










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkUniversidad y ciencia
versión impresa ISSN 0186-2979
Universidad y ciencia vol.24 no.1 Villahermosa abr. 2008
Artículos
Dinámica reproductiva y poblacional del robalo, Centropomus undecimalis (perciformes: Centropomidae), en barra San Pedro, Centla, México
Reproductive and population dynamics of common snook Centropomus undecimalis (Perciformes: Centropomidae) in Barra San Pedro, Centla, Mexico
MA Perera-García1, M Mendoza-Carranza2, S Páramo-Delgadillo3
1 División Académica Multidisciplinaria de los Ríos. Universidad Juárez Autónoma de Tabasco. Carretera Tenosique-Estapilla km 0.5, Colonia Solidaridad S/N, 86901, Tenosique, Tabasco pereragarcia@hotmail.com.
2 ECOSUR Unidad Villahermosa.
3 División Académica de Ciencias Biológicas. UJAT.
Artículo recibido: 25 de junio de 2007
Aceptado: 16 de abril de 2008
Resumen
La biología reproductiva y poblacional fue analizada en poblaciones explotadas de robalo blanco Centropomus undecimalis. El material biológico se obtuvo en los desembarcos de la pesca artesanal de las cooperativas de mayor contribución, en volumen y constancia, en la captura del robalo. La longitud estándar, el peso eviscerado y el sexo de 438 organismos se registró mensualmente, durante 19 meses. Las tallas más frecuentes fueron de 70 a 95 cm de longitud estándar. Se encontraron diferencias significativas entre las longitudes de machos y hembras (Kruskal Wallis; p < 0.05). Los machos representaron el 69% y las hembras el 31%, con una proporción macho:hembra de 2.2:1. La proporción total de sexos presentó diferencias significativas (p < 0.05). La relación longitud-peso no difirió entre hembras y machos (ANCOVA p > 0.05). Se determinó una curva de peso eviscerado, Pev = (0.0106) x (LS)2.9595, con un crecimiento isomètrico (b = 3). Los máximos valores del índice gonadosomático de hembras se registró en agosto de 2004 (1.56) y julio de 2005 (1.52), y para los machos fue en marzo de 2004 (1.67). La longitud estándar (L50) de madurez en hembras y machos fue estimada a los 85 y 68 cm respectivamente, mientras que la longitud estándar de madurez sexual (L100) fue estimada en 100 cm para hembras y 90 cm en machos. Se recomienda la protección de esta especie mediante la disminución de la pesca en las áreas de desove, durante la època reproductiva. Es necesario ampliar los estudios acerca del los procesos migratorios del robalo blanco a travès de los diferentes habitats que esta especie ocupa y como se relacionan con sus eventos reproductivos.
Palabras clave: Pesquería, dinámica poblacional, reproducción, Centropomus undecimalis, Tabasco.
Abstract
The reproductive and population biology of exploited populations of the common snook Centropomus undecimalis was analysed in this study. Biological samples were obtained from the fish landings of the fishing cooperatives with greater constancy and contributions in volume in the snook fishery. The standard length, eviscerated weight and sex of 438 organisms were recorded monthly during 19 months. The most frequent sizes were 70 to 95 cm standard length. Significant differences were recorded between male and female lengths (Kruskal Wallis; p < 0.05). Males represented 69% and females 31%, with a male:female ratio of 2.2:1. The total sex ratio presented significant differences (p < 0.05). The length-weight ratio did not differ between females and males (ANCOVA, p > 0.05). An eviscerated weight curve was determined, Pev = (0.0106) x (LS)2.9595, with an isometric growth of b = 3. The gonadosomatic index recorded its maximum values in August 2004 (1.56) and July 2005 (1.52) for females, and in March 2004 (1.67) for males. The standard maturity length (L50) in females and males was estimated at 85 and 68 cm respectively, whereas the standard sexual maturity length (L100) was estimated at 100 cm for females and 90 cm for males. A decrease in fishing in spawning areas during the reproductive period is recommended to protect this species. It is necessary to increase studies on the migratory processes of the common snook throughout the different habitats it occupies, as well as on how these processes are related to the reproductive events of this species.
Key words: Fishery, population dynamics, reproduction, Centropomus undecimalis, Tabasco.
INTRODUCCIÓN
El robalo blanco, Centropomus undecimalis (Bloch, 1792), es una de las especies más apreciadas por los pescadores artesanales del Golfo de México. Esto se debe a sus características alimenticias, abundancia, alto valor en el mercado local y regional, así como a la facilidad de obtenerlo por parte de las comunidades pesqueras. En México se han identificado 12 especies del género Centropomus, las cuales presentan hábitos diadrómicos y características de ambientes eurihalinos (Muhlia-Melo et al. 1995; Taylor et al. 2000). En el Océano Atlántico se distribuyen seis especies de la familia Centropomidae y en el Océano Pacífico están registradas otras seis especies (Castro-Aguirre et al. 1999; Zarza-Meza et al. 2006). Entre estos centropómidos, el robalo blanco destaca en el Golfo de México por sus volúmenes de capturas, ya que los registros estadísticos oficiales indican que de 1997-2003 se capturaron 12 307 toneladas (Anónimo 2003). Sin embargo, Chacón (1993) mencionó que la pesca del robalo es escasa o, incluso, inexistente, lo cual puede explicarse en función a la marcada estacionalidad en la captura por unidad de esfuerzo (CPUE) del róbalo blanco, principalmente en el sureste del Golfo de México (Caballero 2003). Esta estacionalidad está condicionada tanto por sus hábitos migratorios como sus eventos reproductivos (Lowerre-Barbieri et al. 2003). Así mismo, las variaciones en la CPUE están relacionadas con los cambios en las condiciones ambientales, en donde las anomalías climáticas como El Niño Oscilación del Sur (ENSO) han influido notoriamente en el incremento o disminución del esfuerzo pesquero y han provocado que la captura por pescador sea cada vez menor (Hernández & Kempton 2003; Salas & Gaertner 2004).
