










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencias marinas
versión impresa ISSN 0185-3880
Cienc. mar vol.30 no.4 Ensenada dic. 2004
Artículos
Transport of benthic diatoms across the continental shelf off southern Baja California Peninsula
Transporte de diatomeas bentónicas sobre la plataforma continental en frente de la parte sur de la Península de Baja California
A. Martínez-López1*, D.A. Siqueiros-Beltrones1,2, N. Silverberg1
1 Centro Interdisciplinario de Ciencias Marinas Av. Instituto Politécnico Nacional s/n Col. Playa Palo de Santa Rita Apartado postal 592 La Paz, CP 23096, Baja California Sur, México. *E-mail: amartin@ipn.mx
2 Departamento de Biología Marina Universidad Autónoma de Baja California Sur La Paz, Baja California Sur, México.
Recibido en mayo de 2003;
aceptado en septiembre de 2003.
Abstract
Material collected with a time-series sediment trap moored in San Lázaro Basin, near the NE Pacific upwelling zone off Bahía Magdalena, Baja California Sur (Mexico), from August to November 1996 and December 1997 to June 1998, revealed temporal changes in the flux of benthic diatoms. Fluxes were highest during fall and spring. Lateral transport of benthic diatoms from the coastal zone seems to be an important phenomenon at this site. The species assemblage includes components found mainly in mangrove environments of Bahía Magdalena, as well as nearby rocky coastlines, indicating that locally the northward counter current or coastal gyres carry living cells, and possibly other particles of lagoon origin, across the continental shelf.
Key words: benthic diatoms, sediment trap, transport, Baja California Peninsula.
Resumen
Material recolectado en una trampa de sedimento de serie de tiempo anclada en la Cuenca San Lázaro, cerca de la zona de surgencias del Pacífico nororiental frente a Bahía Magdalena, Baja California Sur (México), de agosto a noviembre de 1996 y de diciembre de 1997 a 1998, reveló cambios temporales en el flujo de diatomeas bentónicas. Los mayores flujos se registraron en otoño y primavera. El transporte lateral de diatomeas bentónicas desde la zona costera parece ser un fenómeno importante en la zona. Las asociaciones de especies encontradas incluye especies que se encuentran principalmente en ambientes de manglar de Bahía Magdalena, así como en los litorales rocosos cercanos, lo que indica que la contracorriente en dirección norte o los giros costeros acarrean localmente células vivas, y posiblemente otras partículas de origen lagunar, a través de la plataforma continental.
Palabras clave: diatomeas bentónicas, trampa de sedimento, transporte, Península de Baja California.
Introduction
When identifying benthic marine diatoms one often relies on key species or floristic inventories from distant localities, some separated by unbridgeable barriers (continents). None-theless, many of the taxa are the same, at least under the morphological species concept (Mann and Kociolek, 1990), as is the case for the coasts of England, Indian Ocean and Baja California Peninsula (Siqueiros-Beltrones, 2002). This wide distribution of benthic diatom species could be explained on the basis of a dispersion biogeographic model considering basic oceanic circulation. The mechanisms of dispersion may not be clear because there is not enough oceanographic information available on a local or regional basis. However, particular environments and/or substrates contain characteristic diatom assemblages. The presence of certain taxa found in some highly productive areas, such as seagrass or mangrove ecosystems, may be used as an indicator of the influence that such systems exert on the offshore environment, which suggests the existence of some type of mechanism for dispersion.
Highly productive diatom mats on the continental shelf have been detected down to water depths of 40 m, caused by the successful establishment of benthic diatoms transported from inshore sediments. They settle and may grow well even under low light levels, possibly down to depths of 200 m (Cahoon et al., 1990).
The marine sediment record of benthic diatoms may provide evidence of displacement of shallow-water material, or when the diatoms are autochthonous, indicate the depth of the sea floor at the time of sediment deposition. The features of marine benthic diatoms that make such applications possible are: (1) in general, benthic forms are readily distinguished from planktonic ones; and (2) benthic diatoms are generally restricted to depths of less than ~100 m.
