










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencias marinas
versión impresa ISSN 0185-3880
Cienc. mar vol.36 no.3 Ensenada sep. 2010
Artículos
La profundidad e hidrodinámica como herramientas para la selección de espacios acuícolas en la zona costera*
Depth and hydrodynamics as tools to select aquaculture areas in the coastal zone**
OE Delgado–González1*, JA Jiménez2, JL Fermán–Almada3, F Marván–Gargollo1, A Mejía–Trejo1, Z García–Esquivel1
1 Instituto de Investigaciones Oceanológicas, Universidad Autónoma de Baja California, Km 106 Carretera Tijuana–Ensenada, Ensenada, Baja California, México. * E–mail: odelgado@uabc.mx
2 Laboratori d'Enginyeria Marítima, ETSECCPB, Universitat Politécnica de Catalunya, c/ Jordi Girona 1–3, Campus Nord edifici D1, 08034 Barcelona, Spain.
3 Facultad de Ciencias Marinas, Universidad Autónoma de Baja California, Km 106 Carretera Tijuana–Ensenada, Ensenada, Baja California, México.
Received December 2009
Accepted July 2010
RESUMEN
Se construyó un índice que permite calificar espacialmente las zonas de cultivo ostrícola en cuanto a su potencialidad productiva en Bahía Falsa, San Quintín (Baja California, México). El índice incorpora las dos principales variables limitantes para el desarrollo del ostión: disponibilidad de alimento y profundidad. La disponibilidad de alimento se caracterizó con base en la distribución final de la concentración de materia orgánica particulada, la cual es tratada como un material pasivo que es transportado por el campo de velocidades asociado con la marea. Se obtuvo un balance a partir del volumen incorporado a la bahía con la marea entrante y el consumo realizado por los organismos distribuidos en las zonas de cultivo, los cuales se caracterizan a través del porcentaje de área dentro de la bahía dedicada a los cultivos. En cuanto a la profundidad, se obtuvo su distribución de área de acuerdo a una clasificación de cinco intervalos que permitió identificar una mayor o menor exposición de los cultivos al alimento disponible propuesto. Ambos criterios se integraron en un valor único que permitió zonificar la bahía en parcelas en términos relativos, indicando cuál de ellas es mejor para el cultivo del ostión. Dadas las características de la zona y los volúmenes de producción, la disponibilidad de alimento no resultó ser un factor limitante, siendo el principal factor que controla la calidad de una parcela la profundidad a la que se desarrolla el cultivo. Al comparar la valoración obtenida con el tiempo de crecimiento de los ostiones cultivados, se encontró correspondencia entre la mayor valoración y el menor tiempo de crecimiento y viceversa.
Palabras clave: Crassostrea gigas, ostiones, acuicultura, San Quintín, zona costera.
ABSTRACT
An index was created for the spatial classification of oyster cultivation areas in terms of their potential productivity in Falsa Bay, San Quintín (Baja California, Mexico). The index included two main variables crucial for oyster development: food availability and depth. Food availability was based on the final distribution of particulate organic material; it is a passive variable that is transported by tidal currents. A balance was obtained between the volume incorporated into the bay by tidal inflow and its consumption by the organisms distributed in the cultivated areas, which are characterized by the percentage of cultivated areas within the bay. The distribution area of depth was obtained from a five–interval classification scale that allowed the identification of the highest and least exposure of the cultivation area to the proposed food availability. Both criteria were combined into one value in order to divide the bay region into zones in relative terms and determine which are the best suited for oyster cultivation. Given the characteristics of the area and production volume, food availability did not appear to be a limiting factor, and culture depth was the main factor controlling the zonal quality. A comparison of the index values and cultivated oyster growth rates showed that the higher index values corresponded to faster growth rates and vice versa.
Key words: Crassostrea gigas, oysters, aquaculture, San Quintín, coastal zone.
INTRODUCCIÓN
De las actividades que se desarrollan en la zona costera, la acuicultura es la que probablemente más ha crecido (FAO 2009). En el contexto mundial existe un crecimiento anual promedio de 8.8%; sin embargo, en las regiones acuícolas de América Latina y el Caribe, este crecimiento es de 22%, debido a la inexistencia de esta actividad durante el periodo de 1950 a 1970 (FAO 2009).
En México, aproximadamente el 2% de los 1.9 × 106 km2 que comprende el territorio nacional corresponde a los espacios costeros protegidos o semiprotegidos (Ortiz–Pérez y Lanza–Espino 2006) distribuidos a lo largo de los más de 11,000 km de litoral. Sin embargo, el marco normativo para promover el desarrollo acuícola con fines comerciales en esta zona quedó establecido a principios de los años noventa y actualmente esta actividad se encuentra en una etapa incipiente en la mayor parte de sus costas.