Vazzoler (1996) mencionó que los diferentes regímenes de pesca que se adoptan al explotar una población pueden afectar los niveles de mortalidad, así como la edad de primera captura. Es decir, que en la medida que aumenta el nivel de explotación y mortalidad por pesca, la proporción de peces viejos disminuye y, como consecuencia, se reduce el tamaño promedio de los individuos en toda la población (Pitcher et al. 1998; Gómez et al. 2001; Pauly et al. 2002).
La pesca del robalo constituye una de las principales actividades económicas y de subsistencia de las comunidades distribuidas a lo largo del litoral y aguas interiores del Golfo de México, que es una de las zonas de mayor producción de recursos pesqueros (Arreguín et al. 1999; Muller et al. 2001). A pesar de la importancia económica y social de esta especie, en algunas regiones del país como en la costa de Tabasco, no existen estudios recientes y accesibles que aporten información sobre sus aspectos biológicos y ecológicos, lo que obliga, en una primera instancia, a analizar el desarrollo, época reproductiva y determinar la edad de primera madurez sexual. Por lo anterior, el objetivo fue estudiar la biología reproductiva y poblacional de Centropomus undecimalis en la localidad de Barra de San Pedro, Tabasco, México. Este estudio apoyará las decisiones de diversas instituciones que permitan establecer la extracción del robalo en forma sustentable, en zonas y programas de gerencia participativa para la protección de la pesca.
MATERIALES Y MÉTODOS
El muestreo biológico se efectuó en las cooperativas con los centros de recepción mayores volúmenes y constancia, en la captura del robalo. Los ejemplares de C. undecimalis fueron obtenidos al azar de los desembarques de la pesca artesanal realizada en el puerto Barra de San Pedro, Centla (18° 40' N y 93° 39' O) en la costa sur del Golfo de México, (Figura 1). Las muestras fueron recolectadas durante el periodo de enero del 2004 a julio del 2005. Las visitas fueron mensuales con duración de una semana, esto se justifica por ser lo más recomendable para la obtención de datos pesqueros de alta confiabilidad y poder obtener un muestreo estadístico y biológicamente significativo (Gulland 1983). El área de influencia de estas cooperativas se extiende sobre la costa del Golfo de México, aproximadamente por 15 km a ambos lados de la desembocadura del Río San Pedro y San Pablo y mar adentro. Dentro de su concesión de pesca están también incluidas las lagunas interiores asociadas con este río.

Para cada individuo se registró datos de longitud estándar (LS) en centímetros y peso eviscerado (Pev) en gramos. Los datos de longitud permitieron determinar la estructura de la población, a través de un histograma de frecuencia de tallas. Para la obtención de las gónadas, a cada ejemplar se le realizó un corte ventral, se extrajeron, midieron y pesaron en una balanza de 0.01g de precisión, se determinó el sexo y el grado de madurez gonádica (Csirke 1993).
La relación talla-peso puede ser utilizado para medir la variación del peso esperado para un pez o grupos de peces, como indicador de su condición fisiológica o grado de bienestar y desarrollo gonádico (Rodríguez 1992, Sparre & Venema 1995). Cabe señalar que a través de esta relación es posible comparar grupos de peces y es también una herramienta en las estimaciones de biomasas y evaluación de reservas de peces (Gulland 1983; Safran 1992; Entsua-Mensah et al. 1995).
El Pev y la LS se relacionaron para cada sexo. Ambos parámetros fueron ajustados a la siguiente función alométrica (Ricker 1958).
W=a Lb
donde W es el peso eviscerado en g, a es una constante, L es la longitud estándar en cm, y b el exponente de la ecuación que al transformarse a una función lineal se convierte en la pendiente de la recta y denota la isometría o alometría en el crecimiento.
El sexo y la madurez sexual de cada ejemplar se determinó con base en las características morfológicas y coloración de la gónada según criterio establecido por Nikolsky (1963) y por Csirke (1993) para desovadores parciales, en los cuales la escala de maduración de las gónadas fue modificado por Overstreet (1983) y Taylor et al. (1998). Esta información sirvió para determinar la proporción mensual de hembras y machos en las muestras durante el período de estudio.
La actividad reproductiva se determinó a través del índice gonadosomático (IGS) a partir de la formula

donde: PG es el peso de la gónada en g y Pev es el peso eviscerado en g de cada individuo.
La longitud de primera maduración gonadal para el 50 % de la población (L50 %) y en la que todos los individuos están aptos para participar activamente en el proceso reproductivo (L100 %) fue obtenida a partir de frecuencias porcentuales de individuos adultos de machos y hembras, con estadios macroscópicos de maduración entre II y VI (Vazzoler 1996).
La menor talla, en la cual al menos un ejemplar muestra su máximo desarrollo gonadal o talla mínima de primera madurez, se determinó por el criterio de Pitt (1964), el cual fue citado por Marcano et al. (2002) en su trabajo sobre Cynoscion jamaicensis.
