











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The California Current is a highly productive eastern boundary current, spanning temperate and subtropical latitudes along the west coast of the United States and the Baja California Peninsula in Mexico. The latitudinal range of this current is dominated by a semidiurnal mixed barotropic tide and bridges the critical latitude of 30ºN for diurnal internal waves, north of which, theoretically, these waves cannot exist as freely propagating internal waves. The inner shelf of most of the California Current System is dominated by conspicuous kelp forests, with their southern range limit extending to Baja California Sur, Mexico. Although for most of the California Current System upwelling provides sufficient nutrients for primary producers year round, other mechanisms of sub-thermocline water transport, such as internal waves, have recently been shown to be important in the Mexican part of the California Current System (northern Baja California) (Ladah et al. 2012) and may be particularly important for kelp forests at their southern limit of distribution in the Northern Hemisphere off Baja California Sur.
The transport of sub-thermocline nutrients by internal waves may be an alternative mechanism for the provision of nutrients in seasons or in regions of reduced upwelling. Internal waves can force vertical and horizontal transport of many scalars, such as particles, larvae, sub-thermocline water, and nutrients for primary producers (Franks 1997, Ladah et al. 2012, Salas-Monreal et al. 2012), and are prevalent features where flows encounter sharp bathymetry in stratified waters (Holloway 1987, Lamb 1997). As internal waves propagate shoreward and touch the bottom, they can increase in amplitude and become nonlinear internal waves (NLIWs), causing rapid physical and chemical changes in shallow water (Franks 1992, Helfrich and Pineda 2003, Helfrich and Melville 2006) and strong diapycnal mixing, with dissipation rates reaching 0.2 W·m-2 over a short distance (Sandstrom and Elliot 1984, 2011).
The supply of inorganic nutrients has been directly linked with NLIWs (Sandstrom and Elliot 1984; Leichter et al. 1996, 2003; Sharples et al. 2001, 2009), which can account for a diapycnal flux of nitrate greater than 3 mmol·m-2·d-1 (Sharples et al. 2007, Schafstall et al. 2010), up to 25% of the annual nitrate contribution on the shelf in the California Current off Santa Barbara (McPhee-Shaw et al. 2007), and increased primary productivity on the inner shelf of the California Current off Southern California (Lucas et al. 2011). The only site to date where internal waves have been explored in the Mexican part of the California Current System is Todos Santos Bay in Northern Baja California (Filonov et al. 2014), where macroalgal tissue nitrogen doubled in response to semidiurnal internal wave nutrient pulses within a few hours (Pérez-Mayorga et al. 2011, Ladah et al. 2012). However, there are no in situ measurements of internal wave activity in any other site of the California Current system off Mexico, particularly in the more subtropical zone where mesoscale wind-induced upwelling becomes infrequent and less intense (Kurzyn et al. 2012) and nutrient provision by internal waves may be even more important for primary producers.
In San Juanico, Baja California Sur, Apel and Gonzalez (1983) documented internal waves using satellite imagery. This site is very near the southern limit of all kelp species in the Northern Hemisphere. Here, perennial individuals of the southern sea palm, Eisenia arborea (now known as Ecklonia arborea), thrive even when sea surface temperature reaches 26 ºC, which is presumably a nutrient-poor condition (Ladah 2003). Although the ability of kelps to survive high temperatures depends on multiple factors, survival is closely tied to tissue nitrogen concentrations (Gerard 1982, 1997) and ambient nitrogen availability (Gerard 1984; Zimmerman and Kremer 1984, 1986). Ecklonia arborea shows a significant reduction in tissue nitrogen to below 1% dry weight in about 20 d when cultured in nutrient-poor outdoor tanks or when exposed to nutrient-poor warm temperatures (Matson and Edwards 2007, Sánchez-Barredo et al. 2011). Therefore, the detection of E. arborea thriving in warm waters, with tissue nitrogen as high as 2.5%, in summer off San Juanico suggests that these kelps are exposed to a nutrient source even during warm sea surface temperatures.
In this study, we show that diurnal internal waves dominate at our study site and discuss the forcing mechanisms and potential importance for the ecology of the inner shelf of the California Current System off Baja California Sur, where upwelling is reduced in late summer and fall compared with other sites along the California Current. In this contribution, we investigate the potential factors controlling variability in high-frequency temperature records collected in the summer and fall seasons of the weak 2004-2005 El Niño event. The motivation for this study arises from observations of healthy kelp beds with high tissue nitrogen (above 2.5% dry weight) during warm summer conditions. We propose that nutrient pulses associated with diurnal internal waves may be important to the summer survival of kelp beds in this region and speculate on their forcing. This study highlights the fact that the southern part of the California Current System is a biogeographic transition zone and suggests that mechanisms that may not be particularly important in one region may be highly relevant in another, considering the great latitudinal span of the California Current.
