











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
La subfamilia Bombacoideae (Malvaceae) está compuesta por cerca de 160 especies de árboles que se clasifican en 17 géneros y tres tribus (Carvalho-Sobrinho et al., 2016). La mayoría de las especies son nativas del Neotrópico y tienen reconocida importancia ecológica, económica y cultural, como Ceiba pentandra (L.) Gaertn., Pachira aquatica Aubl. y Pseudobombax ellipticum (Kunth) Dugand (Zidar y Elisens, 2009; Carvalho-Sobrinho et al., 2016; Das et al., 2021). De las tres tribus, Bernoullieae incluye tres géneros, Bernoullia Oliv., Gyranthera Pittier y Huberodendron Ducke, cercanamente emparentados debido a la presencia de inflorescencias escorpioides, cápsulas loculicidas leñosas y semillas aladas (Carvalho-Sobrinho et al., 2016). Hasta el momento, Bernoullia consta de tres especies: B. jaliscana Miranda & McVaugh, endémica de México, B. uribeana Cuatrec., endémica de Colombia, y B. flammea Oliv., presente desde el sur de México hasta Panamá (Fernández-Alonso et al., 2016; Villaseñor, 2016; Cascante-Marín, 2020).
Bernoullia flammea es una especie rara de árbol que llega a alcanzar 40 metros de altura, presenta troncos sin aguijones y gambas cortas, estípulas deciduas, hojas alternas, digitado-compuestas con cinco a siete folíolos, inflorescencias escorpioides anaranjadas, flores anaranjadas bisexuales y pentámeras con un tubo estaminal aplanado y exerto, frutos capsulares con cinco valvas, fusiformes, leñosos y dehiscentes, y semillas aladas (Fig. 1; Holdridge y Poveda, 1975; Cascante-Marín, 2020). En México estos árboles se conocen como cosanté, platanillo, palo de tortilla, palo de calabaza o palo de corcho (Avendaño, 1998). En Costa Rica se conocen como quesofresco, chiverrón, chiverrillo, tayo, yuco o yucón (Fournier y García, 1998; León y Poveda, 1999; datos propios); mientras que en Guatemala como amapola (Mutchnick y Mccarthy, 1997) y en Belice como pumpkin tree (Bridgewater et al., 2006). Los árboles floreados tienen potencial como ornamentales (León y Poveda, 1999). La madera se utiliza como leña y para hacer enchapados en México (Avendaño, 1998; Pietersen et al., 2018). Las semillas son comestibles (Diego-Pérez y Gómez, 2013; datos propios) y en Guatemala se fuma el humo que se obtiene al quemarlas (Das et al., 2021). Además, se sabe que es una especie no pionera que dispersa sus semillas mediante anemocoria (Ibarra-Manríquez et al., 1991; Martínez-Garza et al., 2016; Guzmán-Luna y Martínez-Garza, 2016; Beltrán et al., 2022).

Figura 1: Bernoullia flammea Oliv. A. tronco con gambas; B. árbol en floración; C. hojas digitadas; D. inflorescencias con botones florales; E. flor en antesis; F. fruto inmaduro; G. fruto maduro cerrado; H. fruto maduro abierto; I. semilla. A, C, G, H, I. Fotografías de Diego Aguilar-Sandí. B, D, E y F. Fotografías de Lisa Robertson.
Las investigaciones con B. flammea son escasas y se han llevado a cabo principalmente en la Reserva de la Biosfera Los Tuxtlas, en Veracruz (México), y en Costa Rica. En Los Tuxtlas los estudios se han enfocado en ecofisiología, fenología, restauración ecológica e interacciones con avifauna (Toledo, 1975, 1977; Ibarra-Manríquez et al., 1991; Martínez-Garza et al., 2011, 2016; Guzmán-Luna y Martínez-Garza, 2016; Beltrán et al., 2022), mientras que en Costa Rica en fenología, taxonomía y polinización por aves (Lobo et al., 2008; Alvarado Quesada et al., 2012; Cascante-Marín, 2020). Sin embargo, hasta ahora no se ha evaluado la capacidad germinativa de las semillas ni se ha descrito la morfología de las plántulas de esta especie. En general, tanto la germinación como el desarrollo de las plántulas son etapas del ciclo de vida críticas y poco conocidas en especies de árboles tropicales (Rajjou et al., 2012; Ley y Chacón, 2017).