Análisis estadístico
Las diferencias de las medias mensuales entre las longitudes estándar de machos y hembras fueron comparadas mediante un análisis de varianza no paramétrico de Kruskal-Wallis (Zar 1984; Sokal & Rohlf 1996), dado que los datos no cumplíeron los supuestos de normalidad y homogeneidad de varianzas para aplicar un ANOVA, (Kolmogorov-Smirnov; p = 0.01 y Levene; p = 0.01).
Análisis de correlación (Gulland 1983; Sparre & Venema 1995), y de covarianza (ANCOVA) fueron aplicados a los valores a y b relativos a las regresiones entre sexos para constatar posibles diferencias entre ambos. Las ecuaciones alométricas de crecimiento se obtuvieron por medio de una transformación logarítmica (Zar 1984; Sokal & Rohlf 1996).
Las proporciones de sexos mensuales y por clase de longitud se analizaron mediante una Chi cuadrada (Underwood 1997). Las proporciones de sexos mensuales y por clase de longitud se analizaron mediante una Chi cuadrada (Underwood 1997).
Los valores de madurez sexual se suavizaron a través de una función logística, debido a que las curvas presentaron discontinuidades en su conformación (Gaertner & Laloe 1986; Sokal & Rohlf 1996; Vazzoler 1996).
RESULTADOS
Estructura de tallas
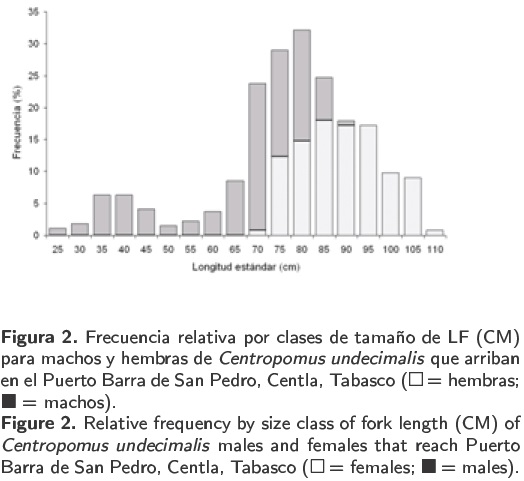
El intervalo de la longitud estándar de 438 organismos de robalo blanco fue de 25 a 110 cm. Las tallas más frecuentes fueron de 70 a 95 cm con una moda a los 80 cm (Figura 2). En los machos, la longitud mínima (25 cm) se presentó en marzo de 2004 y la longitud total máxima (91.2 cm) se registró en abril de 2005. En las hembras, la longitud estándar mínima fue de 70 cm en diciembre de 2004 y la longitud estándar máxima de 110 cm en marzo de 2005 (Figura 3). Entre las medias de las longitudes estándares de machos y hembras se estimaron diferencias significativas (Kruskal-Wallis; p < 0.05).

Relación longitud - peso
Entre las curvas de la relación longitud-peso entre hembras y machos no se estimaron diferencias significativas (F = 2.24, p = 0.13). Para las hembras la ecuación fue Pev= 0.0113 (LE)2'949 y para los machos Pev = 0.0117 (LE)2,9357. El coeficiente de correlación de Pearson entre las dos variables fue de 0.82 para hembras (n = 122, p = 0.001) y 0.97 para los machos (n = 270, p = 0.001). La ecuación general para ambos sexos fue Pev= 0.0106 (LE)2'9595 (Figura 4). Esta ecuación indica que el crecimiento de C. undecimalis es de tipo isométrico.

Proporción de sexos
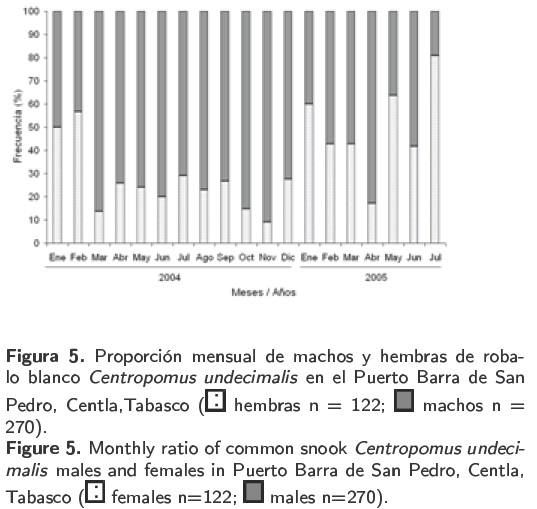
Del total de organismos analizados, los machos representaron el 69 % y las hembras el 31 %, resultando una proporción macho:hembra de aproximadamente 2.2:1. En la mayoría de los meses muestreados (marzo a diciembre de 2004 y febrero-abril y junio de 2005) la proporción de machos fue mayor alcanzando su máximo en el mes de noviembre de 2004 con 91 %. Para las hembras sus mayores proporciones fueron observadas durante el mes de febrero de 2004 y enero, mayo y julio de 2005 (Figura 5). La proporción total de machos y hembras, difirieron significativamente (Chi cuadrada; p < 0.05) en los meses de (marzo-mayo, julio a diciembre de 2004, en abril y julio de 2005). La proporción total de sexos presentó diferencias significativas (p < 0.05). Del total de organismos analizados, los machos representaron el 69% y las hembras el 31%, resultando una proporción macho:hembra de aproximadamente 2.2:1. En la mayoría de los meses muestreados (marzo a diciembre de 2004 y febrero-abril y junio de 2005) la proporción de machos fue mayor alcanzando su máximo en el mes de noviembre de 2004 con 91 %. Para las hembras sus mayores proporciones fueron observadas durante el mes de febrero de 2004 y enero, mayo y julio de 2005 (Figura 5). La proporción total de machos y hembras, difirieron significativamente (Chi cuadrada; p < 0.05) en los meses de (marzo-mayo, julio a diciembre de 2004, en abril y julio de 2005). La proporción total de sexos presentó diferencias significativas (p < 0.05).