Materials and methods
Study site
San Juanico (26.30285ºN, 112.62340ºW) is located in one of the largest and shallowest areas along the California Current, in the Gulf of Ulloa, at the southern end of the California Current System, on the Pacific coast of Baja California Sur. This area is very near the southern limit of the kelp E. arborea in the Northern Hemisphere (Fig. 1). This site is characterized by strong local winds, a very wide, gently sloping shelf, and dominant semidiurnal tides ranging from -0.26 to 2.24 m.

Figure 1 (a) Regional map of Baja California with a light grey bar showing the position of the 200-m isobath. (b) Inset showing the study area at San Juanico and the location of the instrument moorings. Sites A and B indicate thermistor mooring lines and kelp beds at 8 and 15 m depth; Site C corresponds to the offshore thermistor line moored at 20 m depth. (c) Inset showing vertical distributions of temperature (T), salinity (S), and buoyancy frequency (N) near mooring line C.
Water column temperature and nutrients
Temperature loggers sampling every 5 min (0.2 ºC accuracy, approximately 5 min response time; Onset Computer Corp., MA, USA) were installed on vertical mooring lines deployed at the 8-, 15-, and 20-m bathymetric contours (see Fig. 1b). The first 2 mooring lines (8 and 15 m) were adjacent to perennial E. arborea kelp beds, and the 20-m mooring line was 3 km offshore. Loggers were installed every meter on the shallowest mooring and every 3 m on the deeper moorings.
Water column hydrography was measured with vertical casts using a SeaBird Electronics 19 Conductivity-Temperature-Depth (CTD) profiler on 14-16 August and 2-6 September 2004 along an offshore transect encompass-ing the thermistor lines (see Fig. 1). Water samples were collected using 1.2-L Niskin bottles at the surface, mid-water, and 1 m above the CTD at its maximum depth; they were then filtered (GFF, 0.7 μm), frozen, and analyzed in triplicate for dissolved inorganic nitrogen using an autoanalyzer with minimal detectable limits at 0.2 μmol·L-1. Nitrate and nitrite were graphed against temperature (Zentara and Kamykowski 1977) to examine the temperature-nutrient relationship for this area in summer (Ladah 2003).
Wind and tide data
Hourly wind data were available from a meteorological station maintained by the Comisión Federal de Electricidad (Federal Electricity Commission, Mexico), 14 km SE from the study site. Hourly tidal information was available from San Carlos (160 km south; 24º47 N, 112º07 W).
Analysis of temperature and wind data
The vertical displacement of isotherms was calculated from the temperature time
series by using the equation (Filonov and Konyaev
2003) where ξ(t) is the vertical
displacement at a given depth z at time t and
Using 27 CTD casts from 14-16 August, the distribution of the average buoyancy frequency (Brunt-Väisälä frequency) had a minimum period of 2.7 min, at 13 m depth, with the average period for the 5-25 m depths of 6 min (Fig. 1c). Because thermistors sampled every 5 min (a Nyquist period of 10 min) it was not possible to resolve events on the order of minutes. The temperature time series were therefore smoothed with a 1-h sliding cosine filter.
Hourly wind data were analyzed for variability of the zonal (East-West) and meridional (North-South) wind components. For the wind time-series we applied rotational spectral analysis and cross spectral methods (Gonella 1972, Konyaev 1990, Emery and Thomson 1997). Squared coherence was calculated between temperature measured at all depths on mooring line A (within the shallow kelp bed) and (1) the dominant East-West component of the wind (24 h) or (2) the O1 (25.82 h) and М2 (12.42 h) tidal sea level. We calculated the spectra of the temperature time series from 6 depths at thermistor mooring lines A, B, and C and for hourly sea level fluctuations of the same length (43 d; 1,022 hourly values).
Results
The temperature time series shows daily fluctuations reaching 5 ºС (giving a calculated vertical displacement using equation [1] of approximately 18 m) in mid-September (Fig. 2). The maximum water temperature was recorded on 10 September, when it exceeded 28 ºC at the surface (Fig. 2 a, b). Cooler water (<20 ºC) episodically intruded into the kelp beds and reduced bottom temperature up to 5 ºC in an hour during the study period (Fig. 2 c-e).
