











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Pinus patula Schiede ex Schltdl. et Cham. tiene amplia distribución en la sierra Madre Oriental y el este del Eje Neovolcánico Transversal de México, es de gran importancia económica debido a su rápido crecimiento y buena calidad de madera (Velázquez et al., 2004). Es la especie más aprovechada del género Pinus en México (Flores y Moctezuma-López, 2021), por lo que desde hace tiempo se iniciaron programas de mejora en países del hemisferio sur, para su uso extensivo en plantaciones forestales comerciales (Velázquez et al., 2004). En los programas de mejoramiento genético se establecen ensayos de progenie para evaluar el control genético (heredabilidad) de los progenitores sobre las características de interés en sus progenies (White et al., 2007); además, debe evaluarse el mayor número posible de procedencias y familias para realizar la selección adecuada de los orígenes y genotipos superiores con base en una característica deseada (Vallejo y Cob, 2021).
Al establecer plantaciones forestales comerciales se reduce la presión sobre los bosques naturales, aumenta el abasto de materias primas y se favorece la reconversión de tierras agrícolas poco productivas a terrenos con vegetación forestal (Ramírez y Simonetti, 2011). En ese sentido, es deseable evaluar el rendimiento de las especies y sus procedencias con potencial para lugares diferentes a su sitio de origen, como en el caso de Pinus patula, para su uso en plantaciones comerciales en el estado de Durango, entidad que posee aproximadamente 1,000,000 de hectáreas con potencial para plantaciones forestales, pero cuyas especies nativas cuentan con turnos de corta que superan los 60 años (Vázquez-Cisneros et al., 2018), además de presentar supervivencia menor de 50 % al año de plantado (Burney et al., 2015). El alto potencial productivo de P. patula lo hace una opción atractiva de introducción, dado al éxito en su cultivo extensivo en países como Colombia, Ecuador y Sudáfrica (Gotore et al., 2014); sin embargo, uno de los problemas en la mejora genética de los árboles forestales es su longevidad, lo que extiende el tiempo necesario para evaluar los resultados de las progenies. Para acelerar este proceso se usa la selección temprana, consistente en seleccionar genotipos con base en un rasgo que se manifieste a edades jóvenes para mejorar una característica en edades posteriores, lo que puede reducir el ciclo de mejoramiento (Wu, 1999).
La eficacia de la selección temprana depende de la variabilidad entre procedencias y familias, así como de los parámetros genéticos de las características involucradas, como la heredabilidad y la correlación genética entre ellas; además, se espera que los genotipos expresen su potencial de forma diferenciada en ambientes contrastantes. Si la característica juvenil tiene mayor heredabilidad y está fuertemente correlacionada con la característica de interés, la selección temprana será más eficiente que la selección directa en edad adulta (White et al., 2007).
El objetivo del presente estudio fue evaluar la supervivencia y el control genético sobre altura y diámetro de planta en progenies de Pinus patula a los 8, 10 y 12 meses después de la plantación (edad) de un ensayo establecido en Durango, México, además de determinar la correlación entre las características evaluadas y variables geográficas y ambientales de las procedencias. Las hipótesis fueron: i) la procedencia-progenie influye en la supervivencia, altura y diámetro de P. patula establecido en campo en Durango, México, ii) existe control genético sobre la altura y diámetro de las progenies en el ensayo de campo evaluado y, iii) existe correlación alta y significativa entre las características (supervivencia, altura y diámetro) con al menos una variable geográfica o ambiental de las procedencias.