Recognition of associations that recur in similar habitats at different geographic locations is useful for the understanding of habitat preference or selection. Trap samples of particulate matter from 300 m depth in San Lázaro Basin, about 100 km offshore, showed a much greater component of benthic than planktonic diatom species. To evaluate the origin of these species and the possibility of transport from another location to the sampling site, we have undertaken the precise taxonomic identification and description of the assemblage structure of the benthic diatoms, and determined their flux variations in the area.
Materials and methods
Samples were collected using a 12-cup Technicap PPS3/3 sediment trap, at San Lázaro Basin (25°11.8' N, 112°39.9' W), a semi-isolated depression on the edge of the continental shelf, northwest of the highly productive Magdalena Bay lagoon system (fig. 1). This system includes the northernmost presence of mangroves in the northeastern Pacific (outside the Gulf of California).
The trap was deployed from August to November 1996 and from December 1997 to June 1998 at a depth of 300 m and a distance of 150 m from the bottom. Each cup had been acidcleaned and filled with NaCl-enhanced seawater (40 ppt) containing 4% buffered formalin to preserve the samples. The trap was programmed to collect samples on an 8-day basis in 1996, and a 15-day basis during the second deployment. Whole trap samples were split using a MacLaren-Honjo-style rotary splitter. Initially, from samples of a 1/16 split, an aliquot of fresh (untreated) material was placed in a small volume sedimentation chamber and examined to estimate the proportion of "live" cells (pigmentation still evident). Subsequently, the aliquot was returned to the original split, and rinsed and centrifuged in distilled water several times in order to remove salts and preservatives. The entire split was placed in a 15-mL graduated test tube and cleaned following the methods of Hasle and Fryxell (1970). In addition, selected historical phytoplankton samples from surface net tows obtained at the northern entrance (Boca de La Soledad) of Magdalena Bay were analyzed for benthic diatom species.
For the enumeration of diatoms, the acid-cleaned samples were placed in a 10-mL sedimentation chamber. After a 24-h settling period, the entire surface of the bottom of the chamber was examined using an Olympus CH-3 inverted microscope. The resulting counts were converted to daily fluxes of each diatom species, calculated by multiplying by 16 to obtain total trap numbers and dividing by the trap collecting surface and the number of days sampled. After enumeration, the samples were deposited on cover-slips and mounted in Hyrax (R.I. = 1.65). From these slides selected diatoms were photographed at 400x and 1000x magnification.
Taxonomic determinations were done using an Olympus CH-2 phase contrast microscope with mounted camera, following the classic works of Desikachary (1986-1989), Hendey (1964), Hustedt (1955, 1959, 1961-1966), Peragallo and Peragallo (1897-1908), Schmidt et al. (1874-1959); recent literature (Navarro, 1982; Foged, 1984; Round et al., 1990); and regional studies (Siqueiros-Beltrones, 2002).
Several indices were applied to the trap samples in order to describe the structure of the diatom assemblages in samples from six dates representative of the two sampling periods. Because of the scarcity of specimens all the valves present in one or two slides were counted. Planktonic forms were also enumerated.
To determine the most important taxa based on abundance and frequency the Biological Value Index was used. Diversity was measured using indices based on information theory (log 2): Shannon's H', Pielou's equitability index (J'), and the redundancy index (REDI).
Also, the similarity between samples was measured using the Jaccard and Morisita indices. The former is based solely on the presence/absence of species, while the latter also considers the abundance of each taxon.
Results and discussion
Fluxes
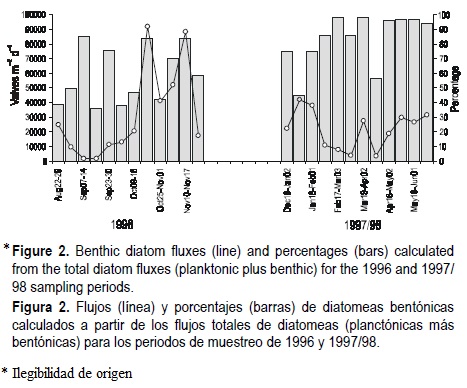
The rates of benthic diatom flux and their percentage of the total diatom flux for the two trapping periods are shown in figure 2. Diatoms showed a seasonal pattern, although the two series are not contiguous. There were maxima in the fall (late October to November 1996), of 91,000 valves m-2 d-1, and winter (late December 1997 to mid-February 1998), of 88,000 valves m-2 d-1. Higher fluxes were observed in January 1998, of 40,000 valves m-2 d-1. A minor peak was recorded during the spring of 1998. This was associated with an increase in the percentage of benthic component after late January, amounting to more than 90% during the spring (fig. 2).