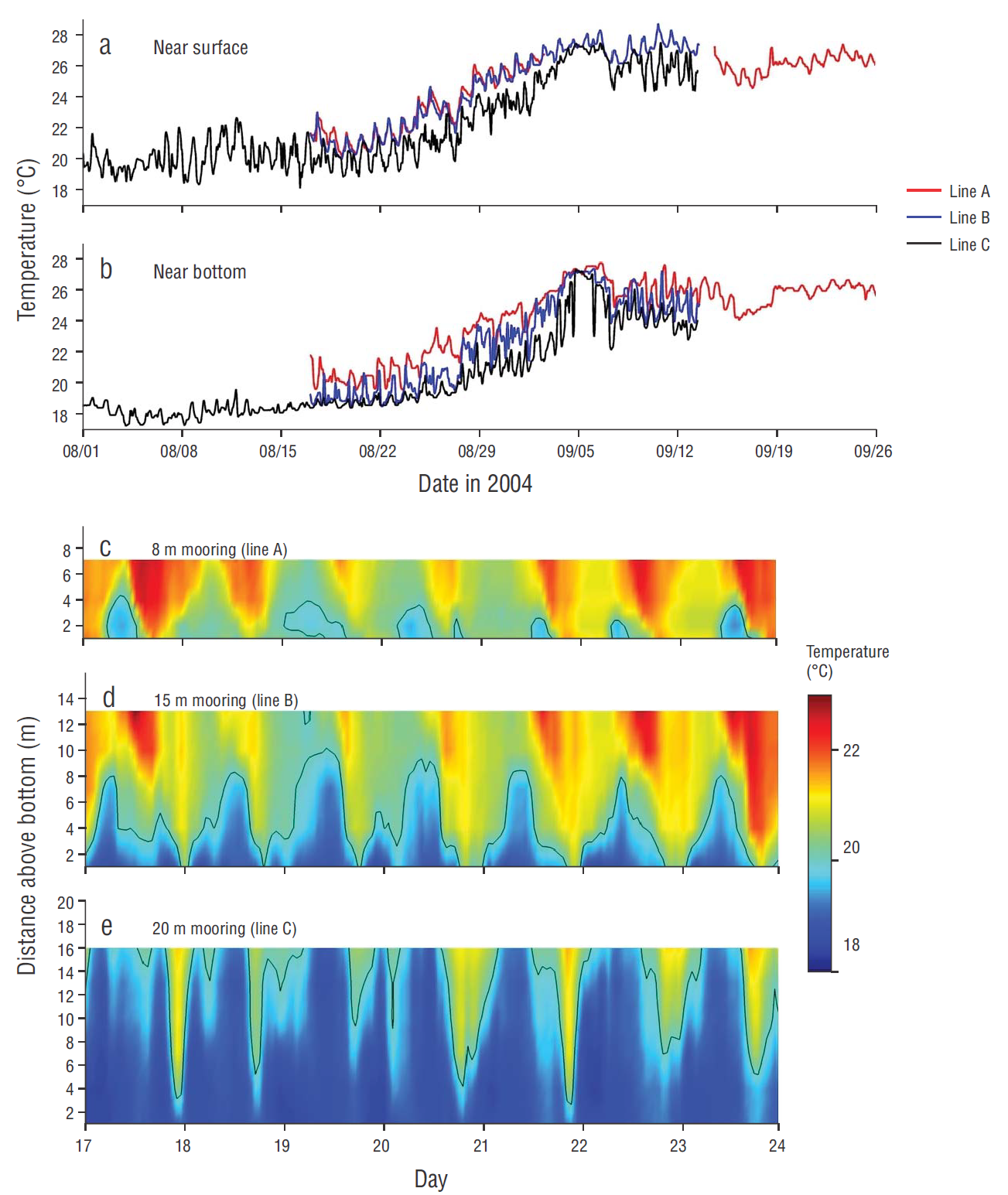
Figure 2 (a) Near-surface temperatures in San Juanico at mooring line A (8 m depth, logger 7 m above bottom), line B (15 m depth, logger 13 m above bottom), and line C (3 km offshore line at 20 m depth, logger 16 m above the bottom) (see map in Fig. 1). (b) Bottom temperatures (logger at 1 m above bottom) for mooring lines A, B, and C; and zoom of water column temperatures on days 17 to 24 of August for the (c) 8 m mooring (site A), (d) 15 m mooring (site B), and (e) 20 m mooring (site C). Black line in zoomed water column panels denotes the 19.7 ºC isotherm, determined to be the nutricline during this period of time.
Water sampled at 19.7 ºC contained measurable nitrate plus nitrite (1.96 μM), with an average of 5.45 ± 4.05 μmol·L-1 between 19.2 and 19.7 ºC. Nitrate concentrations up to 1.4 μmol·L-1, in waters of almost 20 ºC, were found in this sub-thermocline water entering the kelp beds, which is a much greater concentration of nitrate at this temperature than is usually observed for the California Current.