Materiales y métodos
Características del área de estudio
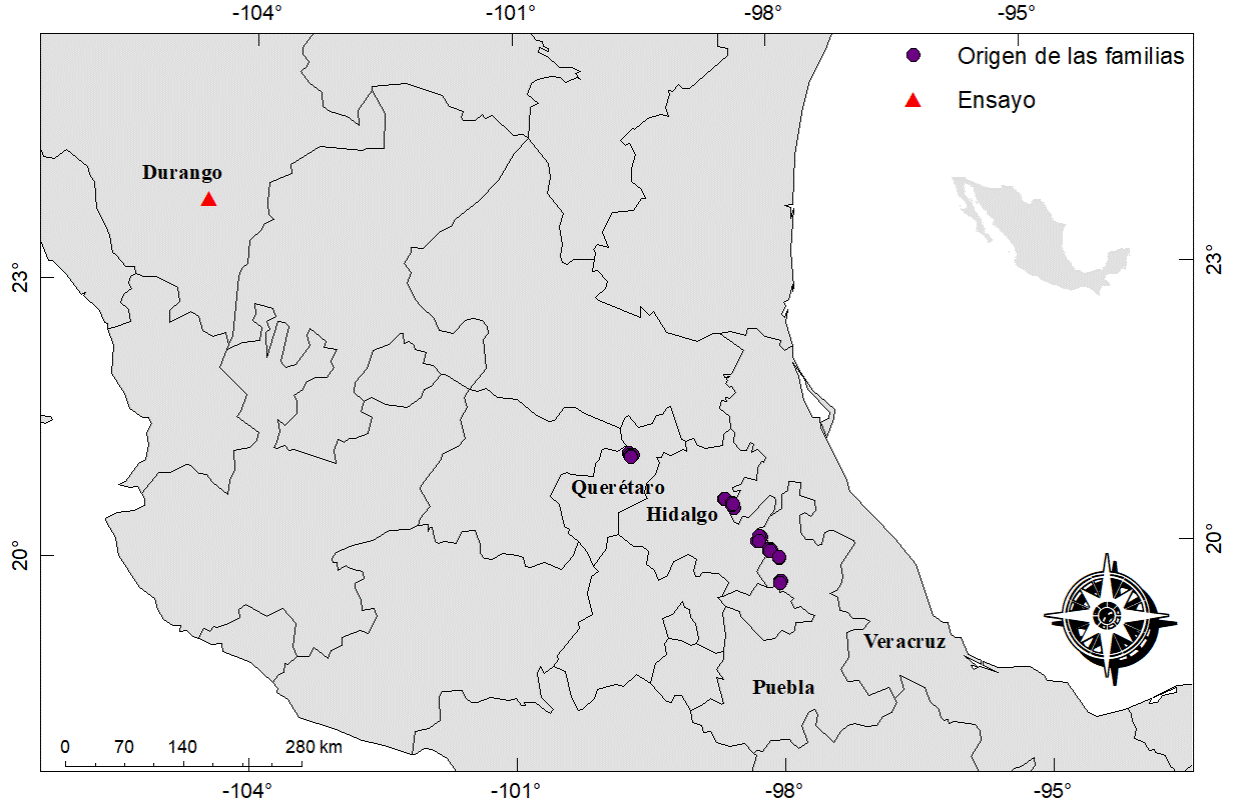
La semilla se recolectó de 42 árboles superiores ubicados en rodales naturales de siete procedencias distribuidas en los estados de Puebla (Ahuazotepec, Zacatlán y Aquixtla - cinco, cuatro y nueve árboles, respectivamente), Querétaro (Pinal de Amoles - siete árboles), Hidalgo (Zacualtipán y Acaxochitlán - cuatro y seis árboles, respectivamente) y Veracruz (Huayacocotla - siete árboles) (Figura 1). La planta se produjo en el vivero de la Facultad de Ciencias Forestales y Ambientales de la Universidad Juárez del Estado de Durango, ubicada a 24° 00’ 48.38’’ N y 104° 41’ 03.64’’ O, a 1860 m de altitud.
Establecimiento, diseño experimental y manejo del ensayo
La siembra se realizó el 18 de febrero de 2022 en charolas de 98 tubetes individuales de 170 mL. Como sustrato se utilizó una mezcla de turba de musgo (40 %), corteza de pino compostada (30 %) y aserrín de pino crudo (30 %), más 10 g L-1 de fertilizante de lenta liberación (ocho meses) MulticoteTM 18-6-12 (N-P-K) más micronutrientes. La planta se mantuvo en forma secuenciada dos meses bajo condiciones de invernadero, tres meses bajo malla sombra de 60 % de retención de luz y un mes en intemperización.