Taxonomy and possible origins
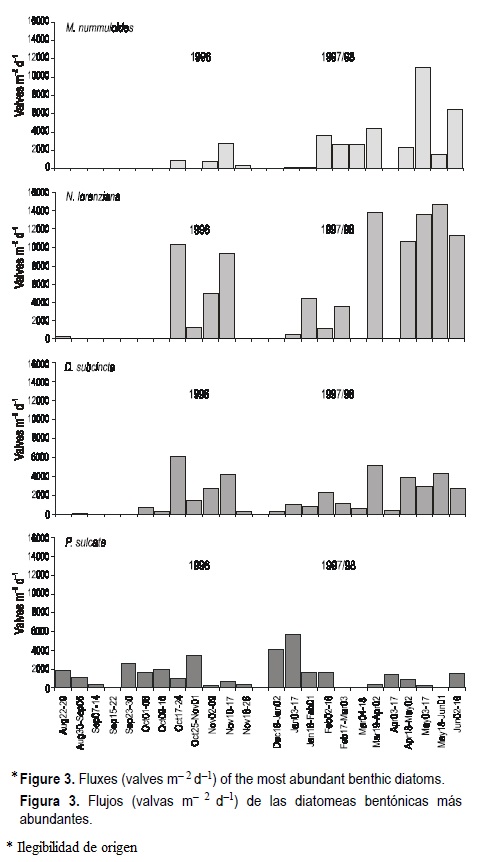
A total of 93 taxa of benthic diatoms were identified in the sediment trap material. These (and four planktonic taxa) are listed in table 1, with their corresponding authorities and type of environment in which they have been noted. Four abundant species (Melosira nummuloides Agardh, Nitzschia lorenziana Grunow, Diploneis subcincta (A.S.) Cleve, and Paralia sulcata (Ehr.) Cleve) accounted for about 74% of the total flux. The numeric fluxes of these species (fig. 3) tended individually to follow that of the total benthic diatom flux, with most displaying notable flux variations as well.
The estimated diversity values were high, similar to those of productive environments (Siqueiros-Beltrones, 1990), in spite of the scarcity of specimens in the small samples available (table 2). Although the lower values corresponded to lower species richness (S), equitability was also low in such cases. The highest H' value was observed in the September sample (4.76), along with the highest J' (0.867) and lowest redundancy (0.215), but not the highest S (45).
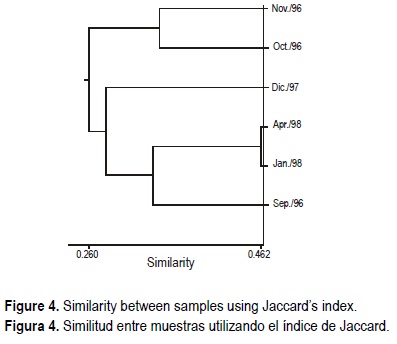
The comparison between samples showed a similar behavior with the Jaccard and Morisita indices, i.e., the grouping of the January and April 1998 samples with the highest value of similarity (fig. 4). Presence/absence was relatively low (Jac = 0.462), but considering the species abundances, Morisita's index reached a value of 1. The samples for October and November 1996 were very similar but clearly segregated from the other samples. This indicated that most of the similarity was caused by abundant taxa that differed in these two months in their order of numerical importance (table 3). Thus, the most abundant species in these two samples were Achnanthes orientalis Hustedt, Diploneis subcincta (A.S.) Cleve, Cyclotella stylorum Brightwell, Nitzschia lorenziana Grunow and Amphora ovalis Kützing, while the other assemblages were dominated mainly by Thallasionema nitzchioides Grunow (two forms), Paralia sulcata (Ehr.) Cleve, P. sulcata var. crenulata Grunow and P. sulcata var. radiata (Grunow) Peragallo & Peragallo.