After a month of bottom temperatures with a mean of 23.6 ºC at the base of the 8-m bed and 22.6 ºC at the base of the 15-m bed, blades of E. arborea had relatively high tissue nitrogen (2.37% ± 0.19 at 8 m, 2.40% ± 0.30 at 15 m, maximum of 2.85% at 8 m). As nutrient-limited kelp tissue has around 1% tissue nitrogen, the value measured suggests kelp tissue was not nitrogen limited at this time. Percent nitrogen, carbon (32.39% ± 1.49, 31.67% ± 1.68), or hydrogen (3.97% ± 0.28, 4.16% ± 0.27) did not vary significantly between 8 and 15 m (P > 0.15, Student’s t test, n = 10).
The daytime sea breeze had maximum wind velocities (8-10 m·s-1) at 4 PM directed to the east, with a weaker reverse night breeze (2-3 m·s-1) (Fig. 3a). At all moorings, rising temperatures occurred throughout the day as the onshore sea breeze caused accumulation of warm surface water at the coast, depressing the thermocline (Figs. 3, 4). This phase lasted from 9 AM to 4 PM, after which time a phase of thermocline erosion occurred, with night cooling and offshore winds. The formation and erosion of the thermocline had the same phase as the zonal wind component. The intensity of daily temperature fluctuations