A los seis meses de edad de las plantas (5 de septiembre de 2022; 15.8 cm de altura y 2.78 mm de diámetro al cuello de la raíz en promedio) se estableció el ensayo de procedencias- progenies en una parcela del ejido Aquiles Serdán, municipio de Durango, Durango, México, ubicada en las coordenadas 23° 53’ 39.24’’ N y 104° 33’ 43.94’’ O, a 1898 m de elevación. Se utilizó un espaciamiento de 2.5 × 2.5 m en un diseño experimental de bloques completos al azar con 15 repeticiones. En cada bloque se colocó una planta de cada familia de forma aleatorizada. Al fondo de cada cepa se agregaron 10 g de fertilizante de lenta liberación (ocho meses) MulticoteTM 18-6-12 (N-P-K) más micronutrientes. En el periodo de estiaje (marzo-agosto de 2023) se aplicaron riegos de auxilio cada 15 días (12 riegos en total con 5 L de agua por planta en cada riego). Por la presencia de hormiga arriera (Atta mexicana Fabricius y A. cephalotes L.) se aplicó mensualmente insecticida Lorsban® [clorpirifos etil (0,0-dietil 0-3,5,6-tricloro-2-piridil fosforotioato)], 0.5 kg sobre los hormigueros y 5 g alrededor de cada planta.
Variables climáticas registradas
La temperatura y grados-día mayor a 5 °C en el periodo de evaluación del ensayo (septiembre de 2022 a agosto de 2023) se obtuvieron con un termohigrómetro (Datalogger Elitech® modelo Rc-4hc, Elitech Technology, San Jose, California, EUA). La precipitación en el sitio del ensayo se midió con un pluviómetro rústico bajo la metodología descrita por Ponce-Rodríguez et al. (2021). La temperatura media anual en el sitio de plantación en el periodo de evaluación fue 19.3 °C. La temperatura máxima se registró en mayo de 2023 (40.6 °C) y la mínima en enero de 2024 (3.1 °C). La precipitación media anual fue 236 mm, con índice de sequía de 0.3. Las principales características físico-químicas del suelo fueron: 33.1 % de arena, 41.1 % de limo y 25.8 % de arcilla, textura franca, 1.0 % de materia orgánica, 6.9 de pH, 0.04 dS m-1 de conductividad eléctrica, 18.67 ppm de nitrógeno-amonio (N-NH4), 4.63 ppm de fósforo y 1.07 ppm de potasio.
Variables respuesta evaluadas
A los 8, 10 y 12 meses de edad del ensayo se evaluó la supervivencia, la altura y el diámetro al cuello de la raíz. La supervivencia se registró con variables dicotómicas: planta viva (1) o muerta (0). También se determinó la causa de mortalidad. La altura se midió con un flexómetro marca Truper® con aproximación en cm y el diámetro con un vernier digital marca SureBilt® con aproximación en mm.
Análisis estadístico y estimación de parámetros genéticos
Se realizaron los análisis de varianza con los procedimientos Mixed y GLM de SAS v9.4 (SAS Institute, 2022) con un nivel de significancia de P ≤ 0.05 de las variables respuesta a los 8, 10 y 12 meses de edad del ensayo. El modelo lineal utilizado fue:
Donde Yijk es el valor observado de la planta en la k-ésima familia, dentro de la j-ésima procedencia en el i-ésimo bloque, µ es la media poblacional, βi es el efecto fijo del i-ésimo bloque (i = 1, 2,…15), Pj es el efecto fijo de la j-ésima procedencia, BPij es el efecto fijo de la interacción bloque × procedencia, Fk(j) es el efecto aleatorio de la k-ésima familia anidada en procedencia (k = 1, 2,…42) ~NID (0,
Los componentes de varianza de familias (
Dónde:
Correlaciones genéticas y fenotípicas
Las correlaciones fenotípicas por familia se estimaron con el coeficiente de correlación de Pearson por medio del procedimiento Corr de SAS v9.4 (SAS Institute, 2022), mientras que las correlaciones genéticas fueron calculadas con la Ec. 5 (Falconer y Mackay, 2006). La covarianza de familias (Ec. 6) y el error estándar de las correlaciones genéticas (Ec. 7) se estimaron con las ecuaciones descritas por White y Hodge (1989) y Falconer y Mackay (2006), respectivamente.