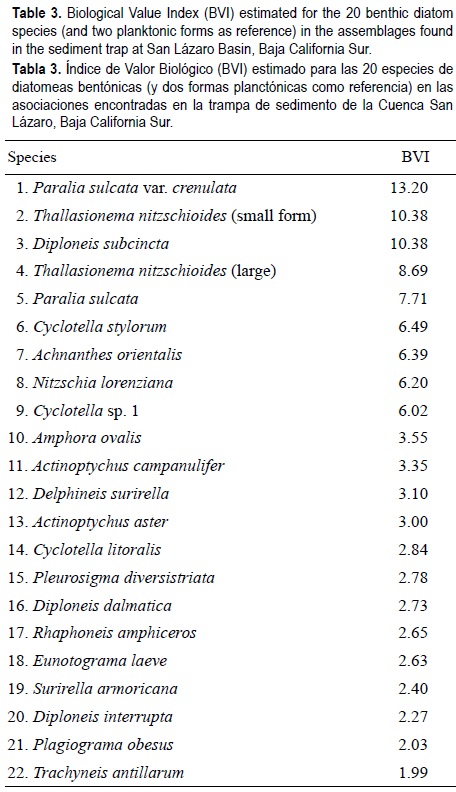
Much of the taxonomic assemblage found in the traps is similar to that found in mangrove environments located in La Paz Bay (Siqueiros-Beltrones and Morzaria-Luna, 1999; Siqueiros-Beltrones and Sánchez-Castrejón, 1999), or in other mangrove habitats (Navarro, 1982; Foged, 1984). Likewise, the H' values are as high as those estimated for benthic diatom assemblages in mangrove systems in the southern peninsula (Siqueiros-Beltrones and Sánchez-Castrejón, 1999; López-Fuerte, 2002).
The Cocconeis species in table 1, which are epilithic or epiphytic forms, are common in the rocky environment outside the Magdalena-Almejas lagoon system (Siqueiros-Beltrones, 2000). They are mostly pioneer species that adhere strongly to rocks or to macroalgae including corallines. Strong growth and little grazing may combine to allow waves and currents to dislodge some of the diatoms and parts of their substratum. A clump of Odontinium marinum Grunow was noted, partially preserved through the mounting process. In the laboratory, the formation and detachment of small clumps of other species has been observed in three-week old microalgal films growing on artificial substratum (Siqueiros-Beltrones, 1999).
Some of the more abundant taxa, such as Diploneis subcincta and D. dalmatica (plate 1), had not been collected previously in southern Baja California. Other species, such as Nitzschia capitellata Hustedt and Amphora coffeaeformis var. dussenii (Brun.) A. Cleve, had not been previously recorded in the south either. Diploneis subcincta was recorded from the Ensenada area and San Quintín Bay, in the northern part of Baja California, while D. dalmatica and Pleurosigma diversistriata Meister, along with 13 other taxa, constitute new records for the entire west coast of the peninsula (table 1). This indicates that much floristic work remains to be done for the northwestern region of Mexico.
Only 18.6% of the taxa collected in our trap (table 1) are known exclusively from mangrove environments, but 40% have been recorded from nearby mangrove and/or rocky habitats, though not in the northern (state of Baja California) part of the peninsula (Siqueiros-Beltrones, 2000, 2002; Siqueiros-Beltrones and Morzaria-Luna, 1999). About 47% of the taxa have been collected in both extremes of the peninsula, and ≈20% of the taxa are unidentified species or of uncertain habitat (new records). Although we did not propose a name for those taxa, our experience indicates that they have not been observed previously in the region (Siqueiros-Beltrones, 2002). Thus, the transitional nature of this region is well represented by the benthic diatom assemblage, containing species of temperate latitudes, and species of tropical (mangrove) affinity (plate 2).
Analysis of historical samples collected in the shallow northern opening (Boca de La Soledad) of Magdalena Bay, which is closer to San Lázaro Basin, yielded 62 taxa of benthic diatoms. Twenty-one of these taxa ocurred in the samples from the traps (table 1), including the second observation of Pleurosigma diversistriata, plus Biddulphia alternans (Bailey) Van Heurck, Cyclotella stylorum Brightwell, Cymatosira belgica Van Heurck, Eunotogramma leave Grunow, Lyrella lyra f. subtypica (A. Schmidt) comb. nov, Paralia sulcata, P. sulcata var. radiata, Trachyneis aspera (Ehr.) Cleve and T. velata Peragallo. These were recorded mainly during spring (April-June) when the maximum phytoplankton production occurs inside Magdalena Bay (Gárate-Lizárraga and Siqueiros-Beltrones, 1998). All of these species (except P. diversistriata) are commonly found on mangrove prop roots in the bay (López-Fuerte, 2002).