decreased from mooring line A to lines B and C (Fig. 3b-d). At line C (Fig. 3d), the offshore mooring, the phase accumulation of heat in the surface layers, and the timing of thermocline depression were slightly lagged compared to the coastal mooring. The hourly-averaged temperature fluctuations (Fig. 3b-d) showed a defined daily periodicity coherent with the zonal wind component (Fig. 3a) (Table 1) but were not correlated to sea level fluctuations (Fig. 3e) (Table 1).
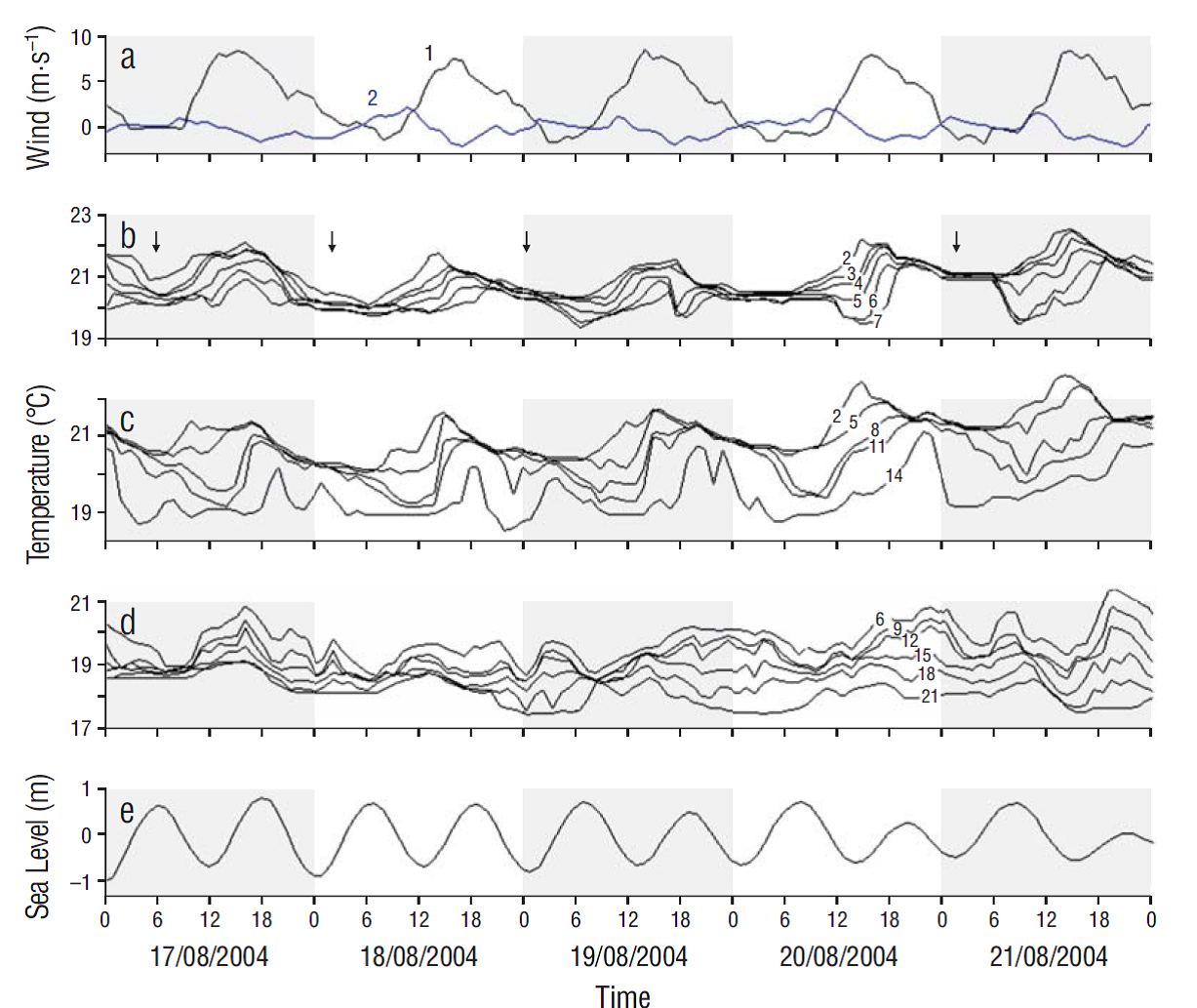
Figure 3 (а) Hourly variability of the zonal (1) and meridian (2) wind components at San Juanico. (b-d) Hourly temperature fluctuations in San Juanico at different depths on the 3 moorings (lines A, B, and C), measured on 17-21 August, and (e) predicted sea level fluctuations in San Carlos, Baja California Sur, located 160 km to the south of San Juanico. Vertical arrows in (b) show the times with maximum vertical mixing near line A.
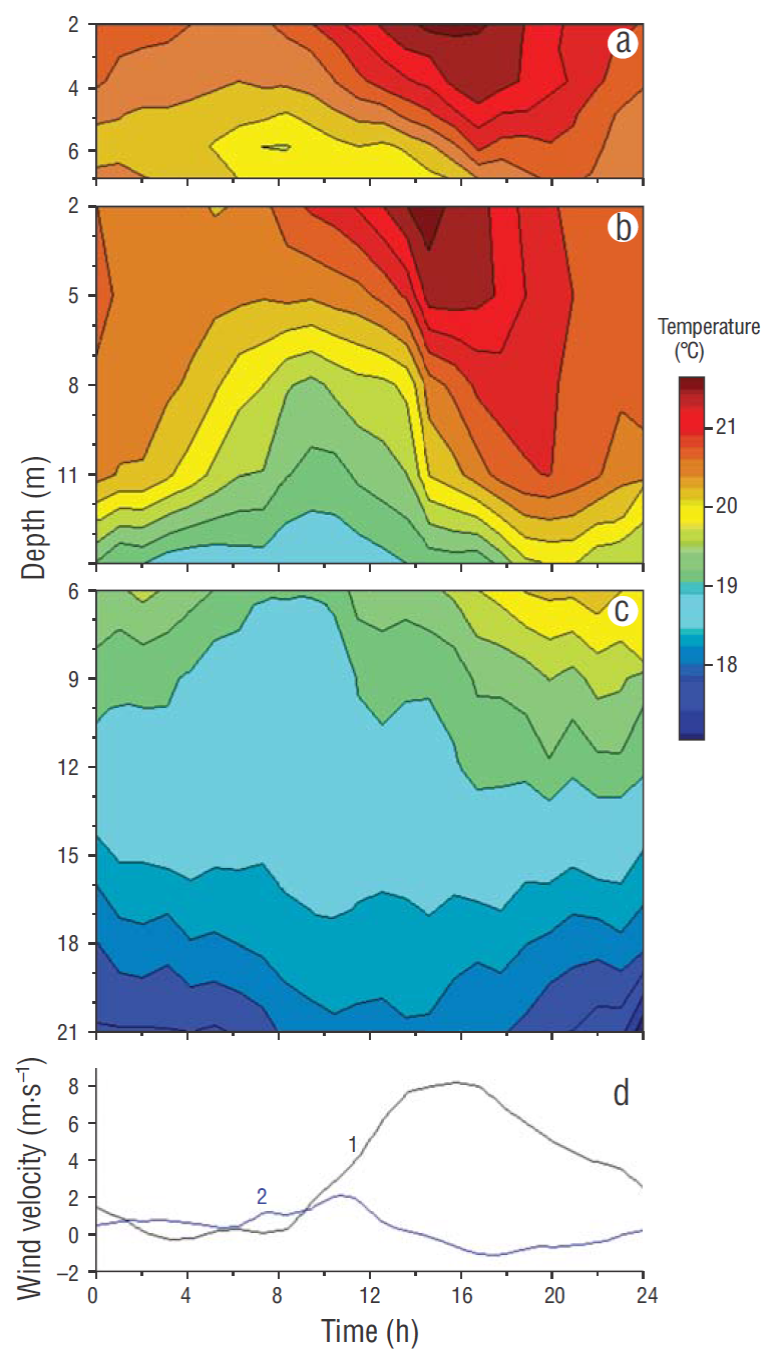
Figure 4 (a-c) Five-day average (17-21 August 2004) of temperature fluctuations at 3 moorings (a, 8 m; b, 15 m; c, 20 m) in San Juanico bay and (d) zonal (1) and meridian (2) wind components.