Dónde: rg(x,y) es la correlación genética entre las variables "x" y "y", Covf(x,y) es la covarianza de familias entre “x” y “y”,
Correlaciones entre las variables respuesta con variables geográficas y climáticas del origen de las familias
Con el procedimiento Corr de SAS v9.4 (SAS Institute, 2022) se obtuvieron las correlaciones de Pearson entre las medias de supervivencia, altura y diámetro al cuello de la raíz a los 8, 10 y 12 meses de edad del ensayo con los valores de ubicación geográfica y variables bioclimáticas de las procedencias.
La temperatura media anual, la temperatura máxima y mínima registradas, la precipitación media anual y los grados-día mayores de 5 °C de los sitios de origen de las familias y del sitio de plantación se obtuvieron con el programa ClimateNA v5.3 (Wang et al., 2016). Las variables climáticas reportadas corresponden al periodo 1961 a 2022 (Cuadro 1), mismas que se usaron para obtener las correlaciones.
Cuadro 1 Valores de ubicación geográfica y variables climáticas de los sitios de procedencia.
| Procedencia | Lat (N) | Long (O) | Altitud (msnm) | TMA (ºC) | TMáx (ºC) | TMín (ºC) | PMA (mm) | GD5 | IS |
| Ahuazotepec | 20° 04.1’ | 98° 06.3’ | 2386 | 13.9 | 24.0 | 3.5 | 1361 | 3926 | 0.05 |
| Pinal de Amoles | 21° 07.3’ | 99° 39.8’ | 2641 | 13.7 | 24.5 | 2.5 | 962 | 3864 | 0.06 |
| Zacualtipán | 20° 37.6’ | 98° 36.2’ | 2076 | 13.4 | 23.4 | 4.2 | 1410 | 3722 | 0.04 |
| Acaxochitlán | 20° 12.0’ | 98° 12.9’ | 2209 | 14.4 | 24.4 | 4.6 | 1160 | 4084 | 0.06 |
| Zacatlán | 20° 02,2’ | 98° 03.1’ | 2338 | 13.9 | 23.9 | 3.4 | 1201 | 3956 | 0.05 |
| Aquixtla | 19° 46.4’ | 98° 00.8’ | 2756 | 12.3 | 24.3 | 0.8 | 777 | 3506 | 0.08 |
| Huayacocotla | 20° 34.1’ | 98° 30.6’ | 2003 | 13.9 | 24.0 | 4.2 | 1319 | 3892 | 0.05 |
| Ensayo | 23° 53.6’ | 104° 33.7’ | 1898 | 17.5 | 29.6 | 4.5 | 576 | 4574 | 0.11 |
Lat (N): latitud en el punto central de la procedencia, Long (O): longitud en el punto central de la procedencia, TMA: temperatura media anual, TMáx: temperatura máxima registrada, TMín: temperatura mínima registrada, PMA: precipitación media anual, GD5: grados-día mayor de 5 °C, IS: índice de sequía (ÍS =√GD5/PPMA), Ensayo: Sitio de plantación.
Resultados y discusión
Supervivencia
Se detectaron diferencias significativas (P ≤ 0.05) en la supervivencia en los tres periodos de evaluación (Cuadro 2), la cual se mantuvo por encima del promedio reportado como media nacional (36 %) para plantaciones de un año de edad establecidas en México (Burney et al., 2015), al registrarse valores de 84.1 ± 1.4, 76.1 ± 1.6 y 67.9 ± 1.8 % en promedio a los 8, 10 y 12 meses después de la plantación, respectivamente.