La proporción de sexos por clases de tallas, fue favorable para los machos de los 25 a 65 cm, con máximos de (100%) en las longitudes de 25 a 70 cm. Esta proporción se invirtió a partir de los 85 cm y las hembras obtuvieron su máximo (100 %) de los 95 a 110 cm (Figura 6). En la proporción de sexos de casi todas las clases de tallas se estimaron diferencias significativas (Chi cuadrada; p < 0.05). La excepción fue en la clase de 85 cm, donde no hubo diferencias significativas (Chi cuadrada; p > 0.05).

Indice gonadosomático y madurez sexual de la población
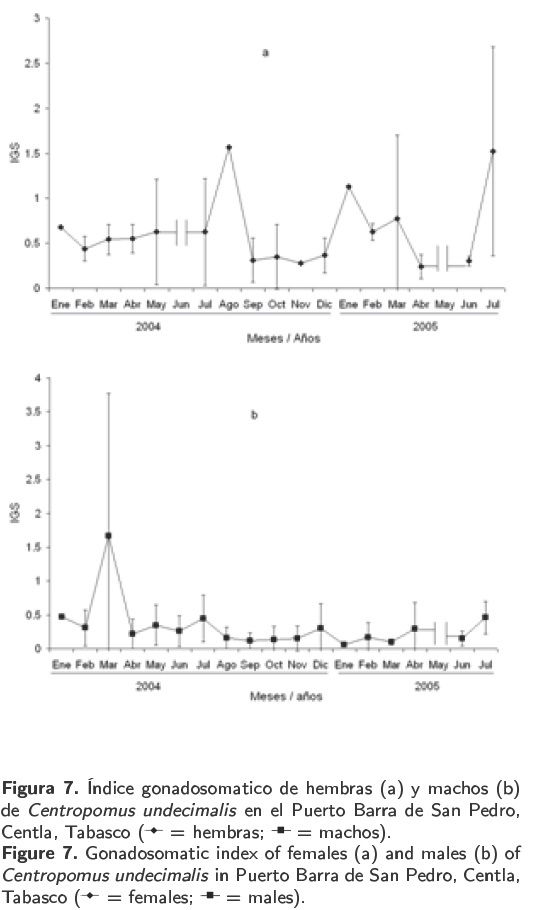
El índice gonadosomatico (IGS) mensual de hembras de C. undecimalis presentó sus valores máximos durante los meses de agosto (1.56) de 2004 y julio (1.52) de 2005. En los meses restantes el valor fue menor a uno (Figura 7a). Por el contrario, en los machos el valor máximo se registró en marzo (1.67) y los demás meses presentaron valores menores a uno, con un mínimo de 0.05 en el mes de enero de 2005 (Figura 7b).
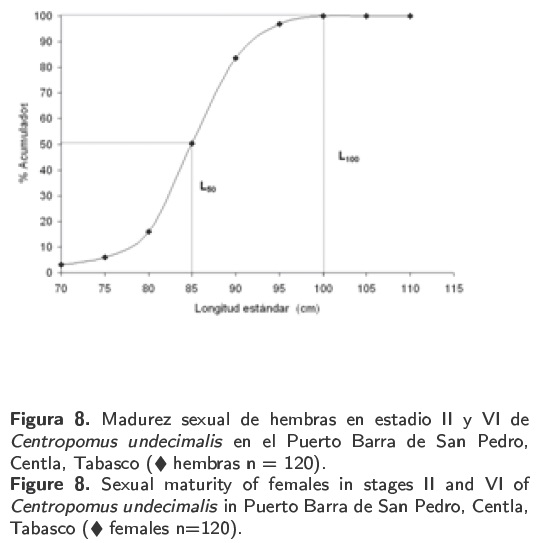
La talla a la cual maduró el 50% de la población (L50) fue a los 85 cm de longitud estándar para las hembras y 68 cm para los machos. Mientras que la talla de los individuos activos para participar en el proceso reproductivo (L100) fue estimado en 100 y 90 cm para hembras y machos, respectivamente (Figura 8 y 9).

DISCUSIÓN
Durante el período de estudio se determinó una estructura trimodal para el robalo blanco, la cual permite suponer que existen dos grupos de edades o cohortes, cuyas tallas fluctuaron entre los 25 y 110 cm (LE). Ruiz (1993) mencionó que los robalos suelen alcanzar tallas considerables y varían conforme a la especie, con intervalos de 15 a 60 cm para las especies del pacifico y de 30 a 90 cm para las especies del Golfo de México. Sin embargo, Tringali et al. (1999) registraron peces de 139.5 cm de longitud total (LT) en la costa de Florida, E.U.A. La variación en la estructura de tallas es determinada por la selectividad del arte de pesca, el tamaño de la misma y la técnica de pesca utilizada (Grimes 2001; Pauly & Watson 2001; Hernández & Kempton 2003). También, la distribución de tallas puede variar de una región a otra y hasta dentro de una misma región (Matsuyama et al. 1987; Vazzoler 1996), ya que el crecimiento es afectado por la disponibilidad de alimento, densidad poblacional y por factores ecológicos (Marcano et al. 2002).