Si bien B. flammea está clasificada en la categoría de “Preocupación Menor” de la Lista Roja de Especies Amenazadas de la Unión Internacional para la Conservación de la Naturaleza (BGCI & IUCN SSC, 2019), es necesario generar conocimiento que permita contribuir a su conservación. En el caso de Costa Rica, esta especie forma parte de la vegetación nativa del Valle Central Occidental (Holdridge y Poveda, 1975), región del país que alberga la mayor parte de la población costarricense y cuyos bosques originales han desaparecido, quedando en la actualidad únicamente remanentes boscosos (Jiménez y Carrillo, 2016). Contar con información sobre el proceso germinativo y la morfología de las plántulas permitirá desarrollar estrategias de conservación in situ y ex situ dirigidas a propagar esta especie.
Aquí se hipotetiza que las semillas de B. flammea mantienen la viabilidad en el tiempo y que su germinación es independiente de la luz, ya que en Costa Rica se ha observado que la dispersión de las semillas ocurre en la estación seca, cuando aún no ha llovido (obs. pers.), y en Los Tuxtlas se ha reportado que esta especie es no pionera (Martínez-Garza et al., 2016; Guzmán-Luna y Martínez-Garza, 2016; Beltrán et al., 2022). De esta manera, se espera que el porcentaje de germinación, el tiempo medio de germinación y la tasa de germinación sean similares en semillas con distintos tiempos de almacenamiento y que la germinación ocurra tanto en condiciones de iluminación como de oscuridad.
Considerando lo anterior, los objetivos de este trabajo fueron evaluar la germinación de semillas de B. flammea en condiciones de iluminación con diferentes tiempos de almacenamiento y la germinación en oscuridad, así como describir la morfología macroscópica de la plántula.
Materiales y Métodos
Procedencia de los frutos y almacenamiento
El trabajo se llevó a cabo en Costa Rica, con material proveniente de El Güízaro de Atenas, Alajuela (9°57'47.8"N, 84°24'03.6"O; 700 m s.n.m.). La zona presenta estacionalidad climática marcada, con estación lluviosa de mayo a noviembre y estación seca de diciembre a abril (Cambronero et al., 2023). La temperatura media y precipitación media anual rondan los 25 ºC y 1900 mm, respectivamente (Rodríguez-Barboza, 2024). La vegetación corresponde a la de bosque húmedo premontano, según el sistema de zonas de vida de Holdridge (Cambronero et al., 2023).
En febrero de 2023 se recolectaron frutos maduros (i. e., frutos con el pericarpo leñoso y con las líneas de dehiscencia abiertas), de un árbol de B. flammea. Los frutos se colocaron en una bolsa plástica sin cerrar y se trasladaron a la Escuela de Biología de la Universidad de Costa Rica en San Pedro de Montes de Oca, San José, en donde se almacenaron en un cuarto a temperatura ambiente (ca. 24 ºC), hasta ejecutar los experimentos de germinación.
Evaluación de la germinación y descripciones
La capacidad germinativa de las semillas se evaluó mediante dos experimentos realizados en un invernadero con techo y paredes de policarbonato transparente. En el primer experimento se evaluó el efecto del tiempo de almacenamiento en la germinación. Los tratamientos consistieron en: (i) semillas con dos meses de almacenamiento puestas a germinar en condiciones de iluminación con luz natural y fotoperiodo de 12 h, y (ii) semillas con ocho meses de almacenamiento puestas a germinar en condiciones de iluminación con luz natural y fotoperiodo de 12 h. En el segundo experimento se evaluó el efecto de la ausencia de luz en la germinación. El tratamiento consistió en semillas con dos meses de almacenamiento puestas a germinar en condiciones de completa oscuridad. Los resultados de este experimento se compararon con los del primer tratamiento del primer experimento.
Para cada tratamiento se seleccionaron aleatoriamente 100 semillas sin daños visibles. Las semillas se distribuyeron en cuatro recipientes plásticos (cada uno correspondió a una repetición) de 30 × 21.5 × 6.5 cm con turba negra humedecida con agua destilada (25 semillas por recipiente). Se aplicó una disolución de fungicida Amistar® al 0.1% en cada recipiente. Posteriormente, tapados se colocaron dentro de bolsas plásticas transparentes para mantener constante la humedad.
