











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La producción de papaya (Carica papaya) en Costa Rica representa una actividad social y económica de alto valor, con un crecimiento promedio anual de 5 % entre 2018 y 2022 (Acuña, 2023). Las principales áreas de producción se encuentran en las regiones Atlántica, del Pacífico Central y en la zona al norte del país. Casi el 75 % de la producción de papaya es vendida localmente, y el remanente es exportado. Durante el 2022, las exportaciones se realizaron a un total de 10 destinos, donde el mercado norteamericano (Canada y EE. UU.) representaron el 94 % de los embarques, mientras que el 4 % del total exportado por Costa Rica fue al mercado europeo (Acuña, 2023).
Mundialmente, este cultivo es afectado por varias enfermedades virales (Quito-Avila et al., 2023) y fitoplasmas (Fiore et al., 2018), causando pérdidas significativas. Adicionalmente, en algunos países de la región del Caribe, un organismo semejante a las rickettsias causa la enfermedad del cogollo racemoso (“papaya bunchy top”, PBT) (Davis et al., 1998). Los productores (principalmente en los países en desarrollo) no mantienen registros de los rendimientos y pérdidas de sus cultivos, ni de su asociación con enfermedades o síntomas específicos de las plantas. Es bastante común que los productores coexistan con los síntomas de enfermedades hasta que el problema es ubicuo y ponen en riesgo la sobrevivencia de las plantas. Desde 2014, los productores de papaya en Costa Rica han reportado un nuevo desorden, localmente llamado ‘pecíolo rayado’. Esta condición afecta diferentes variedades comerciales, incluyendo Maradol, Pococí, y Tainung, así como híbridos silvestres. La sintomatología se caracteriza por pecíolos curveados con distintivas estrías color purpura, acompañada de clorosis foliar y arrugamiento de la lámina. Al progresar la infección, los entrenudos se acortan y se produce una inhibición significativa del cuajado de frutos, generando un riesgo potencial a la producción local. Estos síntomas se observaron en plantaciones comerciales de papaya en las regiones Norte y Atlántica de Costa Rica. Por lo anterior, el objetivo de este trabajo fue determinar si un virus está asociado a la enfermedad con síntomas de pecíolo rayado de la papaya en Costa Rica.
Desarrollo experimental
Muestras foliares de 70 plantas maduras de papaya (8-10 meses) mostrando al menos dos de los síntomas asociados a la enfermedad del “peciolo rayado”, incluyendo el pecíolo curveado, meristemo apical con número reducido de hojas, cese del cuajado de frutos, y estrías de color púrpura en el pecíolo, fueron recolectadas (finales del 2014) en Muelle de San Carlos (provincia de Alajuela), Guácimo y Guápiles (provincia de Limón) en Costa Rica. Las plantaciones de papaya seleccionadas estaban libres de deficiencias de minerales y los suelos no mostraban signos de encharcamiento o exceso de agua. Las muestras recolectadas fueron analizadas serológicamente mediante pruebas de ELISA para detector las especies virales Potyvirus papayanuli (papaya ringspot virus, PRSV), Potexvirus papayae (papaya mosaic virus, PapMV), Orthotospovirus tomatomaculae (tomato spotted wilt virus, TSWV), Cucumovirus CMV (cucumber mosaic virus, CMV) y con anticuerpos género específico para potyvirus. Los anticuerpos específicos y los controles positivos para cada virus fueron adquiridos de Agdia Inc. (Elkhart, IN). Las pruebas se realizaron de acuerdo a los protocolos y recomendaciones de Agdia Inc.
Debido a los resultados negativos obtenidos en las pruebas de ELISA, evaluaciones mediante microscopia electrónica de transmisión (MET) se realizó para visualizar potenciales fitopatógenos. Un subgrupo de ocho muestras foliares de plantas de papaya sintomáticas (Muelle n=4, Guácimo n=2 y Guápiles n=2), y una planta sana recolectada de un jardín en el Valle Central, todas ellas negativas para los virus evaluadas mediante ELISA, fueron recolectadas (Enero, 2015) y procesadas para MET, de acuerdo a Montero-Astúa et al. (2017). Secciones de lámina y pecíolo fueron fijados toda la noche usando solución de Karnovsky amortiguada con cacodilato de sodio 0.05 M a 15 °C, y luego de cuatro lavados con amortiguador fueron post-fijadas con tetraóxido de osmio (1 %) a temperatura ambiente. Las muestras fueron deshidratadas y luego embebidas usando una serie de etanol / óxido de propileno, seguido por mezclas de óxido de propileno con resina epóxica de Spurr de dureza media, y embebido en resina pura, finalmente fue polimerizada con calor. Las secciones ultradelgadas con doble tinción (acetato de uranilo acuoso y citrato de plomo de Sato) fueron observadas usando un microscopio electrónico Hitachi H-7100 (Tokyo, Japón) a 100kV. Diez fotografías fueron seleccionadas para medir el diámetro y largo de 100 partículas virales, usando el programa informático ImageJ (Abramoff et al., 2004) para obtener el promedio de las mediciones.