Los resultados obtenidos por Osorio & González (1986) en Venezuela coincidieron con este trabajo, donde la longitud y el peso total de las hembras fue superior al de los machos. Sin embargo, ni en la diferencia en longitud, ni la del peso total resultaron significativamente diferentes, lo que puede significar que la diferencia en el crecimiento de machos y hembras se debe más a factores genéticos, que determinan la fisiología y reproducción del pez, que a los factores ambientales. Posiblemente, ellos no consideraron la condición protándrica de esta especie registrada por Taylor et al. (2000).
La magnitud del exponente "b" derivada de este estudio para C. undecimalis, coincidió con lo registrado en el suroeste de Campeche, donde se estimó que el valor de "b" para ambos sexos fue de 2.94, lo cual indica que esta especie es de crecimiento isométrico (Aliaume et al. 2005). Para estimar el tipo de crecimiento de una población, se debe especificar la época de muestreo y los intervalos de las variables "a" y "b" en que fueron muestreadas, debido a que esta relación de crecimiento es particular para cada especie en un determinado estadio, desarrollo ontogénico y sexo (Bouchon-Navaro et al. 2006; Rodrigues et al. 2006). Los cambios en la alimentación y épocas de reproducción influyen en el crecimiento y en las variaciones temporales de la biomasa de la población (Torres-Lara & Salas-Márquez 1990).
En la composición por sexos, los machos dominaron en relación a las hembras con una proporción macho: hembra en casi todos los meses de muestreo de 2.2:1. Esta proporción difiere a las obtenidas para C. undecimalis en Isla Margarita, Venezuela de 1:1 en un año de estudio (Osorio & González et al. 1986) y de 1:1.6 para Centropomus medius (Maldonado-García et al. 2005). En contraste, la proporción de sexos de este estudio coincidió con la calculada solamente para agosto (2:1) por Osorio & González et al. (1986) y la registrada para C. undecimalis (2:1) en el suroeste de Campeche (Caballero 2003).
Por clases de tallas se pudo observar que a partir de los 90 cm de longitud estándar, la mayoría de los organismos fueron hembras debido probablemente a un cambio de sexo por su condición hermafrodita protándrica (Taylor et al. 2000). Al respecto, el hermafroditismo protándrico, el potencial reproductivo de los individuos machos ha sido registrado mayor en tallas mayores (Vazzoler 1996; Grier 2000; Devlin & Nagahama 2002).
La interpretación de la proporción de sexos y estructura de tallas los datos obtenidos pueden ser afectada por el tipo de arte de pesca utilizado, debido a que usualmente se capturan organismos de tallas menores en menor cantidad de la que realmente podría ser una muestra representativa de la población (Pauly & Watson 2001).
El mérito del empleo del índice gonadosomático es que, aunque tal índice no muestra una información precisa del máximo desove, sí indica la época máxima de desarrollo gonádico y determina el periodo cuando se encuentra una mayor cantidad de productos sexuales (Quiroga & Solís 1999; Rodrigues 2006). Los valores más altos, indican el periodo máximo de madurez sexual. En este estudio, el reducido número de organismos recolectados para hembras y machos, así como a la falta de obtención de la longitud y peso de las gónadas en los meses de (junio de 2004 y mayo de 2005) pudo haber provocado una estimación sesgada del IGS medio mensual.
El periodo reproductivo descrito en este trabajo difiere con otros estudios realizados con C. undecimalis, donde se ha indicado generalmente un periodo reproductivo de abril a mayo y diciembre, con máximos valores de desoves entre mayo y agosto (Osorio & González 1986; McMichael et al. 1989; Taylor et al. 1998).
El desove de esta especie es asincrónico, en la época de reproducción se presenta un desfase, el cual provoca que haya muchos individuos con un desove parcial característico de las regiones tropicales (Wallace et al. 1993; Tobias 2001; Lowerre-Barbieri et al. 2003). No obstante, los centropómidos presentan desoves sincrónicos cuando están agregados, lo cual es una características de los teleósteos (Alonzo & Mangel 2005; Maldonado-García et al. 2005).
La talla en la que madura el 50% de la población (L50), se utiliza como estrategia de administración en el ordenamiento pesquero, para que las especies sean capturadas a una talla mayor de la primera madurez. Las hembras registraron esta característica a los 85 cm y los machos a los 65 -70 cm de longitud total. La talla en que los individuos están activamente participando en el proceso reproductivo fue estimada en 100 cm para hembras y 90 cm en machos. Los resultados obtenidos resultaron similares a los registrados por Caballero (2003) en Campeche, donde el 50% de organismos maduros presentaron una longitud de 79 cm de longitud furcal, la L50 fue de 82-83 cm para hembras y los machos entre 79-80 cm y el 100 % de las hembras de 97 cm estuvieron maduras y a los 91 cm los machos. La posibilidad de que a estas tallas, casi el 100 % de los ejemplares sea susceptible al cambio de sexo, se sugiere ya que en una población de esta especie, los peces más grandes y viejos son hembras. En este sentido, un grupo de robalos que pierde a sus peces mas grandes ha perdido hembras, por lo que algunos de los machos experimentan el proceso de reversión sexual, el cual tarda de 60 a 90 días (Grier & Taylor 1998; Pope et al. 2006). La primera madurez sexual registrada para los machos de C. undecimalis en Florida, E.U.A., fue entre 15 y 20 cm de longitud estándar (Taylor et al. 2000). La probables diferencias en los datos se deban a las diferentes zonas de estudio y a las condiciones climáticas, relacionados principalmente con la temperatura del agua entre Campeche, Tabasco y Florida.