Para lograr condiciones de completa oscuridad, los recipientes plásticos, una vez colocados dentro de las bolsas transparentes, se cubrieron con cuatro capas de papel periódico y se colocaron dentro de una caja de cartón sellada y cubierta con bolsas plásticas oscuras. La temperatura de los invernaderos no se controló, sino que estuvo sujeta a fluctuaciones diarias que oscilaron entre 19 y 33 ºC, según datos obtenidos con un termómetro electrónico de pared marca Extech® (Shanghai, China).
La germinación se evaluó cada tres o cuatro días durante 21 días en los tratamientos expuestos a iluminación y al final de los 21 días en el tratamiento de oscuridad, ya que para ese momento la curva de germinación acumulada presentó un comportamiento constante. Se consideró que una semilla había germinado cuando la radícula emergía a través de la testa.
Todos los análisis estadísticos se realizaron en el programa R v. 4.3.3 (R Core Team, 2024). Los porcentajes de germinación (PG) entre tratamientos se compararon con pruebas de t de Student, mientras que el tiempo medio de germinación (T50) y la tasa de germinación (TG) con pruebas de Wilcoxon. Estos valores se calcularon de la siguiente forma:
La terminología utilizada para describir el tipo de germinación y la morfología de la plántula se tomó de Duke (1969), Copeland y McDonald (2001), Harris y Harris (2001) y Schmidt (2007). Las descripciones se hicieron desde el momento en que la radícula emergió y hasta que las plántulas tuvieron tres meses de edad.
Resultados
Germinación de semillas
La germinación inició a los 7.5±0.5 días (promedio ± error estándar) en ambos tratamientos de almacenamiento. En ese momento, el porcentaje de germinación fue mayor en semillas con ocho meses que en semillas con dos meses de almacenamiento (t=7.44, g.l.=6, p<0.001, Fig. 2A). Sin embargo, no hubo diferencias en el porcentaje final de germinación entre tratamientos (t=1.08, g.l.=6, p>0.05, Fig. 2A). El tiempo de almacenamiento tampoco tuvo un efecto en el número de días necesario para que 50% de las semillas germinaran (T50) ni en la tasa de germinación (Cuadro 1). Tampoco hubo diferencias en el porcentaje final de germinación entre las semillas que germinaron en condiciones de iluminación y de oscuridad (t=-0.30, g.l.=6, p>0.05, Fig. 2B). Adicionalmente, se observó que las plántulas obtenidas en oscuridad mostraron síntomas de etiolación (Fig. 3A), estado que se revirtió al exponerlas a la luz (Fig. 3B).
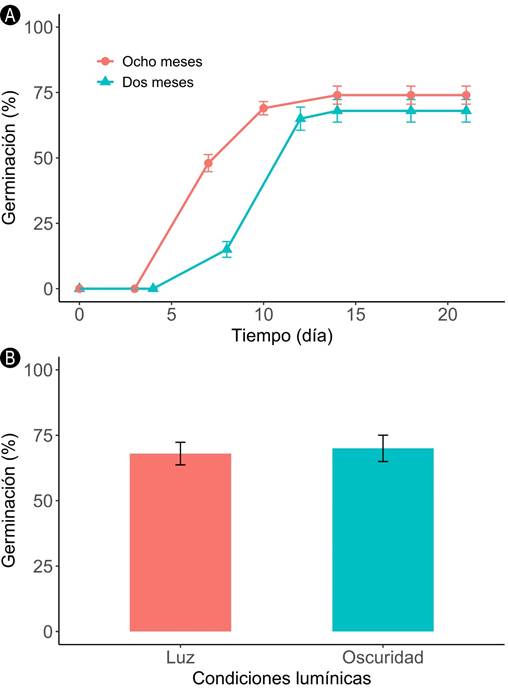
Figura 2: Germinación de semillas de Bernoullia flammea Oliv. A. porcentaje acumulado de germinación (promedio ± error estándar) según el tiempo de almacenamiento; B. porcentaje final de germinación (promedio ± error estándar) según las condiciones lumínicas.