Bahía San Quintín (BSQ) es uno de los espacios protegidos en el litoral mexicano que ha experimentado el uso continuo de suelo con fines acuícolas por más de 30 años. En esta zona existen cultivos extensivos del ostión Crassostrea gigas y se emplea la técnica de estantes. Esta técnica de cultivo es similar a la utilizada por la mayor parte de los productores y aun así es posible encontrar diferencias superiores a dos meses en el tiempo que tardan los organismos cultivados en alcanzar la talla comercial en algunos puntos dentro de la laguna (García–Esquivel et al. 2000).
Los principales factores ambientales asociados con la producción de bivalvos son la temperatura, la salinidad, la profundidad y la disponibilidad de alimento; sin embargo, las características de cada región determinan su importancia y resaltan la particularidad con la que deben ser analizados otros factores ambientales (Héral y Deslous–Paoli 1991).
En este contexto, el objetivo de este trabajo es desarrollar un índice que valore la viabilidad de ambientes lagunares para la explotación acuícola de bivalvos y aplicarlo en BSQ. Aunque el índice se desarrolla para BSQ, tanto la metodología seguida como el índice en sí mismo son aplicables a otros ambientes lagunares costeros para valorar su viabilidad acuícola con bivalvos.
MATERIALES Y MÉTODOS
Área de estudio
Bahía San Quintín es una laguna costera con una extensión aproximada de 42 km2 localizada en la zona semiárida noroeste de las costas de la península de Baja California, México (fig. 1). El intercambio de agua que tiene BSQ con el Océano Pacífico es permanente a través de su única boca y el efecto de la marea mantiene la mayor parte del cuerpo de agua interior bien mezclado en sus gradientes verticales de propiedades disueltas (Millán–Núñez et al. 1982). El canal principal se divide en dos brazos: el brazo oeste, conocido como Bahía Falsa, y el brazo este, conocido como Bahía San Quintín (fig. 1). El agua que inunda BSQ tienen el efecto de las aguas oceánicas de surgencias, principalmente durante primavera y verano (Hernández–Ayón et al. 2004).
En BSQ se realizaron estudios orientados a establecer su viabilidad para realizar actividades acuícolas durante los años setenta (Chávez–de–Nishikawa y Álvarez–Borrego 1974, Lara–Lara y Álvarez–Borrego 1975, Del–Valle–Lucero 1979, Monreal–Gómez 1980, Plascencia–Díaz 1980, Álvarez–Borrego y Álvarez–Borrego 1982, Martori–Oxamendi 1989). Estos estudios permitieron obtener los primeros datos biofísicos, lográndose entre otros avances identificar que el sistema está gobernado por la marea, la cual es mixta con predominancia semidiurna. Se han registrado intensidades de corrientes de marea de 1 m s–1 durante el reflujo en la boca y desfases en tiempo de 40 min de la componente semidiurna de la onda de marea entrante entre la boca y la región de la cabeza en el brazo este (Ocampo–Torres 1980).
Bahía San Quintín está relativamente aislada de centros urbanos importantes y de descargas residuales e industriales, lo que ha permitido mantener en buenas condiciones su hábitat natural. Esta característica y la forma de realizar las actividades acuícolas que se desarrollan en su interior han despertado interés y son numerosos los esfuerzos orientados a caracterizar diferentes aspectos biofísicos en BSQ (Aguirre–Muñoz et al. 2001; Camacho–Ibar et al. 2003; Ward et al. 2003, 2004; García–Esquivel et al. 2004; Hernández–Ayón et al. 2004, 2007; Angulo–Larios 2006; Lara–Espinoza 2007; Aveytua–Alcázar 2008; Aveytua–Alcázar et al. 2008; Valle–Levinson et al. 2009). Esta información se sintetizó para lograr la construcción del índice que se describe a continuación.
Desarollo del índice biofísico
Se construyó el índice biofísico (BI) para tener una medida del potencial que posee Bahía Falsa (BF) para su explotación acuícola con ostiones. El BI valora dos variables clave que controlan la cría de ostiones: la disponibilidad de alimento y la profundidad a la que se encuentran los cultivos. Para la construcción del BI se requiere caracterizar la distribución espacial de los flujos de agua y de la profundidad en BF durante las posibles condiciones de marea y viento. Dado que entre 90% y 95% de la varianza de las corrientes medidas en superficie y fondo en BF está explicada por la marea (Angulo–Larios 2006, Flores–Vidal 2006), en este trabajo se optó por incorporar sólo la componente de marea. Una vez que se caracterizaron los flujos de agua, se asociaron con la disponibilidad de alimento (IPOMd), en este caso caracterizado con los valores de las concentraciones de la materia orgánica particulada (POM, mg L–1) reportados para BF (García–Esquivel et al. 2004) y el intervalo de profundidad normalizada (DRn):
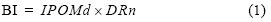
La distribución espacial de los flujos en BF está determinada principalmente por el estado de la marea. Estos flujos incorporarán el alimento a la bahía y varían a medida que cambia la carrera de marea. Para usar el intervalo de posibles intensidades del flujo, se tomó una condición de marea viva y otra de marea muerta. Así, se tomaron en cuenta los siguientes supuestos: (a) la marea llenante asociada con la pleamar de la marea viva representa la situación en la que se incorpora un mayor volumen de agua a la bahía y las propiedades biofísicas pasivas en el agua entrante se pueden detectar en los lugares más alejados de la boca, y (b) el volumen de agua oceánica que introduce el flujo de marea entrante está relacionado con la diferencia entre la altura subsiguiente y el tiempo que tarda en cubrir esa diferencia. Con base en esto, se realizó un análisis a partir de las predicciones de marea del año 2009 (Centro de Investigación Científica y de Educación Superior de Ensenada [CICESE]) y se eligieron la marea viva del 10 de enero de 2009, en el periodo de 2:00 a 8:00, y la marea muerta de las 20:00 del día 14 de junio a las 2:00 del día 15 de junio de 2009 como condiciones de los casos utilizados para caracterizar la distribución espacial de los flujos en BF (fig. 2).