El cambio de la mortalidad por pesca y la edad a la primera captura, son parámetros poblacionales que el hombre puede controlar, cuando aumenta o disminuye su capacidad de captura u orienta sus faenas de pesca hacia captura de ejemplares de menor o de mayor tamaño (Csirke 1993). De acuerdo con el régimen de pesca que se adopte, la población puede registrar cambios mayores o menores en el número y la biomasa total de sus integrantes, en su estructura por edad o por tamaños, en su velocidad de crecimiento e inclusive en su capacidad de reproducción.
Marcano et al. (2002) señalaron que la estimación a través de la curva linearizada de CPUE minimiza los posibles sesgos por errores en los muestreos y en las diferencias de algunas frecuencias de tallas de los ejemplares. Las tallas, desde el punto de vista del manejo del recurso, deben ser consideradas fundamentales para la selectividad de los artes de pesca empleados en la explotación, ya que permitiría el escape de los ejemplares de tallas inferiores.
Los datos del índice gonadosomático, en el contexto acuacultural, son importantes, pues permiten al acuicultor seleccionar por su talla a las hembras reproductoras del medio natural y en la época con mayor probabilidad de encontrarlas maduras sexualmente, dentro de la talla en que haya una mayor probabilidad de encontrarlas maduras. En el caso del robalo, sería a tallas mayores de 90 cm. Sin embargo, hay que considerar de que esta información sirve de guía y no determina de alguna forma, que siempre serán encontradas hembras con tallas mayores (Taylor et al. 2000). La presencia de individuos en transición en los meses inmediatamente siguientes a la época de mayor madurez sexual, indica que el cambio de sexo no ocurre durante la época reproductiva, sino posteriormente (Peters et al. 1998; Grier 2000). Las hembras originadas de este cambio sexual estarán maduras para el siguiente periodo reproductivo como ocurre con otros peces hermafroditas (Lozano & Olaya-Nieto 2004). Aunque, Sánchez (datos no publicados) encontraron que el cambio de sexo de un especimen de 2 kg en cautiverio se dió en el transcurso de un mes. Dicho pez fue macho a principios de junio y a finales de ese mismo mes se determinó como hembra en vitelogenésis.
Actualmente, en Tabasco se desconoce la magnitud del efecto de la pesca sobre las poblaciones de Centropomus undecimalis. Por lo que, es necesario establecer programas de monitoreo a largo plazo, con la finalidad de obtener datos confiables para la toma de decisiones sustentadas en la biología y dinámica pesquera del recurso. La continuación de estudios sobre la actividad reproductiva y reclutamiento de la especie, tanto en tiempo como geográficamente, constituirían una importante base para la administración del recurso. El desarrollo de programas que incluyan aspectos biológicos y datos ambientales, como temperatura, salinidad y precipitación pluvial como apoyo, pueden ayudar a explicar los patrones reproductivos de esta especie a largo plazo. Así, esta investigación es un marco de referencia para futuros trabajos sobre pesquería del robalo en la costa y zonas ribereñas de Tabasco.
Dentro de las estrategias de administración pesquera generalmente se busca que las especies sean capturadas a una talla y edad (de reclutamiento) mayor que la de primera reproducción. Los resultados obtenidos al respecto revelan que C. undecimalis se recluta a la población reproductora a los 85 cm para hembras y de 65 a 70 cm para machos, lo que revela que desde este punto de vista el recurso no está siendo adecuadamente explotado en el área de estudio, debido a que se refleja una disminución en los volúmenes de capturas.
AGRADECIMIENTOS
Al ECOSUR y CONACYT por la facilidades prestadas, a Maricela Huerta Ortiz, Noe Mejia, Darwin Manolo, en la obtención de datos, a Rodimiro Ramos Reyes de Ecosur, a Mayte Perera colaboradora de la FAO y a los revisores por sus acertadas y valiosas observaciones.