Cuadro 1: Tiempo medio de germinación (T50) y Tasa de germinación (TG) de semillas de Bernoullia flammea Oliv., según el tiempo de almacenamiento. Los valores corresponden a promedios ± error estándar. Letras iguales por columna indican ausencia de diferencias significativas según la prueba de Wilcoxon (p>0.05).
| Tratamiento | T50 (días) | TG (1/días) |
|---|---|---|
| Dos meses | 8.69±2.59a | 0.33±0.24b |
| Ocho meses | 8.30±0.24a | 0.12±0.003b |

Tipo de germinación y morfología de la plántula
Se determinó que la germinación es epigea y fanerocotilar. La radícula emerge a través de la testa, la cual es alada, papirácea y café (Fig. 4A). El hipocótilo se elonga por encima del sustrato, luego los cotiledones se despliegan y la testa cae (Figs. 4B, C). Los cotiledones son opuestos, peciolados, foliáceos, simples, enteros, con venación palmada, cordados, discoloros y fotosintéticos (Figs. 4D, E). Conforme se desarrolla el epicótilo se forman las primeras hojas (eófilos); las cuales son alternas, pecioladas, simples, enteras, con venación palmada, cordadas y concoloras (Fig. 4F). Las estípulas son pequeñas y deciduas (Fig. 4G).
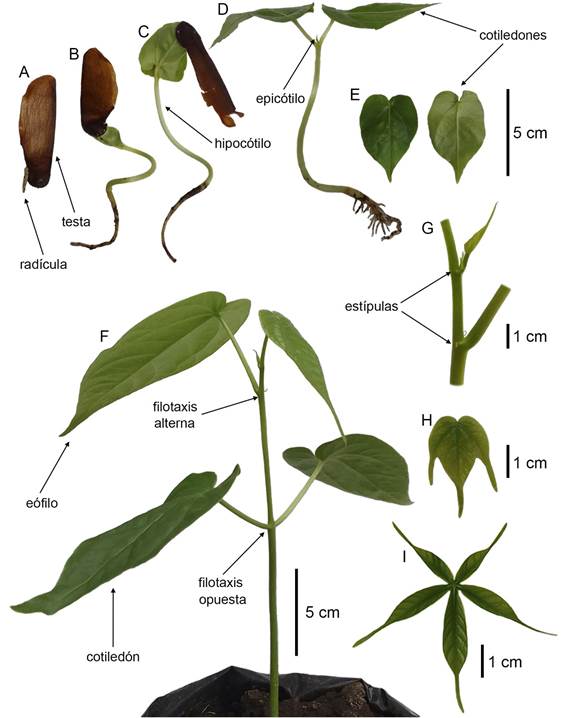
Figura 4: Proceso de germinación y morfología de la plántula de Bernoullia flammea Oliv. A. emergencia de la radícula; B. emergencia de los cotiledones; C. caída de la testa; D. plántula con cotiledones extendidos; E. detalle de los cotiledones; F. plántula con cotiledones y hojas simples enteras; G. detalle de las estípulas; H. hoja simple lobulada; I. inicio de la formación de una hoja compuesta.
Las siguientes hojas son simples lobuladas, generalmente con tres lóbulos (Fig. 4H), aunque bastante irregulares. Las hojas compuestas digitadas (Fig. 4I), típicas de los árboles adultos, se forman cuando las plántulas tienen aproximadamente tres meses de edad. Las plántulas carecen de indumento y tienen una coloración verde clara.
Discusión
Germinación
El momento y las condiciones ambientales que detonan la germinación son fundamentales para garantizar el establecimiento y supervivencia de las plántulas (Lambers et al., 2008; Donohue et al., 2010; Ley y Chacón, 2017). En ambientes estacionales como el Valle Central Occidental y la vertiente del Pacífico de Costa Rica, en donde se encuentra B. flammea (Holdridge y Poveda, 1975; Cascante-Marín, 2020), cabe esperar que el detonante de la germinación sea la llegada de la estación lluviosa, ya que las semillas son dispersadas durante la estación seca (obs. pers.). Esto implica que las semillas deben estar adaptadas para soportar hasta cuatro meses en estado de reposo hasta la llegada de las lluvias. En este sentido, se comprobó que las semillas de B. flammea mantienen la viabilidad hasta por ocho meses, logran germinar durante la primera semana tras la imbibición y alcanzan el T50 al octavo día. Esto indica que son quiescentes, es decir, son semillas maduras con bajos niveles de humedad y poca actividad metabólica que germinan tan pronto entran en contacto con el agua (Copeland y McDonald, 2001; Bewley et al., 2013). No obstante, el contenido de humedad de las semillas no fue determinado en este trabajo.