Cuadro 2 Supervivencia promedio (± error estándar) de Pinus patula por procedencia.
| Procedencia | Supervivencia promedio (%) | ||
| 8 meses (P ≤ 0.0059) | 10 meses (P ≤ 0.0001) | 12 meses (P ≤ 0.0001) | |
| Ahuazotepec | 73.3 ± 5.7 c | 65.0 ± 6.3 c | 53.3 ± 5.3 c |
| Pinal de Amoles | 81.9 ± 3.7 b | 80.9 ± 3.8 b | 74.3 ± 4.3 b |
| Zacualtipán | 83.3 ± 4.8 b | 80.0 ± 5.2 b | 80.0 ± 5.2 a |
| Acaxochitlán | 85.5 ± 3.7 b | 85.5 ± 3.7 a | 73.3 ± 4.6 b |
| Zacatlán | 81.3 ± 4.5 b | 62.6 ± 5.6 c | 58.6 ± 5.7 c |
| Aquixtla | 88.1 ± 2.7 b | 87.4 ± 2.8 a | 79.2 ± 3.5 a |
| Huayacocotla | 95.2 ± 2.1 a | 71.4 ± 4.4 b | 57.1 ± 4.8 c |
Medias con letras iguales en las columnas no son estadísticamente diferentes (Tukey, P ≤ 0.05).
El 97 % de la mortalidad fue debido a sequía y 3 % por ataque de tuzas (Thomomys umbrinus Richardson). Lo anterior indica que los riegos de auxilio no fueron del todo eficientes, debido quizá a la elevada temperatura en los meses de estiaje, lo que pudo elevar la tasa de evaporación. El 35 % de las plantas del ensayo (220) fueron atacadas por defoliadores (hormiga arriera , Atta cephalotes), aunque se observó rebrote en el 36 % (79) de las plantas afectadas. El 54 % de los brinzales muertos por sequía fueron dañados previamente por el defoliador citado. El 100 % de la mortalidad de las familias de Ahuazotepec, Zacualtipán y Huayacocotla y el 91 % de las plantas muertas de las familias de Aquixtla se debió a la sequía. Las plantas de las familias con mayor daño por defoliador fueron las de Zacualtipán (76 %), mientras que las de Acaxochitlán presentaron el menor daño (55 %). El mayor y menor porcentaje de rebrote se observó en las familias de Aquixtla (40 %) y Zacatlán (26 %).
En dos ensayos de progenies de Pinus patula establecidos en dos sitios de Aquixtla, Puebla, México (2600 y 2980 m de elevación) se reportó supervivencia de 66 a 83 % a los cinco años de edad (Morales et al., 2013). Por su parte, Salaya-Domínguez et al. (2012) registraron 73 y 90 % de supervivencia en ensayos en Aquixtla (2930 m) y Acaxochitlán (2200 m), respectivamente, con daño por heladas y sequía como las principales causas de mortalidad. Si bien, la supervivencia observada en esta investigación es menor que los valores reportados en aquellos establecidos en el centro de México, debe considerarse que las condiciones a las que fue sometida la especie son contrastantes con respecto a las condiciones de los sitios de origen del germoplasma, dada la distancia geográfica que recorrieron vía migración asistida, particularmente en movimiento latitudinal; sin embargo, una supervivencia mayor de 50 % en plantaciones de especies exóticas al primer año de plantado puede considerarse prometedora (Vázquez-Cisneros et al., 2018).