Para caracterizar el flujo asociado, se utilizó el modelo hidrodinámico MIKE 21 (DHI 2008), el cual resuelve las ecuaciones de Navier–Stokes (conservación de momentum) y la ecuación de continuidad (conservación de masa) en dos dimensiones (integrado en la vertical) mediante un esquema de diferencias finitas implícito. El modelo permite la simulación de áreas sumergidas y expuestas, lo cual activa las celdas que tengan una profundidad mínima (que para la zona de estudio se tomó como 7 cm), y desactiva las celdas cuya profundidad es inferior a un valor determinado. Esta capacidad es especialmente relevante en un sistema como BSQ, donde el área sumergida en marea baja es significativamente menor que el área sumergida en marea alta. El modelo se implementó usando una malla rectangular con tamaño de 100 m y se forzó utilizando series temporales de la marea en la boca de la laguna. Para calibrar el modelo se utilizaron series de tiempo de velocidad de corrientes en marea viva y marea muerta en dos puntos internos de la BF, las cuales fueron medidas mediante un perfilador de corrientes ADCP (Acoustic Doppler Current Profiler).
Para el análisis espacial, se dividió BF en una serie de elementos o cajas (fig. 3) utilizando como criterios la morfología y la orientación del campo de velocidades de pleamar de marea viva llenante obtenido con el modelo MIKE 21 (fig. 4). La orientación del campo permitió definir tres ejes longitudinales a lo largo de BF: un eje delimitó el cauce del canal secundario en el costado este, el segundo la parte central de BF por la convergencia observada del campo de velocidades y el último, el más corto, delimitó un espacio con movimiento tipo giro en la parte noroeste de BF. El criterio morfológico se utilizó para seleccionar el límite de la frontera sur de BF, y éste consistió en incorporar la mayor área posible trazando una línea de la barra de arena hacía la región costera central, cuidando no incorporar la zona del canal principal. Se obtuvo así un seccionamiento de cinco franjas horizontales y tres verticales, cuyos cruces definen 15 cajas, enumeradas en orden creciente partiendo del límite suroeste de la frontera sur de BF.

En cada caja se ubicaron los estantes identificados en trabajo de campo. En términos generales, cada estante está constituido por 120 sartas y cada sarta tiene un número variable de individuos; sin embargo, se reconoce que una buena cosecha se obtiene cuando cada sarta posee tres docenas de individuos, cantidad que se eligió para calcular el número de ostiones por caja control. Se calculó un total aproximado de 37.6 millones de individuos para BF al extrapolar el número de ostiones por caja control y el número de estantes reportados por los concesionarios del espacio acuícola en BF (Delgado–González 2010).
La profundidad de la zona es un factor determinante al utilizar estantes en el cultivo de ostión. Si los estantes se colocan a una profundidad de 1.5 m referido al nivel medio del mar, profundidad que coincide con la altura aproximada que tienen los estantes desde el fondo, cada vez que baje la marea se descubrirá una parte o toda la estructura con los ostiones, limitando la posibilidad de que éstos consigan alimento sólo en los tiempos de inmersión. Considerando que el máximo descenso de marea es alrededor de 1.4 m por debajo del nivel medio del mar, los estantes instalados en aquellas zonas de la planicie en BF con una profundidad mayor a 2.5 m permanecerán sumergidos la mayor parte del tiempo, permitiendo un mayor acceso al alimento para los ostiones de los cultivos. Con base en los datos de la profundidad con relación al nivel medio del mar, se definieron los siguientes intervalos de profundidad: 0–1, 1.01–1.5, 1.51–2, 2.01–2.5 y 2.51–3.0 m. Para cada una de las 15 cajas se obtuvo el porcentaje de área en cada intervalo de profundidad (DR):
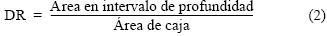
Con el propósito de resaltar la importancia relativa que tienen entre sí los cinco intervalos de profundidad propuestos en el área para el cultivo de ostión en BF, y dado que el intervalo con menor profundidad es el menos recomendado para reducir el tiempo de crecimiento del ostión empleando la técnica de estantes, se asoció un valor numérico unitario a cada intervalo de profundidad: 1 para el intervalo de menor profundidad (0.0 a –1.0 m) y 5 para el de mayor profundidad (–2.51 a –3.0 m). Este valor numérico se multiplicó por el porcentaje de área encontrado en cada caja (ecuación 2) y se normalizó con respecto al mayor valor numérico encontrado entre las cajas para determinar el DRn.