Table 1 Squared coherence values between the dominant E-W (U) component of the wind and temperature fluctuations, and between the sea level and temperature fluctuations. Calculations are shown for thermistors on mooring line A, coastal kelp bed, 8 m depth. Values above the 95% confidence interval (0.521) are shown in bold.
| Thermistors (height above the bottom, m) | |||||||
| Period (h) | variable | 2 | 3 | 4 | 5 | 6 | 7 |
| 12.42 M2 | Sea level | 0.200 | 0.238 | 0.343 | 0.387 | 0.324 | 0.337 |
| 25.82 O1 | Sea level | 0.167 | 0.289 | 0.439 | 0.334 | 0.177 | 0.278 |
| 24.09 | Wind | 0.772 | 0.833 | 0.835 | 0.865 | 0.814 | 0.774 |
Cotidal maps for the 4 basic tidal waves, O1 (25.8 h), К1 (23.9 h), М2 (12.4 h), and S2 (12.0 h), for the western coast of the Baja California Peninsula (Godin et al. 1980) show that the amplitude of the semidiurnal tide is almost twice that of the diurnal tide for this area (Fig. 5). The reverse was found for the spectrum of mooring temperature fluctuations, however, where the diurnal peak was one order of magnitude greater than the semidiurnal peak, suggesting that daily water temperature fluctuations near the coast were not caused by the tide (Fig. 5).
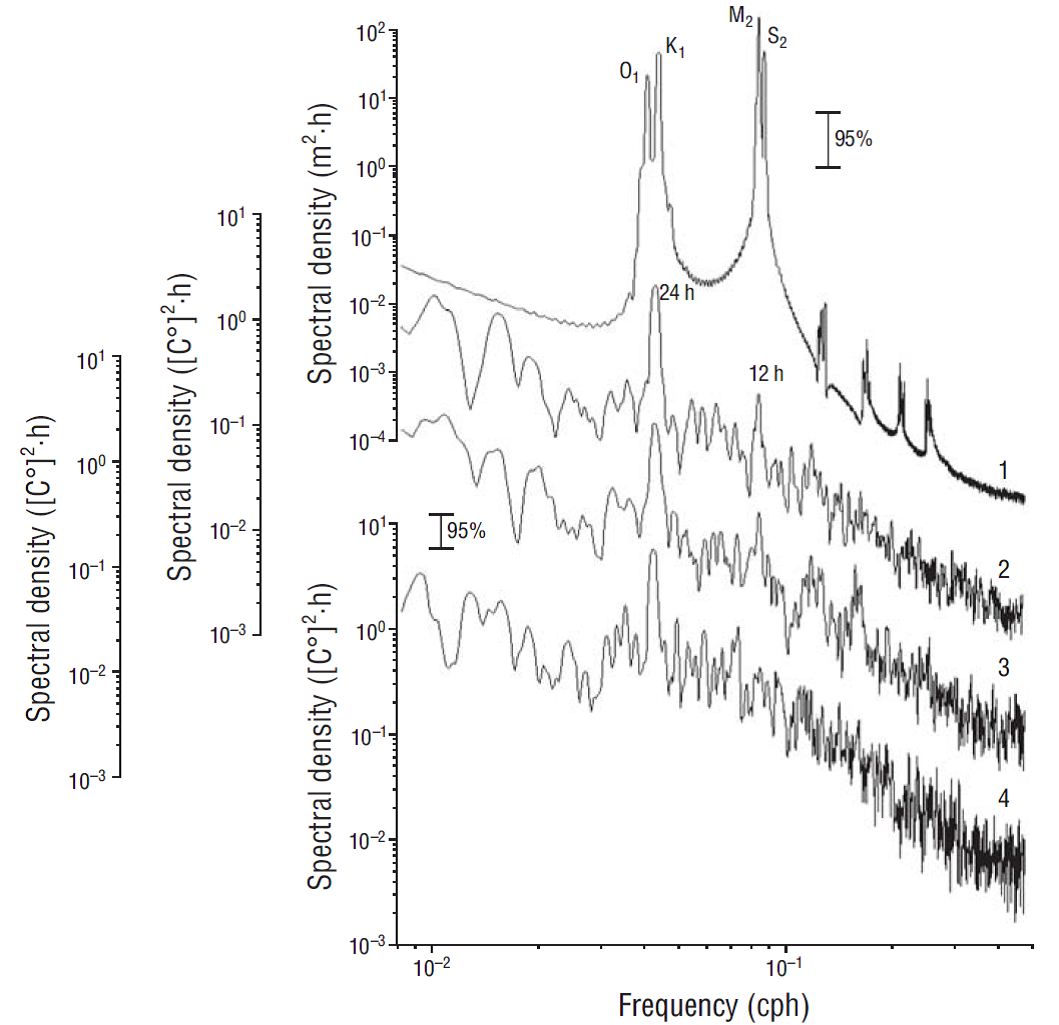
Figure 5 Spectral density of sea level fluctuations in San Carlos (1) and average spectral density of temperature fluctuations at mooring lines A (2), B (3), and C (4), calculated for temperature data collected at all measurement depths. Vertical error bar indicates the 95% confidence interval for the power spectral density function.