Observation of the fresh material from the trap samples showed that, although loose valves and fragments were also present, most of the benthic diatoms collected were alive. The additional buoyancy provided by their organic contents would have made their transport more feasible. It is still uncertain if all of the (living) benthic diatom cells are local, which would imply that this area of the coast is an important transition region, or if some are from distant localities, which would imply a great resistance over meso-scale distances. The occurrence of more temperate species this far south suggests that the California Current can apparently transport these cells to lower latitudes. On the other hand, the similarities in species composition clearly indicate a strong influence from the Magdalena-Almejas lagoon system. The occurrence of benthic diatoms in the samples from tows in the northern opening of Magdalena Bay supports our assumption that biogenic material is exported from this system.
The precise method of dispersal of the benthic diatoms so that they end up being intercepted by our sediment trap at 300 m depth is not known. Some of the valves are likely carried away from the coast and interior shallow portions of Magdalena Bay by tidal currents (Gárate-Lizárraga and Siqueiros-Beltrones, 1998), and exported to the shelf. Here they may undergo prolonged random dispersion by storm waves and shelf currents (Stanley and Swift, 1974) and simply sink permanently in the calm waters of San Lázaro Basin. Others may derive from the sandy shelf platform east of the basin. The trap was situated in a deep, broad valley tributary to the main basin, and may thus have been exposed to mass movements (such as sand cascades or less likely, turbidity currents) from the walls of the basin. It is difficult to imagine such seafloor-hugging events being responsible for carrying the fine-grained, biogenic-rich material collected to the center of the valley 150 m above the bottom.
The phenomenon discussed here is relevant to this subtropical region because the export of benthic diatoms as well as dissolved and particulate carbon and nutrients contribute to biological production in this area. This could be very important during anomalous periods of oligotrophic condition related to events like El Niño 1997-98, since it attenuates its impact.
Acknowledgements
Financial support was provided by the Dirección de Estudios de Posgrado e Investigación, Instituto Politécnico Nacional, for the research projects "Variabilidad estacional a decenal de indicadores de la productividad en el Pacífico nororiental: Un estudio combinado de la columna de agua y los sedimentos laminados" (DEPI 966592) and "Variación estacional del régimen biooceanográfico en la Cuenca San Lázaro" (DEPI 968104), and by the Mexican Consejo Nacional de Ciencia y Tecnología for the same project (CONACYT 3920P-T9608). Salaries for the authors are supported in part by Estímulos al Desempeño en Investigación (EDI fellowships) and COFFAA from the Instituto Politécnico Nacional.
References
Cahoon, L.B., Redman, R.S. and Tronzo, C.R. (1990). Benthic microalgal biomass in sediments of Onslow Bay, North Carolina. Estuar. Coast. Shelf Sci., 31: 805-816. [ Links ]
Desikachary, T.V. (1986). Atlas of Diatoms. Fasc. I, 8 p., Plates 1-77. Madras Science Foundation, India. [ Links ]
Desikachary, T.V. (1987). Atlas of Diatoms. Fasc. II, 10 p., Plates 78-221. Madras Science Foundation, India. [ Links ]
Desikachary, T.V. (1987). Atlas of Diatoms. Fasc. III, 10 p., Plates 222-400. Madras Science Foundation, India. [ Links ]
Desikachary, T.V. (1988). Atlas of Diatoms. Fasc. IV, 13 p., Plates 401-621. Madras Science Foundation, India. [ Links ]