Para analizar la distribución de POM en BF se utilizaron los valores mínimo y máximo (0.7 ± 0.1 y 2.5 ± 0.2 mg L–1) reportados por García–Esquivel et al. (2004), quienes evaluaron las tasas de ingestión y biodepositación de C. gigas en BF utilizando mediciones de 6 a 8 h de duración, en cuatro ocasiones diferentes, y obtuvieron tasas de ingestión de 2.6 ± 0.4 a 3.9 ± 0.3 mg h–1. En el presente trabajo, la hipótesis para establecer la forma en que POM se distribuye entre las cajas contempló el comportamiento pasivo de su distribución y, en consecuencia, que su distribución dependerá de las condiciones hidrodinámicas. Un porcentaje de esta fracción orgánica es consumida por los ostiones presentes en la caja correspondiente, otro porcentaje sale hacia las zonas contiguas al norte, este u oeste y el resto se retiene con el volumen que eleva el nivel de agua en la caja al subir la marea. La tasa de ingestión neta que reportan García–Esquivel et al. (2004) es para organismos de C. gigas con talla comercial (93 mm) y esta tasa la encontraron incluso en condiciones con mayores concentraciones de POM. Esta habilidad de mantener tasas de ingestión similares, independientemente de la concentración de alimento disponible, lo atribuyen a la plasticidad fisiológica de C. gigas, de ahí que la tasa de ingestión utilizada para estimar la distribución de POM haya sido 3.9 ± 0.3 mg h–1.
La cantidad de POM que ingresa a cada caja (POM in) se obtuvo multiplicando el volumen que entra durante las 6 h de marea (Vin ) por la fracción orgánica del seston (POM):

El consumo (Con) se obtuvo al multiplicar el número de ostiones (Os) que se encuentran en cada caja por la ingestión (Ing = 0.023 g 6 h–1) que cada individuo realiza (García–Esquivel et al. 2004):

El volumen que se retiene y hace subir el nivel del agua en cada caja (Va) tiene asociada una fracción de POMin. Esta fracción (POMa) es la cantidad de POM retenido y resulta de obtener la diferencia entre POMin y Con, multiplicada por la fracción entre Va y Vin:

donde Va es la diferencia entre Vin y el volumen que sale (Vout). Si en la caja no hay ostiones, la cantidad de POM retenido será el producto de POMin por la fracción de Va y Vin.
La cantidad de POM que sale de una caja (POMout) es la diferencia de POMin y el Con, multiplicada por la fracción de volumen que sale (Vout) con respecto al que entra en la caja (Vin):

La relación por caja de POMa con respecto a Va es el indicador de la distribución de alimento (IPOMd) y permite tener una idea de los cambios de POMin y, por tanto, del efecto que tienen los organismos conforme la POM se distribuye en el área de cultivo. El IPOMd toma en cuenta la hidrodinámica, la ingestión que realizan el total de los ostiones durante las 6 h de marea llenante (aproximadamente 0.9 t si se usa la razón de consumo de 3.9 mg L–1) y la cantidad que se retiene en la caja formando parte del volumen que hace que suba el nivel de agua en la caja:
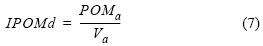
Con la distribución de alimento para cada caja (ecuación 7) se obtuvo BI (ecuación 1), expresión que puede reescribirse realizando algunas sustituciones algebraicas con las expresiones encontradas anteriormente para adquirir la siguiente forma:
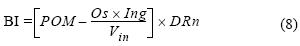
Esta expresión revela que BI está en función de una componente que relaciona el alimento y organismos contabilizados, y otra que incorpora la profundidad normalizada.
RESULTADOS
El campo de velocidades que se obtuvo al modelar el flujo de la marea alta entrante seleccionado permitió observar la participación del delta interno de marea en reducir la velocidad del agua que pasa por su parte menos profunda y mantener mayores velocidades del agua que fluye por los canales que definen los límites este y oeste del delta (fig. 4). El volumen de agua que inunda la planicie de BF sigue los contornos del fondo y se distinguen las siguientes características principales: a través del canal este fluye el agua que más logra penetrar en la zona de la cabeza de BF; el canal oeste incorpora agua con velocidad similar al canal este, mas sólo alcanza la parte media de BF, debido a la poca profundidad con que discurre su cauce; los canales este y oeste tienen ramificaciones de menores dimensiones que distribuyen el flujo de agua hacia la parte central de BF, formando en este encuentro de parcelas de agua una zona de convergencia; y la predominancia del canal este en la incorporación de agua sobre el oeste genera una zona de circulación, formando un giro en la segunda mitad del costado oeste de BF (fig. 4).