LITERATURA CITADA
Aliaume C, Zerbi A, Miller JM (2005) Juvenile snook species in Puerto Rico estuaries: Distribution, abundance and habitat description. Proc. Gulf Carib. Fish. Institute (47): 499-519. [ Links ]
Anónimo (2003) Anuario Estadístico de Pesca. SAGARPA. Distrito Federal. 265 pp. [ Links ]
Alonzo SH, Mangel M (2005) Sex-change rules, stock dynamics, and the performance of spawning-per-recruit measures in protogynous stocks. Fish. Bull. 103: 229-245. [ Links ]
Arreguín F, Sánchez J, Flores HD, Ramos MJ, Sánchez GP, Arancibia AY (1999) Stock-Recruitment Relationships: A scientific Challenge to Support Fisheries Management in the Campeche Bank, Mexico. En Kumpf H, Steidinger K, Sherman K (eds) The Gulf of Mexico Large Marine Ecosystem: Assessment, Sustainability and Management. Blackwell Science. Inc. Malden, USA. 255-280 pp. [ Links ]
Bouchon-Navaro Y, Bouchon C, Kopp D, Louis M (2006) Weight-length relationships for 50 fish species collected in seagrass beds of the Lesser Antilles. J. Appl. Ichthyol. (22): 322-324. [ Links ]
Caballero CV (2003) Estudio biológico pesquero del robalo Centropomus undecimalis en el Suroeste de Campeche. Facultad de Ciencias. Universidad Nacional Autónoma de México, Distrito Federal. 71 pp. [ Links ]
Castro-Aguirre JL, Espinosa-Pérez H, Schmitter-Soto JJ (1999) Ictiofauna estuarino-lagunar y vicaria de México. Colección textos politécnicos. Serie biotecnológicas. Noriega-Limusa. Distrito Federal. 711 pp. [ Links ]
Chacón D (1993) Aspectos biométricos de una población de sábalo, Megalops atlanticus (Pisces: Megalopidae). Rev. Biol. Trop. 41: 13-18. [ Links ]
Csirke J (1993) Introducción a la dinámica de poblaciones de peces. FAO, Doc. Téc. Pesca (192): 82. [ Links ]
Devlin HD, Nagahama Y (2002) Sex determination and sex differentiation in fish: an overview of genetic, physiological, and environmental influences. Aquaculture (208): 191-364. [ Links ]
Entsua-Mensah MA, Osci-Abunyewa R, Palomares M (1995) Length-weight relationship of fishes from tributaries of the Volta river, Ghana. Part I. [ Links ]
Analysis of pooled data sets. ICLARM Q, 18(1/2): 36-38. [ Links ]
Gaertner D, Laloe F (1986) Etude biometrique de la talle a' premiere maturità sexualle de Geyron maritae, Maning et' Holthuis, 1881 de Senegal. Oceanol. Acta 9(4): 479-486. [ Links ]
Gómez GR, Guzmán R, Astudillo L (2001) Análisis de la captura, esfuerzo y rendimiento (CPUE) de la pesquería artesanal con palangre en el golfo de Paria, estado Sucre, Venezuela. Memorias Acta Científica Ven. 52 (Supl. 3): 26-29. [ Links ]
Grier H (2000) Ovarian germinal epithelium and folliculogenesis in the common snook, Centropomus undecimalis (Teleosteo: Centropomidae). Journal of Morphology 243 (3): 265-281. [ Links ]
Grier HJ, Taylor RG (1998) Testicular maturation and regression in the common snook. J. Fish Biol. (3): 521-542. [ Links ]
Grimes CB (2001) "Fishery production and the Mississippi river discharge". Fisheries 26(8):17-26. [ Links ]
Gulland JA (1983) Fish stock assessment: a manual of basic methods Vol. 1. Wiley Interscience, FAO/Wiley series on food and agriculture. Chischester. 223 pp. [ Links ]
Hernández A, Kempton W (2003) Changes in fisheries management in Mexico: Effects of increasing scientific input and public participation. Ocean & Coastal Management 46: 507-526. [ Links ]
Lozano GE, Olaya-Nieto CW (2004) Reproductive aspects of common snook (Centropomus undecimalis) in the Cispata bay, Colombia. Proceedings of the Gulf and Caribbean Fisheries Institute (55): 1029. [ Links ]
Lowerre-Barbieri SK, Vose FE, Whittington JA (2003) Catch-and-release fishing on a spawning aggregation of common snook: does it affect reproductive output. Trans. Am. Fish. Soc. 132: 940-952. [ Links ]
Maldonado-García M, Gracia-López V, Carrillo M, Hernández-Herrera A, Rodríguez-Jaramillo C (2005) Stages of gonad development during the reproductive cycle of the blackfin snook, Centropomus medius Günther. Aquaculture Research 36 (6): 554-563. [ Links ]
Marcano L, Alió J, Altuve D (2002) Biometría y talla de primera madurez de la tonquicha, Cynosción jamaicensis, de la costa norte de la península de Paria, estado Sucre, Venezuela. Zootecnia Tropical 20(1): 83-109. [ Links ]
Matsuyama M, Matsuura S, Hidaka T (1987) Maturity classification and group maturity of the red sea bream Pagrus major. Mar. Biol. 96: 163-168. [ Links ]
McMichael RH, Peters KM, Parsons GR (1989) Early life history of the snook, Centropomus undecimalis in Tampa Bay, Florida. Northeast Gulf Science 10(2):113-126. [ Links ]
Muhlia-Melo A, Arvizu-Martínez J, Rodríguez-Romero J, Guerrero-Tortolero D, Gutiérrez-Sánchez FJ, Muhlia-Almazán A (1995) Sinopsis de información biológica, pesquera y acuacultural a cerca de los robalos del género Centropomus en México. Programa de Evaluación de Recursos Naturales del Centro de Investigaciones Biológicas del Noreste, S. C. Volumen Especial. Centro de Investigaciones Biológicas del Noreste, S. C. La Paz. 52 pp. [ Links ]