Desikachary, T.V. (1989). Atlas of Diatoms. Fasc. V, 13 p., Plates 622-809. Madras Science Foundation, India. [ Links ]
Foged, N. (1984). Freshwater and littoral diatoms from Cuba. Bibliotheca Diatomologica, Kramer. Germany, 123 pp., 60 plates. [ Links ]
Gárate-Lizárraga, I. and Siqueiros-Beltrones, D.A. (1998). Time variation in phytoplankton assemblages in a subtropical lagoon system after the 1982-1983 El Niño event (1984 to 1986). Pac. Sci., 52, 79-97. [ Links ]
Hasle, G.R. and Fryxell, G.A. (1970). Diatoms: Cleaning and mounting for light and electron microscopy. Trans. Am. Microscop. Soc., 89 (4): 469-474. [ Links ]
Hendey, N.I. (1964). An introductory account of the smaller algae of Brittish coastal waters. Part V: Bacillarophyceae (Diatoms). Fish. Invest. Ser. IV, HMSO, London, 317 pp. [ Links ]
Hustedt, F. (1955). Marine littoral diatoms of Beaufort, North Carolina. Duke Univ. Mar. Stat. Bull., 6: 1-67. [ Links ]
Hustedt, F. (1959). Die Kieselalgen Deutschland, Osterreichs und der Schweiz. In: L. Rabenhorst (ed.), Kryptogamen-Flora von Deutschland, Osterreich und Schweiz, VII Band, II Teil. Koetz Scientific Books (Rep. 1991), Germany, 845 pp. [ Links ]
Hustedt, F. (1961-1966). Die Kieselalgen Deutschland, Osterreichs und der Schweiz. In: L. Rabenhorst (ed.), Kryptogamen-Flora von Deutschland, Osterreich und Schweiz, VII Band, III Teil. Koeltz Scientific Books (Rep. 1991), Germany, 816 pp. [ Links ]
López-Fuerte, F.O. (2002). Estructura de las asociaciones de diatomeas epífitas en raíces de Rhizophora mangle L. en Bahía Magdalena, BCS, México. B.Sc. thesis, Universidad Autónoma de Baja California Sur, Mexico, 56 pp. [ Links ]
Mann, D.G. and Kociolek, J.P. (1990). The species concept in diatoms: Report on a workshop. In: H. Simola eEd.), Proc. 10th International Diatom Symposium, Koenigstein, Koeltz. [ Links ]
Navarro, N. (1982). Marine diatoms associated with mangrove prop roots in the Indian River, Florida, USA. Bibliotheca Phycologica, 61: 1-151, 37 plates. [ Links ]
Peragallo, H. and Peragallo, M. (1897-1908). Diatomees marines de France et districts maritimes voisins. Ed. M.J. Tempere, Grez-sur-Loing, France, 492 pp. [ Links ]
Round, F.E., Crawford, R.M. and Mann, D.G. (1990). The Diatoms: Biology and Morphology of the Genera. Cambridge Univ. Press, 747 pp. [ Links ]
Schmidt, A., Schmidt, M., Fricke, F., Heiden, H., Müller, O. and Hustedt, F. (1874-1959). Atlas der Diatomaceenkunde. Reisland, Leipzig. [ Links ]
Siqueiros-Beltrones, D.A. (1990). A view of the indices used to assess species diversity in benthic diatoms associations. Cienc. Mar., 16(1): 91-99. [ Links ]
Siqueiros-Beltrones, D.A. (1999). Estructura y sucesión en asociaciones de diatomeas bentónicas utilizadas como pastura de abulones Haliotis spp. en cultivo. Ph.D thesis, Universidad Autónoma de Baja California, Ensenada, Mexico, 93 pp. [ Links ]
Siqueiros-Beltrones, D.A. (2000). Benthic diatoms associated to abalone (Haliotis), Magdalena, Baja California Sur, Mexico. Oceánides, 15(1): 35-46. [ Links ]
Siqueiros-Beltrones, D.A. (2002). Diatomeas bentónicas de la península de Baja California; diversidad y potencial ecológico. Oceánides-CICIMAR/IPN-UABCS, 102 pp. [ Links ]
Siqueiros-Beltrones, D.A. and Morzaria-Luna, H.N. (1999). New records of marine benthic diatom species for the northwestern Mexican region. Oceánides, 14(2): 89-96. [ Links ]
Siqueiros-Beltrones, D.A. and Sánchez-Castrejón, E. (1999). Structure of benthic diatom assemblages from a mangrove environment in a Mexican subtropical lagoon. Biotropica, 31(1): 48-70. [ Links ]
Stanley, D.J. and Swift, D.J.P. (1974). Marine Sediment Transport and Environmental Management. John Wiley, New York, 638 pp. [ Links ]