Los flujos de la marea entrante incorporan el principal volumen de agua a BF por las cajas 1 y 3, las cuales enmarcan el inicio de los canales secundarios. La caja 2 recibe mayor volumen por su frontera este que por su frontera sur debido a la menor profundidad en esta frontera, tal como se puede apreciar por la batimetría mostrada en la figura 1. Esta batimetría permite identificar la zona de bajos en la caja 2.
Los intercambios de volumen entre las cajas se obtuvieron a partir de los resultados del modelo hidrodinámico. Por ejemplo, en la caja 1 entran 4.67 × 106 m3 por la frontera sur y 0.14 × 106 m3 por la frontera este, mientras que por la frontera norte salen 2.29 × 106 m3 y por la frontera este salen 0.27 × 106 m3. La diferencia entre Vin y Vout representa el volumen que se retiene y eleva el nivel de agua en la caja: Va = 2.25 × 106 m3. Los volúmenes respectivos para cada una de las cajas se presentan en la tabla 1. La distribución de POM, principal constituyente de la dieta de los ostiones, se calculó a partir de estos valores.
Al combinar la condición de marea muerta con un valor de POM de 0.7 mg L–1, la IPOMd obtenida bajo este escenario (fig. 5) puede considerse como el escenario menos favorable, dada la forma en que disminuye la concentración de alimento desde su ingreso en las primeras tres cajas hasta llegar a la cabeza de BF (caja 15; tabla 2). El consumo en siete de las cajas es del mismo orden de magnitud que el valor obtenido por IPOMd.
Cuando se consideró la condición de marea viva, al incluir una concentración de POM de 2.5 mg L–1 (fig. 5), las concentraciones iniciales de POM fueron de hasta un orden de magnitud mayor que el consumo en todas las cajas y en algunas pudo ser de hasta dos órdenes mayor (tabla 2). Los valores de IPOMd se incrementaron significativamente con respecto a la situación de marea muerta, indicando una mayor disponibilidad de alimento y excediendo el consumo esperado.
La distribución espacial de DRn en BF (fig. 6) muestra que las cajas 5, 8 y 9 presentan las mayores proporciones de profundidad propuestas como favorables para el cultivo y las cajas 3 y 11 las proporciones más bajas y menos favorables. En la caja 3 las pendientes del cauce del canal este determinan la baja viabilidad de cultivo, mientras que en la caja 11 lo determina la escasa profundidad (fig. 6). Es conveniente señalar que las profundidades del canal este (cajas 3 y 6) se encuentran dentro del intervalo de 2 a 3 m; sin embargo, éstas profundidades forman parte de la pendiente del cauce y son inapropiadas para colocar los estantes, lo cual explica el porcentaje de cero en algunas cajas (tabla 3).

La normalización de BI con respecto al mayor valor encontrado en las cajas durante marea viva permite identificar su distribución espacial. Teniendo en cuenta que la concentración de alimento utilizada para marea viva fue mayor que la concentración utilizada para marea muerta, esta normalización reposiciona la diferencia de concentraciones. Se observa cómo los mejores sitios de cultivo en marea muerta se ubican en el lado oeste (cajas 1, 4 y 5; fig. 7), mientras que en mareas vivas se ubican en la parte central (cajas 5, 8 y 9). La escala de valores (fig. 7) en marea viva es mayor que en marea muerta, lo cual puede interpretarse como mejores condiciones para toda BF.
DISCUSIÓN
La aproximación utilizada para evaluar el potencial acuícola de BF empleando el volumen asociado a la componente hidrodinámica inducida por la marea está directamente relacionada con el alimento que se incorpora y con la elevación del nivel de agua en su interior. La hidrodinámica obtenida empleando el modelo numérico (MIKE 21) permitió distinguir el papel de los canales en el llenado de BF en condiciones de una marea viva y una muerta. En la condición de la marea muerta seleccionada el canal que discurre por el lado oeste de BF (caja 1) incorpora mayor volumen que el canal este (caja 3), mientras que en la condición de marea viva ocurre lo contrario (tabla 1, fig. 4).
La concentración inicial de POM asociada al volumen inicial y el efecto que tiene la distribución e ingestión de organismos fueron factores determinantes para la calificación del espacio acuícola de este trabajo (ecuación 8). Aunque el número y la disposición de las cajas podrían modificarse, es de esperar que la distribución espacial de POM sea similar a la obtenida empleando las 15 cajas y el modelo hidrodinámico, ya que la concentración POMa es hasta dos órdenes de magnitud mayor que el consumo en el escenario de condiciones de marea viva, cuya concentración inicial fue de 2.5 mg L–1.