La mortalidad a los 12 meses de edad es entendible, dadas las condiciones contrastantes entre el sitio de plantación y el origen de las familias, pues a pesar de haber aplicado riegos de auxilio, las elevadas temperaturas en los meses de estiaje (> 35 °C de abril a julio) pudieron impactar a los brinzales. La sequía y el calor excesivo son causas de proliferación de plagas forestales, pues las masas arbóreas reducen su capacidad de resistencia a agentes bióticos (Lencinas et al., 2021). Aunque no existe registro de ensayos de progenies en Durango, México se han reportado algunas experiencias en el uso de especies exóticas, principalmente Pinus greggii. Vázquez-Cisneros et al. (2018) obtuvieron 62.4 % de supervivencia en Pinus greggii Engelm. ex Parl. var. greggii al año de plantado en campo. Por otra parte, Álvarez et al. (2018) advirtieron de la presencia del barrenador de yemas (Rhyacionia sp.) en plantaciones de P. greggii establecidas en Durango, presencia que aumentó posiblemente por la disminución de la precipitación, lo que incrementó su tasa de mortalidad. Lencinas et al. (2021) señalaron que las plagas nativas son agresivas con especies introducidas debido a la poca resistencia que presentan cuando existe debilidad por la falta de agua o bajas temperaturas. En el ensayo de P. patula evaluado, la principal causa de mortalidad fue la sequía, particularmente de los brinzales atacados por hormiga arriera, la cual defolia a la planta. Por otra parte, se observó que el rebrote de las plantas se relacionó con su origen; en ese sentido, Aparicio-Rentería et al. (2014) observaron mayor capacidad de rebrote en plantas de P. patula propagadas por estacas de familias de procedencias de mayor elevación.
Características fenotípicas de las familias de Pinus patula evaluadas
Se observaron diferencias significativas (P ≤ 0.05) en la altura y diámetro de planta en los tres periodos de evaluación (Cuadro 3). Las alturas promedio en cada evaluación fueron de 22.8, 30.7 y 43.9 cm, mientras que los diámetros promedio fueron 5.9, 7.7 y 9.3 mm a los 8, 10 y 12 meses después de la plantación, respectivamente.
Cuadro 3 Valores promedio ± error estándar y comparación de medias de Tukey en altura y diámetro de Pinus patula por procedencia.
| Procedencia | Tiempo después de la plantación (meses) | |||||
| 8 | 10 | 12 | ||||
| Altura (m) (P ≤ 0.0002) | Diámetro (mm) (P ≤ 0.0001) | Altura (m) (P ≤ 0.0001) | Diámetro (mm) (P ≤ 0.0001) | Altura (m) (P ≤ 0.0001) | Diámetro (mm) (P ≤ 0.0222) | |
| Ahuazotepec | 23.7 ± 0.8 c | 6.01 ± 0.10 c | 32.1 ± 1.6 c | 7.55 ± 0.13 b | 50.9 ± 2.1 b | 9.19 ± 0.23 b |
| Pinal de Amoles | 20.3 ± 0.5 d | 6.03 ± 0.07 b | 27.6 ± 0.9 d | 7.72 ± 0.09 b | 36.1 ± 0.8 d | 9.52 ± 0.13 a |
| Zacualtipán | 25.2 ± 0.6 b | 6.01 ± 0.11 c | 35.1 ± 1.3 b | 7.56 ± 0.14 b | 51.5 ± 1.7 b | 9.32 ± 0.14 b |
| Acaxochitlán | 20.6 ± 0.5 d | 5.93 ± 0.08 d | 30.2 ± 1.0 d | 7.46 ± 0.10 c | 39.1 ± 1.2 c | 9.20 ± 0.12 b |
| Zacatlán | 20.3 ± 0.6 d | 5.65 ± 0.10 e | 25.7 ± 1.2 e | 7.03 ± 0.15 d | 39.0 ± 1.4 c | 9.11 ± 0.15 c |
| Aquixtla | 19.9 ± 0.4 d | 5.89 ± 0.06 d | 22.9 ± 0.6 f | 7.56 ± 0.07 b | 32.4 ± 0.7 e | 9.26 ± 0.09 b |
| Huayacocotla | 29.9 ± 0.5 a | 6.33 ± 0.06 a | 41.4 ± 1.0 a | 8.02 ± 0.09 a | 58.7 ± 1.3 a | 9.53 ± 0.14 a |
Medias con letras iguales en las columnas no son estadísticamente diferentes (Tukey, P ≤ 0.05).