The squared coherence between the temperature variability at all depths on mooring line A and the dominant East- West component of the wind was almost twice as large as the 95% confidence level (CI) in the diurnal period (Table 1). For the deeper offshore mooring (line C), only the surface layers (top 7 m of the water column) were significantly coherent with diurnal wind energy above the 95% CI and dropped to near zero (0.034) at depth. In the area of mooring A (300 m from the shore, in approximately 7 m depth), the phase lag between the maximum wind speed and the depression of the thermocline down to the bottom was about 4 h. In the area of mooring B (2.5 km from the shore, at a depth of approximately 14 m), this lag was almost 6 h. There was no significant coherence between temperature and the semidiurnal (12.42 h) tide (Fig. 6, Table 1). This suggests that local diurnal winds cause the observed temperature fluctuations in the kelp beds, in contrast to others sites off Baja California, where semidiurnal tidal forcing has been shown to be more important (Ladah et al. 2012, Filonov et al. 2014).
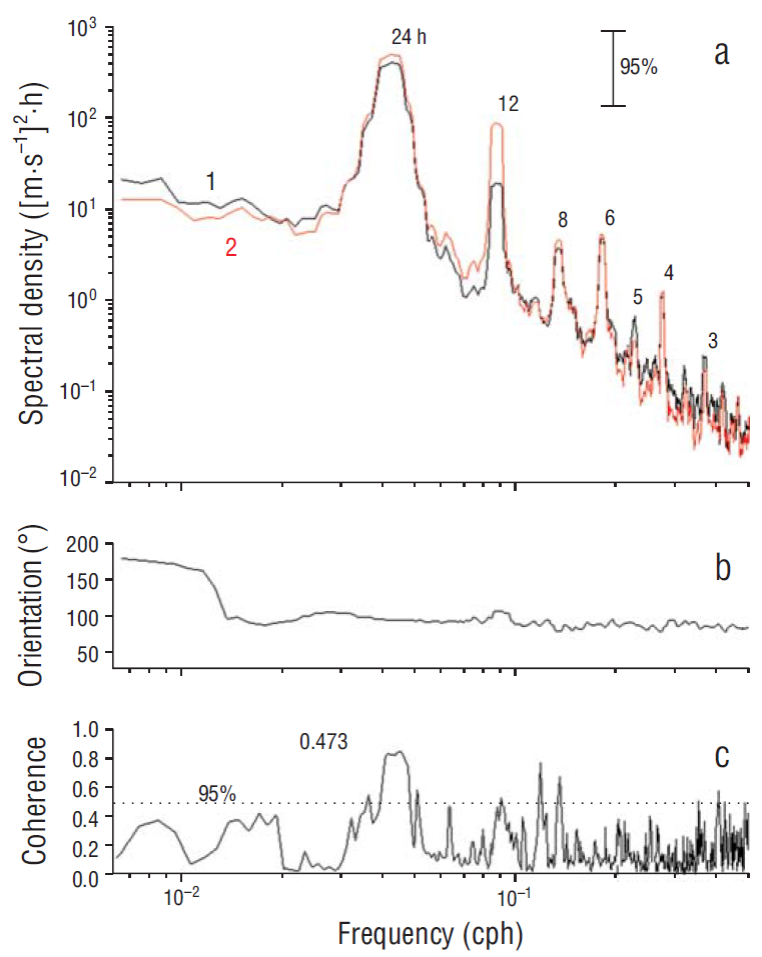
Figure 6 (a) Clockwise (1) and counterclockwise (2) rotational spectra of wind variability at San Juanico. Vertical error bar indicates the 95% confidence interval for the power spectral density function. (b) Main axis orientation for the elliptical orbits, calculated in degrees from North clockwise. (c) Coherence between temperature variability at the surface at mooring line A and variability of the wind energy (sum of the zonal and meridional wind components). Dotted vertical line indicates the 95% confidence interval for the squared coherence.