El consumo de POM en cada caja reflejó la densidad de organismos cultivados por metro cúbico. En las zonas de mayor densidad (cajas 4 y 5) los organismos cultivados consumirían 30% del total de POM en BF (tabla 2). Sin embargo, de la POM incorporada durante la marea muerta (3.45 t), el consumo total que realizarían los 37.6 millones de ostiones cultivados, a razón de 0.023 g 6 h–1, sería de 0.9 t, es decir, se consumiría solamente 26% del alimento disponible, distribuyéndose las 2.5 t restantes en el cuerpo de agua. Este porcentaje de consumo se reduciría a 3% si se considera el escenario de marea viva, en el cual se introducen aproximadamente 36 t. Este último porcentaje es del orden de magnitud de los valores reportados por Banas et al. (2006), quienes dan valores de consumo entre 8% y 15% del fitoplancton neto incorporado por la marea. Estas estimaciones permiten sugerir que aun bajo las condiciones menos favorables asociadas con marea muerta y la concentración inicial utilizada, la cantidad indicada de ostiones en cultivo dispondría de alimento suficiente para soportar al cambio de marea viva.
García–Esquivel et al. (2004) estimaron que la capacidad de carga ostrícola de BF se encuentra entre 1913 y 5979 t, considerando individuos con peso promedio de 52.2 g; las cantidades representan aproximadamente 36 y 114 millones de ostiones, respectivamente. Mientras que el consumo realizado por los 36 millones de ostiones es similar al estimado para la cantidad de ostiones en cultivo y, por tanto, dispondrían de suficiente alimento en condiciones de marea muerta, los 114 millones de individuos consumirían aproximadamente 2.6 t, es decir, 75% de la concentración que introduce esta marea, un porcentaje comprometedor para lograr el cultivo y la subsistencia de otras especies que ocupan el mismo espacio y que compiten por el mismo tipo de alimento. Sin embargo, la condición de marea viva con una concentración inicial de 2.5 mg L–1 incorpora aproximadamente 36 t, y el consumo que realizarían los más de 100 millones de organismos representa sólo 7% de esta cantidad que se incorpora, lo cual refleja la importancia de la combinación del estado de la marea con las concentraciones de POM.
Para las condiciones de marea muerta y viva, los valores de IPOMd disminuyeron hacia la cabeza de BF (tabla 2, fig. 5). Las cajas 1, 2, 3, 6 y 10 mantuvieron valores similares a las concentraciones iniciales, las tres últimas principalmente por estar ubicadas en el cauce del canal este, en el cual se dificulta la colocación y mantenimiento de estantes. Independientemente de esta distribución se debe tomar en cuenta que para valores absolutos la diferencia de una celda a otra es pequeña y, en cualquier caso, excede los valores mínimos requeridos. Esta no limitación por disponibilidad de alimento hace que el factor profundidad aumente su importancia relativa en cuanto a la definición de microregiones productivas, de tal forma que podría considerarse como el factor predominante en las condiciones actuales.
De acuerdo con el DRn, al comparar las cajas, las condiciones de ocho cajas disponen de menos de la mitad de las características de la caja mejor calificada. Las cajas que se encuentran en la segunda y la tercera franja (4, 5, 7, 8 y 9), así como la caja 12 en la cuarta franja, son las mejor evaluadas. La zona que define la tercera franja dispone de mayor área con condiciones apropiadas para el cultivo empleando la técnica de estantes, mientras que las zonas definidas por la primera, cuarta y quinta franjas presentan menos área proporcional (fig. 6). La incorporación de DRn permitió comparar las características que tiene cada caja y reconocer que existen diferencias que deben ser consideradas en el uso del espacio para volver más eficiente las prácticas del cultivo; por ejemplo, buscar un equilibrio que permita reducir la altura de los estantes pero que siga siendo redituable su mantenimiento o cambiar la técnica de cultivo.
La diferencia en tiempo mostrada en las curvas de crecimiento por organismos localizados en la cabeza y boca (García–Esquivel et al. 2000) corresponden a la clasificación de BI de las cajas 15 y 5, cuyos valores son de 0.02 y 0.17, respectivamente, en condición de marea muerta, y 0.36 y 0.88, respectivamente, en marea viva (fig. 7). Mientras que la distribución del alimento y la profundidad en la condición de marea muerta contribuyen a los valores bajos de BI, en marea viva la profundidad determina la valoración dados los valores de IPOMd, lo cual sugiere que la profundidad de instalación es el factor que caracteriza o determina las microregiones de BF señaladas por estos autores.
Además del papel que determina la hidrodinámica y el filtrado de los bivalvos cultivados en la distribución del fitoplancton en BF, habría que considerar el posible efecto ocasionado por la presencia de las granjas. Estas granjas pueden funcionar como barreras y alterar localmente la circulación de agua, lo cual podría generar acumulación de materia orgánica en los espacios con mayor cantidad de estantes, como lo sugiere la distribución de lodos en la parte central de BF (Daesslé et al. 2009), o distribuciones espaciales particulares de fitoplancton dentro de la zona de cultivos, de acuerdo con la posición, orientación y profundidad del cultivo (Gillibrand y Turrell 1997). Se esperaría que aquellos cultivos próximos al canal principal y a los secundarios, colocados en planicies con profundidades de 2.5 m o más, estarían en mejores condiciones de disponibilidad de alimento, ya que estarían expuestos a los flujos generados por la marea entrante por más tiempo que aquellos alejados de éste y con profundidad menor a la indicada. Sin embargo, se desconocen los efectos particulares que tienen los cultivos en la distribución natural sedimentos y del alimento, situación que tendrá que ser atendida con mediciones y modelos hidrodinámicos con mayor capacidad espacial resolutiva.