Gotore et al. (2014) reportaron altura de 0.9 a 1.1 m en plantaciones de P. patula al año de plantado en Zimbabwe en dos sitios de 1000 y 1960 m de altitud, con precipitación media anual de 753 y 1524 mm, respectivamente; aunque no se describe el tamaño inicial ni el método de producción de planta, la altura en dicho estudio fue el doble con respecto a la del presente ensayo, lo que podría deberse principalmente a la baja precipitación del sitio de plantación (296 mm, muy inferior al registro histórico de 576) con respecto al promedio de precipitación del origen de las procedencias (1120 mm). Ante esto, se recomienda el uso de materiales que permitan aumentar el aprovechamiento de la precipitación y reducir los riegos de auxilio, como el uso de hidrogel (Mudhanganyi et al., 2018). Los periodos prolongados de sequía limitan el intercambio de gases y la fijación de CO2 debido al cierre de estomas y afecta el proceso de fotosíntesis (Ding et al., 2020; Laanisto y Niinemets, 2015), lo que se traduce en mayor asignación de recursos al crecimiento radicular y disminución al de altura (Shao et al., 2008).
Componentes de varianza y control genético
Las varianzas de familias y del error incrementaron con la edad del ensayo en las características de altura y diámetro de planta (Cuadro 4), y se espera que la varianza de familias aumente con el paso del tiempo al diferenciarse el crecimiento entre familias. Se ha documentado aumento similar en las mismas variables en progenies de Pinus oocarpa Schiede ex Schltdl. (Fabián-Plesníková et al., 2020) y de P. patula (Salaya-Domínguez et al., 2012). La
Cuadro 4 Parámetros de control genético en las características fenotípicas.
| Variable (meses de edad) | Parámetro de control genético | |||
|
|
|
|
CVGA (%) | |
| Altura (8) | 0.61 | 15.70 | 0.11 ± 0.01 | 6.5 |
| Altura (10) | 8.81 | 60.61 | 0.38 ± 0.14 | 17.2 |
| Altura (12) | 6.12 | 73.38 | 0.23 ± 0.13 | 10.4 |
| Diámetro (8) | 0.02 | 0.42 | 0.20 ± 0.01 | 4.9 |
| Diámetro (10) | 0.07 | 0.66 | 0.29 ± 0.11 | 6.1 |
| Diámetro (12) | 0.07 | 0.93 | 0.23 ± 0.10 | 4.5 |
Dónde:
Viveros-Viveros et al. (2005), en P. oocarpa en ensayos de vivero, determinaron una
Los coeficientes de variación genética aditiva (CVGA) obtenidos para las dos características evaluadas son mayores que los reportados para altura (0.0 a 5.8) y diámetro (0.0 a 3.9) en P. oocarpa en diferentes edades (uno a cinco años) (Fabián-Plesníková et al., 2020). Por su parte, Morales et al. (2013) reportaron que para P. patula el CVGA para altura fue de 6.5 y 9.9 % a los cuatro y cinco años de edad, respectivamente, valores que también fueron superados por el ensayo evaluado en el presente estudio.
La disminución en el CVGA con respecto al aumento en edad (de 10 a 12 meses) podría deberse a que los árboles estaban aún en crecimiento (septiembre) creando disparidades en el desarrollo dentro de cada familia (Fabián-Plesníková et al., 2020; Farfán et al., 2002). Es de esperarse que el CVGA aumente una vez que incremente la diferenciación entre familias por la superioridad en algunas de sus características (Farfán et al., 2002). Aunque el CVGA permite estimar el potencial en la ganancia genética por selección (Vencovsky, 1987), no sería adecuado el realizar una selección temprana (a un año de edad) hasta tener resultados posteriores de supervivencia y crecimiento. Estos resultados pueden mejorar en futuras evaluaciones del ensayo, pues Cotterill y Dean (1988) indicaron que a corta edad no suele expresarse a plenitud el potencial de crecimiento de cada familia; sin embargo, dada la heredabilidad obtenida, dichos autores mencionaron que se podría mejorar la altura total y el diámetro en 12 y 5 % de la siguiente generación si se selecciona el 10 % de las mejores familias para obtener semilla al cruzar entre ellas.
