precipitación media anual son más altos para Q. crassipes (1408 mm), lo que indica que requiere mayor humedad y coincide con su rango geográfico, debido a que esta especie tiende a preferir altitudes más elevadas (Arizaga et al., 2009), aunque es posible encontrarlos en terrenos planos, tal como se observó en el rango de pendientes que van de planas a moderadas. También presenta una coincidencia con el tipo de vegetación, pues principalmente se encuentran en zonas boscosas y en terrenos agrícolas de la cuenca, mientras que la preferencia por las laderas con orientación al norte estaría indicando que recibe menos luz solar directa durante gran parte del día. Posiblemente, estas condiciones hacen que esta especie tenga una mayor área de distribución. Por lo tanto, Q. frutex y Q. glaucoides, ambas tienen una distribución geográfica restringida (Arizaga et al., 2009), con una precipitación media de 1059 mm y 989 mm, donde se establecen los bosques de encino y matorrales, coincidiendo con el rango de pendientes moderadas y una preferencia en la orientación de laderas al noroeste y sur, es decir, reciben mayor exposición de la luz solar durante el día. La distribución limitada que presentan podría indicar una mayor especialización para adaptarse a las condiciones específicas de su hábitat, como la disponibilidad de luz solar, la competencia con otras especies vegetales y el acceso al agua, entre otros, lo cual podría ser objeto de futuras investigaciones. Además, en esta zona de la cuenca se han venido registrando desde hace algunos años una mayor degradación de la vegetación debido al cambio de uso de suelo, impulsado por actividades antrópicas como la agricultura y la ganadería (López et al., 2006). Este factor podría reducir las áreas aún más donde estas especies pueden crecer y prosperar, limitando su distribución.
Es relevante señalar que no se han realizado estudios previos sobre la distribución ecológica y geográfica de dichas especies en la cuenca de Cuitzeo que permitan contrastar los hallazgos de esta investigación. No obstante, Arizaga et al. (2009), en el manual de biodiversidad de encinos michoacanos, detallan los requisitos ecológicos de estas especies, los cuales coinciden con los resultados obtenidos. Es por ello que la obtención de esta información representa la primera aproximación sobre las condiciones ambientales y geográficas bajo las cuales están creciendo estas especies a nivel local, lo que es fundamental para el monitoreo y la identificación de áreas críticas de Q. frutex y Q. glaucoides dada su distribución restringida. Además, esta información puede ser una herramienta valiosa para evaluar posibles escenarios futuros de cambio climático, tal como se ha reportado en otras regiones con especies que habitan en entornos similares a los encinos (Manzanilla et al., 2019).
Conclusiones
Los modelos predicen que las áreas de distribución geográfica potencial de las tres especies de encinos se concentran principalmente al sur de la cuenca, en regiones caracterizadas por la presencia de bosques de pino y encino-pino. La especie Q. crassipes muestra la mayor extensión de áreas con aptitud para su presencia, mientras que Q. frutex y Q. glaucoides se registraron como áreas de distribuciones más restringidas.
Los perfiles bioclimáticos y topográficos indican que las tres especies de encinos comparten un rango de temperatura similar, pero presentan ligeras diferencias en sus preferencias de precipitación y altitud. Q. crassipes se encuentra en altitudes y pendientes más variadas, mientras que Q. frutex y Q. glaucoides tienen distribuciones más restringidas en cuanto a estas variables. Además, cada especie muestra una preferencia clara por la orientación de laderas, lo que sugiere que la topografía juega un papel importante en la determinación de su distribución geográfica.
Este aporte es importante para desarrollar acciones de conservación, considerando las zonas específicas y las condiciones ambientales que los modelos predicen para cada una de estas especies, y con ello priorizar la protección y manejo sostenible en la zona de estudio.
Conflicto de interés
Los autores declaran que no existen conflictos de interés.