Las cinco cajas con mejor clasificación BI, fueron las cajas 4, 5, 8, 9 y 12 (fig. 7). Las primeras cuatro coinciden con los espacios que albergan las concesiones con mayor número de estantes instalados. Los mejores valores de BI en estas áreas de cultivo, en principio, pueden atribuirse a la profundidad, que permite mantener los estantes bajo el agua la mayor cantidad del tiempo, así como a la posición relativa que guardan los estantes con respecto a los cauces de los canales a través de los cuales fluyen los principales flujos de marea y con ello el alimento nuevo que se incorpora del Pacífico.
La aplicación del índice para valorar la viabilidad de ambientes lagunares aquí desarrollado y aplicado en BF, indica que para este ambiente lagunar, las diferencias observadas en el tiempo que tardan los organismos en alcanzar su talla comercial de 0.09 m están asociadas con el tiempo de exposición impuesto por la profundidad del área de cultivo, dado que la disponibilidad de alimento estimada con escenarios de marea muerta y marea viva supera las exigencias alimenticias. El producto del indicador de la distribución de alimento con el de profundidad permite obtener un índice que caracteriza el estado biofísico para el desarrollo acuícola del ostión de forma diferenciada a lo largo de la BF, con lo cual se logran identificar las zonas óptimas para la explotación (fig. 7) pudiendo ser utilizada para planificar desarrollos futuros.
AGRADECIMIENTOS
El primer autor agradece el apoyo económico brindado por CONACYT. A la UABC el apoyo económico recibido a través de la decimocuarta convocatoria interna a proyectos de investigación. A los revisores anónimos por las sugerencias y comentarios emitidos al presente trabajo.
REFERENCIAS
Aguirre–Muñoz A, Buddemeier RW, Camacho–Ibar V, Carriquiry JD, Ibarra–Obando SE, Massey BW, Smith S, Wulff F. 2001. Sustainability of coastal resource use in San Quintín, Mexico. Ambio 30: 142–149. [ Links ]
Álvarez–Borrego J, Álvarez–Borrego S. 1982. Temporal and spatial variability of temperature in two coastal lagoons. CalCOFI Rep. 23: 188–197. [ Links ]
Angulo–Larios NT. 2006. Hidrodinámica de la Bahía de San Quintín, Baja California. M.Sc. thesis, Universidad Autónoma de Baja California, Mexico, 128 pp. [ Links ]
Aveytua–Alcázar L. 2008. Modelación de la dinámica biogeoquímica en una laguna costera. Ph.D. thesis, Universidad Autónoma de Baja California, Mexico, 219 pp. [ Links ]
Aveytua–Alcázar L, Camacho–Ibar VF, Souza AJ, Allen JI, Torres R. 2008. Modelling Zostera marina and Ulva spp. in a coastal lagoon. Ecol. Model. 218: 354–366. [ Links ]
Banas NS, Hickey BM, Newton JA, Ruesink JL. 2007. Tidal exchange, bivalve grazing, and patterns of primary production in Willapa Bay, Washington, USA. Mar. Ecol. Prog. Ser. 341: 123–139. [ Links ]
Camacho–Ibar V, Carriquiry JD, Smith S. 2003. Non–conservative P and N fluxes and net ecosystem production in San Quintín Bay, Mexico. Estuaries 26: 1220–1237. [ Links ]
Chávez–de–Nishikawa AG, Álvarez–Borrego S. 1974. Hidrología de la Bahía de San Quintín, Baja California, en invierno y primavera. Cienc. Mar. 1: 31–62. [ Links ]
Daesslé LW, Rendón–Márquez G, Camacho–Ibar VF, Gutiérrez–Galindo EA, Shumilin E, Ortiz–Campos E. 2009. Geochemistry of modern sediments from San Quintín coastal lagoon, Baja California: Implication for provenance. Rev. Mex. Cienc. Geol. 26: 117–132. [ Links ]
Del–Valle–Lucero I. 1979. Aplicaciones de un modelo numérico y análisis de condiciones hidrodinámicas en Bahía San Quintín, BC, México. M.Sc. thesis, Centro de Investigación Científica y de Educación Superior de Ensenada, Mexico, 36 pp. [ Links ]
Delgado–González O. 2010. Desarrollo y aplicación de una herramienta de gestión para el aprovechamiento acuícola en Bahía San Quintín, Baja California. Ph.D. thesis, Universidad Politécnica de Cataluña, España. 182 pp. [ Links ]
DHI. 2008. Hydrodinamic module (HD), http://www.dhigroup.com. [ Links ]
FAO. 2009. The State of World Fisheries and Aquaculture 2008. FAO Fisheries and Aquaculture Department, Rome, 107 pp. [ Links ]