Dscussion
Kelp beds are common in the California Current, but reach their southern limit in the southern part of this ecosystem in areas off Baja California Sur, where upwelling becomes intermittent. The relatively high tissue nitrogen concentration in the kelp, E. arborea (up to 2.85%), measured after a month of warm water in summer (bottom temperature mean of 23 ºC) in San Juanico, along with previous data showing the kelp species E. arborea responds to low nitrogen conditions within 20 d both in tank cultures and in the field (Matson and Edwards 2007, Sánchez-Barredo et al. 2011), suggests a recent source of external inorganic nitrogen reached the kelp beds. The measurement of nitrate and nitrite in water as warm as 19.7 ºC, the short diurnal intrusions of water cooler than 19 ºC into the kelp beds even when sea surface temperatures were high, and the coherence of intrusions with daily winds suggest that diurnal internal waves forced by winds may transport nutrients to these kelp beds during warm periods.
Coastal nitrate enrichment at 18.5 ºC has been previously shown for the California Current off Baja California in Tortugas Bay (Hernández-Carmona et al. 2001), and an analysis of older California Cooperative Oceanic Fisheries Investigations data has suggested that anomalous warm-water nitrate enrichment might occur during El Niño events (Ladah 2003). However, in the present study, nitrate was detected at even higher temperatures (at almost 20 ºC) at a site further south in the California Current than previously analyzed. In a previous study, Ladah (2003) showed a latitudinal increase in nitrate enrichment (with 19.5 ºC water having 1.1 μM nitrate, at a site 100 km to the north of San Juanico) in warm waters during El Niño and speculated this was due to the shoaling of subtropical subsurface water. The present study also occurred during an El Niño event (2004- 2005), further supporting the idea that nutrient-rich water can shoal during El Niño in Baja California Sur. However, the persistence, seasonality, and source of enrichment have still not been identified.
Diurnal temperature changes were detected within the kelp beds during this study and showed significant coherence with the diurnal frequency in winds. Although diurnal internal waves are not unprecedented in the California Current and have been previously documented in California (Lerczak et al. 2001, Pidgeon and Winant 2005, Beckenbach and Terrill 2008, Cudaback and McPhee-Shaw 2009), we expected to find stronger semidiurnal variability in our temperature time series similar to what has been found for other sites in Mexico. The southern subtropical part of the California Current encompasses latitudes where theory predicts that unforced, freely propagating, diurnal internal waves can exist, as they are not subinertial at these latitudes. However, off the west coast of Mexico, the barotropic tide is a mixed semidiurnal tide, often resulting in a much larger variance at the semidiurnal frequency in nearshore water column temperatures than at the diurnal frequency (Filonov and Trasviña 2000; Filonov and Konyaev 2003, 2006; Filonov and Novotryasov 2005; Ladah et al. 2005; Filonov et al. 2014). Because San Juanico is located near the edge of a long shallow shelf in the Gulf of Ulloa (200 m isobath, see Fig. 1), where internal tides could be generated (Baines 1982, Holloway 1991, Konyaev and Sabinin 1992) approximately 120-150 km from the coast-line, we expected tidally forced, semidiurnal internal waves to reach the coast, with wave lengths in the range of hundreds of meters and frequencies close to the maximum buoyancy frequency (periods of 3-15 min). We did not expect winds to have a greater forcing effect than the semidiurnal tide, certainly not at the base of the kelp beds, as most areas studied to date in the Mexican Pacific show a strong semidiurnal internal tide, except very far inside Todos Santos Bay (Filonov et al. 2014), where weak diurnal internal motions occur trapped to the coast.
We propose that in San Juanico, semidiurnal internal waves are generated offshore, but dissipate on the long shallow shelf before reaching the kelp beds inshore, where wind-forced diurnal internal waves become more prevalent. Because mesoscale upwelling of nutrient-rich water can be weak and intermittent in summer off Baja California Sur (Kurzyn et al. 2012), the transport of nutrients into shallow zones by the daily thermocline shoaling measured in this study may be particularly relevant for southern-limit kelp beds. Although the percentage of overall nutrient flow to the system that diurnal internal waves can provide to this site is currently unknown, this topic merits further study as a mechanism to sustain primary productivity, particularly during upwelling-limited summers and El Niño years.