Correlaciones fenotípicas y genéticas
Las correlaciones fenotípicas fueron relativamente altas a los 8 y 10 meses de edad y moderadas a los 12 meses, mientras que las correlaciones genéticas podrían considerarse altas, todas ellas positivas y con tendencia decreciente con el aumento en edad del ensayo (Cuadro 5). Mayor correlación genética se ha observado en P. oocarpa (Viveros-Viveros et al., 2005).
Cuadro 5 Correlaciones fenotípicas (rpf) y genéticas (rg) entre las características fenotípicas.
| Correlación | Edad (meses) | ||
| 8 | 10 | 12 | |
| rpf | 0.67 | 0.68 | 0.43 |
| rg | 0.87 ± 0.01 | 0.76 ± 0.10 | 0.65 ± 0.25 |
La disminución en las correlaciones con el aumento en edad también ha sido documentada por Bustillos-Aguirre et al. (2018), pues observaron una disminución en ambas correlaciones con el aumento en edad de un ensayo de P. patula. Esto también fue reportado para P. oocarpa (Fabián-Plesníková et al., 2020), al menos para la correlación genética de altura-diámetro, al disminuir de 0.91 a 0.74 entre los tres y cinco años de edad, respectivamente. En ese sentido, es de esperarse que las condiciones poco controladas (sequía y ataque de plagas) causen disminución en la correlación de las características por la afectación sobre su desarrollo (Farfán et al., 2002); aun así, los resultados confirman la elevada correlación que existe entre altura y diámetro en especies de coníferas (Morales et al., 2013).
Correlaciones entre las variables respuesta y las variables climáticas del origen de las familias
Se encontraron correlaciones altas negativas y significativas de la altura a las tres edades evaluadas con la elevación y el índice de sequía; alta y positiva con la temperatura mínima, precipitación media anual de los sitios de origen de las familias, mientras que la supervivencia estuvo correlacionada negativamente con la precipitación y positivamente con el índice de sequía a los 10 y 12 meses de edad (Cuadro 6).
Cuadro 6 Correlaciones de Pearson entre altura y supervivencia con las variables de origen.
| Variable del origen de las familias | Altura (m) | Supervivencia (%) | |||
| 8 meses | 10 meses | 12 meses | 10 meses | 12 meses | |
| Elevación | -0.50** | -0.70*** | -0.73*** | NS | NS |
| Temperatura mínima | 0.50** | 0.58** | 0.63** | NS | NS |
| Precipitación media anual | 0.33** | 0.58** | 0.69** | -0.45** | -0.45** |
| Índice de sequía | -0.33** | -0.58** | -0.69** | 0.45** | 0.45** |
** y ***: correlación significativa con P ≤ 0.01 y ≤ 0.001, respectivamente; NS: correlación no significativa.
Las correlaciones negativas indican que la altura es menor en las familias de procedencias de mayor elevación. Las poblaciones de sitios más elevados tienen un crecimiento más lento (Sáenz-Romero et al., 2016), lo que les permite tolerar o evitar daños por bajas temperaturas (Ishizuka et al., 2015), situación que confiere capacidad de sobrevivir en condiciones climáticas adversas (Bansal et al., 2016); además, la altura fue mayor en las familias provenientes de sitios con mayor precipitación, lo cual es normal, puesto que las progenies de estas procedencias suelen crecer más rápido que las provenientes de sitios secos; sin embargo, es de esperarse que las familias de procedencias con mayor precipitación presenten una elevada mortalidad al plantarse en sitios secos, situación que se observó en la presente investigación. Ishizuka et al. (2015) señalaron que las condiciones climáticas, particularmente la precipitación del sitio de origen, influyen de manera importante en el crecimiento de especies forestales.
Conclusiones
La respuesta de Pinus patula a sitios de plantación en Durango, México es variable y depende de su origen, se observó mayor supervivencia en las plantas provenientes de mayor elevación y menor precipitación, aunque fueron las que tuvieron menor crecimiento. Las familias evaluadas mostraron variabilidad genética en las características de altura y diámetro en edades tempranas. Aunque los resultados obtenidos no son concluyentes para realizar selección temprana de familias, son de utilidad para estudios posteriores que evalúen el comportamiento inicial de especies introducidas.