Muller RG, Murphy MD, Kennedy FS (2001) The 2001 stock assessment update of common snook, Centropomus undecimalis. Fish and Wildlife Conservation Commission, Florida Marine Research Institute. St. Petersburg. 48 pp. [ Links ]
Nikolsky GV (1963) The ecology of fishes. Academic Press. Nueva York. 352 pp. [ Links ]
Osorio J, González L (1986) Aspectos reproductivos del robalo Centropomus undecimalis (Bloch, 1792) (Pisces: Centropomidae) de la Isla Margarita, Venezuela. Contr. Cient. (9): 1-35. [ Links ]
Overstreet R (1983) Aspects of the biology of the spotted sea trout, Cynoscion nebulosus, in Mississippi Gulf. Res. Rep. Supp. 1: 1-43. [ Links ]
Pauly D, Watson R (2001) Systematic distortions in world fisheries catch trends. Nature Magazines. Fisheries Centre, University of British Columbia. Ottawa. 414 pp. [ Links ]
Pauly D, Christensen V, Guenette S, Pitcher TJ, Zeller D (2002) Towards sustainability in world fisheries. Nature 418: 689-695. [ Links ]
Peters KM, Matheson RE, Taylor RG (1998) Reproduction and early life history of common snook, Centropomus undecimalis (Bloch), in Florida. Bull. Mar. Sci. 62: 509-529. [ Links ]
Pitcher TJ, Hart PB, Pauly D (1998) Reinventing Fisheries Management. Kluwer. London. 435 pp. [ Links ]
Pitt T (1964) Fecundity of American plaice, Hippoglosoides platessoides from Green Bank and new Underland avers. J. Fish. Res. Bd. Canada 21 (3): 597-612. [ Links ]
Pope KL, Blankinship DR, Fisher M, Patiño R (2006) Status of the common snook (Centropomus undecimalis) in Texas. Texas Journal of Science 58: 325-332. [ Links ]
Quiroga BC, Solís CF (1999) Estado actual de la pesquería de robalo en México, Pesquerías Relevantes de México. XXX Aniversario del INP. SEMARNAP/INP. (IV): 559-578. [ Links ]
Ricker WE (1958) Handbook of computation for biological statistics of fish populations. Bull. Fish. Res. Board Can. (119): 300 Rodrigues PP, [ Links ] Oliveira NL, Martins AS (2006) Fat snook (Centropomus parallelus) reproductive aspects in the Doce River estuary, east coast of Brazil. En: Mees J; Seys J (ed) VLIZ Young Scientists' Day, Brugge, Belgium 31 March 2006: Book of abstracts. VLIZ Special Publication 30: 52 pp. [ Links ]
Rodríguez GM (1992) Técnicas de Evaluación Cuantitativa de la Madurez Gonádica en Peces. AGT S.A. Distrito Federal. 79 pp. [ Links ]
Ruiz DMF (1993) Recursos Pesqueros de las Costas de México, su Conservación y Manejo Socioeconómico. Limusa. Distrito Federal. 208 pp. [ Links ]
Safran P (1992) Theoretical analysis of the weight-lenght relationship in fish juveniles. Mar. Biol. 112, 545-551. [ Links ]
Salas S, Gaertner D (2004) The behavioral dynamics of fishers: management implications. Fish and Fisheries 5: 153-167. [ Links ]
Sokal R, Rohlf F (1996) Biometry. Freeman Company. San Francisco. 887 pp. [ Links ]
Sparre P, Venema SC (1995) Introducción a la evaluación de recursos pesqueros tropicales, Parte 1 manual. FAO Documento técnico de pesca No. 306. Santiago de Chile. 420 pp. [ Links ]
Taylor G, Grier HJ, Witthington JA (1998) Spawning rhythms of common snook in Florida. J. Fish. Biol. 53: 502-520. [ Links ]
Taylor GR, Wittington JA, Grier HJ (2000) Age growth, maturation and protandric sex reversal in the common snook Centropomus undecimalis, from the east and west coasts of south Florida. Fish. Bull. 98: 612-624. [ Links ]
Tobias WJ (2001) Mangrove habitat as nursery grounds for recreationally important fish species - great pond, St. Croix, USA. virgin islands. Proc. Gulf Carib. Fish. Institute (52): 468-487. [ Links ]
Torres-Lara R, Salas-Márquez S (1990) Crecimiento y mortalidad de la rubia Lutjanus synagris de las costas de Yucatán durante las temporadas de pesca 1983-1985. Anales Inst. Cienc. Mar Limnol. Univ. Nal. Autón. México 17 (2): 205-214. [ Links ]
Tringali MD, Beter TM, Seyoum S, Berminigham E, Bartolacci D (1999) Molecular Phylogenetics and ecological diversification of the transisthmian fish genos Centropomus (Perciformis: Centropomidae). Mol. Phylogenet. Evol. 13: 193-207. [ Links ]
Underwood AJ (1997) Experiments in ecology-Their logical desing and interpretation using analysis of variance. Cambridge University Press. Cambridge. 504 pp. [ Links ]
Vazzoler AEA (1996) Biologia da reproducäo de peixes teleósteos: Teoria e prática. Maringá: EDUEM. Maringá. 169 pp. [ Links ]
Wallace RA, Boyle SM, Grier HJ, Selman K, Petrino TR (1993) Preliminary observations on oocyte maturation and other aspects of reproductive biology in captive female snook, Centropomus undecimalis. Aquaculture 116(2-3): 257-273. [ Links ]
Zar JH (1984) Biostatistical Analysis. Prentice International, Inc. Englewood Cliffs. 718 pp. [ Links ]
Zarza-Meza AE, Villalobos JB, Vásquez CP, Álvarez PT (2006) Cultivo experimental del robalo Centropomus undecimalis (Bloch, 1792) y Chucumite Centropomus parallelus (Poey, 1860) (Perciformes: Centropomidae) en agua dulce en un estanque de concreto en Alvarado, Veracruz, México. Vet. Méx. 37 (3): 327-333. [ Links ]