La quiescencia también explica que no hubiera diferencias en la germinación entre las semillas con dos meses y ocho meses de almacenamiento, pues la quiescencia es un estado de reposo asociado a la capacidad de mantener la viabilidad en el tiempo (Bewley et al., 2013). Sin embargo, los porcentajes finales de germinación nunca superaron 75%. Esto puede deberse a la mortalidad de embriones o a que las condiciones experimentales no estimularon la totalidad de la germinación, por lo que se recomienda estimar el porcentaje de semillas viables antes de la germinación mediante pruebas de viabilidad con tetrazolio (Schmidt, 2007) y determinar el efecto de la temperatura en la tasa de germinación, ya que la temperatura no se controló durante los experimentos. Esto último permitiría determinar la temperatura óptima, valor en el que se obtendría el porcentaje de germinación máximo en el menor tiempo posible (Copeland y McDonald, 2001; Bewley et al., 2013).
Si bien las semillas son quiescentes, aún se desconoce si son ortodoxas o recalcitrantes y, por lo tanto, si son capaces de soportar largos periodos de almacenamiento (Magnitskiy y Plaza, 2007; Rajjou et al., 2012). Para llenar este vacío de información, es necesario recurrir a un diseño experimental que involucre tiempos de almacenamiento prolongados en diferentes condiciones térmicas y determinar, previo a su almacenamiento, el contenido de humedad de las semillas (Schmidt, 2007). De esta manera, se podrá determinar la factibilidad de establecer bancos de semillas de esta especie con fines de conservación a largo plazo (Primack, 1998).
Los resultados también indican que la germinación ocurre tanto en condiciones de iluminación como de oscuridad. Por lo tanto, la luz no es un factor determinante para la germinación, rasgo asociado a semillas de especies no pioneras (Escobar Escobar y Cardoso, 2015), como lo es B. flammea (Martínez-Garza et al., 2016; Guzmán-Luna y Martínez-Garza, 2016; Beltrán et al., 2022). La etiolación de las plántulas confirma que la germinación se dio en condiciones de total oscuridad, pues este estado fisiológico se caracteriza por la ausencia de cloroplastos en plantas que crecen en entornos sin iluminación (Armarego-Marriott et al., 2020).
Morfología de las plántulas
Las descripciones taxonómicas por lo general se basan en organismos adultos en estado fértil, dejando por fuera las características morfológicas de las plántulas, cuya apariencia casi siempre es desconocida (Ley y Chacón, 2017). El tipo de germinación de B. flammea y la morfología de la plántula coinciden, en parte, con el de otras especies anteriormente clasificadas en Bombacaceae, ya que la germinación es fanerocoltilar y las plántulas tienen cotiledones peciolados, estípulas y eófilos alternos con desarrollo gradual (Duke, 1969). Sin embargo, las plántulas de B. flammea se diferencian de otras bombacoides por la forma de los eófilos, los cuales son simples de ápice marcadamente acuminado en Pseudobombax septenatum (Jacq.) Dugand, digitados con 3 a 5 folíolos en Pachira aquatica, trifoliolados de margen entero en P. sessilis Benth., y trifoliolados de margen aserrado en Ceiba pentandra (Ley y Chacón, 2017).
Valga señalar que B. flammea está ausente de los manuales de manejo de semillas de árboles de Centroamérica (Salazar, 2000, 2001; Cordero y Boshier, 2003). Además, en los pocos estudios realizados con plántulas de esta especie no se da información sobre su morfología (Martínez-Garza et al., 2011, 2016; Guzmán-Luna y Martínez-Garza, 2016). En consecuencia, los hallazgos realizados en esta investigación son los primeros que se publican al respecto.
En conclusión, las semillas de B. flammea son quiescentes, mantienen la viabilidad al menos por ocho meses, no requieren de luz para germinar y las plántulas se pueden diferenciar de otras bombacoides con base en la morfología de los eófilos. Con esta información es factible desarrollar programas de reproducción in situ y ex situ, así como realizar estudios poblacionales que tomen en cuenta las plántulas de esta especie.