Flores–Vidal X. 2006. Circulación residual en Bahía San Quintín, BC, México. M.Sc. thesis, Centro de Investigación Científica y de Educación Superior de Ensenada, México, 91 pp. [ Links ]
García–Esquivel Z, González–Gómez MA, Ley–Lou F, Mejía–Trejo A. 2000. Microgeographic differences in growth, mortality, and biochemical composition of Pacific oysters (Crassostrea gigas) from San Quintin Bay, Mexico. J. Shellfish Res. 19: 789–797. [ Links ]
García–Esquivel Z, González–Gómez MA, Ley–Lou F, Mejía–Trejo A. 2004. Oyster culture potential in the west arm of San Quintin Bay: Current biomass and preliminary estimate of the carrying capacity. Cienc. Mar. 30: 61–74. [ Links ]
Gillibrand PA, Turrell WR. 1997. The use of simple models in the regulation of the impact of fish farms on water quality in Scottish sea lochs. Aquaculture 159: 33–46. [ Links ]
Héral M, Deslous–Paoli JM. 1991. Oyster culture in European countries. In: Menzel W (ed.), Estuarine and Marine Bivalve Mollusk Culture. CRC Press, Florida, pp. 154–186. [ Links ]
Hernández–Ayón M, Galindo–Bect MS, Camacho–Ibar VF, García–Esquivel Z, González–Gómez MA, Ley–Lou F. 2004. Nutrient dynamics in the west arm of San Quintín Bay, Baja California, Mexico, during and after El Niño 1997/98. Cienc. Mar. 30: 119–132. [ Links ]
Hernández–Ayón JM, Camacho–Ibar VF, Mejía–Trejo A, Cabello–Pasini A. 2007. Variabilidad del CO2 total durante eventos de surgencia en Bahía San Quintín, Baja California, México. In: Hernández de la Torre B, Gaxiola–Castro G (eds.), Carbono en Ecosistemas Acuáticos de México. DF SEMARNAT–INE–CICESE, Mexico, pp. 187–200. [ Links ]
Lara–Espinoza A. 2007. Variabilidad espacial de alcalinidad total durante eventos de surgencia en Bahía San Quintín, Baja California, México. B.Sc. thesis, Universidad Autónoma de Baja California, Mexico, 65 pp. [ Links ]
Lara–Lara JR, Álvarez–Borrego S. 1975. Ciclo anual de clorofilas y producción orgánica primara en Bahía San Quintín, Baja California. Cienc. Mar. 2: 77–97. [ Links ]
Martori–Oxamendi JI. 1989. Variabilidad de la circulación y sus causas en Bahía San Quintín, BC. M.Sc. thesis, Centro de Investigación Científica y de Educación Superior de Ensenada, Mexico, 110 pp. [ Links ]
Millán–Núñez R, Álvarez–Borrego S, Nelson DM. 1982. Effects of physical phenomena on the distribution of nutrients and phytoplankton productivity in a coastal lagoon. Estuar. Coast. Shelf Sci. 15: 317–335. [ Links ]
Monreal–Gómez MA. 1980. Aplicaciones de un modelo de dispersión en Bahía San Quintín, Baja California, México. M.Sc. thesis, Centro de Investigación Científica y Educación Superior de Ensenada, Mexico, 90 pp. [ Links ]
Ocampo–Torres FJ. 1980. Análisis y predicción de velocidad mediante un modelo unidimesional en Bahía San Quintín, BC, México. B.Sc. thesis, Universidad Autónoma de Baja California, Mexico, 90 pp. [ Links ]
Ortiz–Pérez MA, Lanza–Espino DL. 2006. Diferenciación del espacio costero de México: Un inventario regional. Serie Textos Universitarios, Instituto de Geografía, UNAM, México 137 pp. [ Links ]
Plascencia–Díaz R. 1980. Determinación de la circulación en Bahía San Quintín, BC, por gradientes de densidad. B.Sc. thesis, Universidad Autónoma de Baja California, Mexico, 123 pp. [ Links ]
Valle–Levinson A, Gutiérrez–de–Velasco G, Trasviña A, Souza AJ, Durazo R, Mehta AJ. 2009. Residual exchange flows in subtropical estuaries. Estuar. Coasts 32: 54–67. [ Links ]
Ward DH, Morton A, Tibbitts LT, Douglas DC, Carrera–González E. 2003. Long–term change in eelgrass distribution at Bahía San Quintín, Baja California, Mexico, using satellite imagery. Estuaries 26: 1529–1539. [ Links ]
Ward DH, Tibbitts TL, Morton A, Carrera–González E, Kempa R. 2004. Use of digital multispectralvideography to assess seagrass distribution in San Quintín Bay, Baja California, México. Cienc. Mar. 30: 47–60. [ Links ]
* Traducido al español por Christine Harris.
** Descargar versión bilingüe (Inglés–Español) en formato PDF.