El ARN total fue extraído de las ocho plantas sintomáticas evaluadas por MET, y dos plantas sanas, usando el RNeasy Plant Mini Kit (Qiagen, Alemania). La retro-transcripción fue realizada empleando el kit para síntesis de primera cadena de ADNc (Maxima H Minus, Thermo Scientific, Lituania) empleando los hexámeros aleatorios como iniciadores, siguiendo las recomendaciones del fabricante. Para determinar si las partículas virales con forma de bala, encontradas en hojas sintomáticas de papaya mediante TEM, corresponden tentativamente a nucleorhabdovirus o dichorhabdovirus, el ADNc fue evaluado para ambos géneros de virus de plantas mediante PCR punto final usando un par de iniciadores degenerados para rhabdovirus de plantas (Rhab-R / Rhab-F) desarrollados por Lamprecht et al. (2009) para amplificar el gen L que codifica la ARN polimerasa ARN dependiente (RdRp) viral, y para los dichorhavirus OFV y CiLV-N (LdF / LdR), diseñados por Ramos-González et al. (2016) para amplificar una región conservada del gen L (Cuadro 1). Posterior a esto, un segundo grupo de 24 muestras foliares de papaya fueron recolectadas (Junio, 2015) y también evaluadas usando el par de iniciadores Rhab-R / Rhab-F.
Doce amplicones obtenidos con el par de iniciadores Rhab-R / Rhab-F (Lamprecht et al., 2009) fueron purificados y secuenciados directamente en ambas direcciones (Macrogen, Korea) con el mismo par de iniciadores empleados en la PCR. La secuencia ensamblada correspondiente a cada amplicón fue obtenida usando BioEdit v7.7.1 (Hall 1999), y la búsqueda de similitud de secuencias fue realizada utilizando el algoritmo BLASTn en el GenBank (Centro Nacional para la Información de Biotecnología, EE. UU.). Cuatro de las secuencias obtenidas para un alphanucleorhabdovirus tentativo fueron depositadas en GenBank (PX637683, PX637684, PX637685 y PX637686).
Durante 2015-2016, adicionalmente 193 muestras foliares (126 de plantas sintomáticas) fueron recolectadas de plantaciones en Muelle, Guácimo y Guápiles, y analizadas siguiendo el protocolo RT-PCR para detectar rhabdovirus de plantas descrito previamente.
Para determinar la relación filogenética del virus con forma de bala observado, se realizó un análisis filogenético con cuatro de las secuencias obtenidas en este estudio (PX637683 a PX637686), incluyendo para su comparación a 31 diferentes rhabdovirus de plantas (un total de 34 secuencias debido a especies repetidas) y el virus Farmington (CT114) como grupo externo; todas obtenidas del GenBank (https://www.ncbi.nlm.nih.gov/genbank). El conjunto de datos final contó con un total de 1 099 posiciones. Las 39 secuencias se alinearon mediante el algoritmo ClustalW en BioEdit y luego se analizaron utilizando el método de Máxima Verosimilitud (modelo General Time Reversible y variación de tasas entre sitios mediante una distribución gamma con sitios invariables y 1 000 permutaciones) en MEGA 12 (Kumar et al., 2024).
Cuadro 1 Iniciadores y perfiles del termociclado empleados para detectar virus ARN en las plantas de Carica papaya analizadas en este estudio.
| Nombre del iniciador | Secuencia del oligonucleótido (referencia) | Par de iniciadores (tamaño del producto de la PCR), perfil térmico (referencia) |
|---|---|---|
| Rhabdovirus de plantas | ||
| Rhab-R | 5’GTCCABCCYTTTTGYC3´ | Rhab-R/ Rhab-F (900 pb, Lamprecht et al., 2009) |
| Rhab-F | 5’GGATMTGGGGBCATCC3’ | 94 ºC por 2 min; 36 ciclos de (94 ºC - 30 s; 37 ºC - 30 s; 72 ºC – 90 s); extensión final: 72 °C por 7 min (este estudio) |
| Rhab-R / Rhab-F (Lamprecht et al., 2009) | ||
| Dichorhavirus orchidaceae (Orchid fleck virus) y | ||
| Dichorhavirus leprosis (Citrus leprosis virus N) | ||
| LdF | 5’CCYGTGAGAGAATTCYTGGATG3’ | LdR/ LdF (390-400 pb) |
| LdR | 5’CCAGATTGGTGTARCCRAACAG3’ | 95 °C por 5 min, 35 ciclos de (95 °C por 30 s, 55 °C por 30 s), extensión final: 72 °C por 30 s. (Ramos - González et al., 2016) |
| LdF / LdR (Ramos - González et al., 2016) | ||
En plantaciones comerciales de papaya en las regiones norte y atlántica de Costa Rica, se observaron con frecuencia (50-70 % incidencia) plantas de papaya con cese evidente del cuajado de frutos (Figuras 1A y 1B), pecíolos curvados (Figuras 1B y 1E), estrías de color purpura en los pecíolos (Figura 1D); también se observó clorosis y arrugamiento de la lámina foliar, entrenudos y pedúnculos de frutos más cortos. Dependiendo de la etapa de crecimiento de la planta, se observaron claramente síntomas adicionales, como una menor cantidad de flores y frutos por planta; sin embargo, éstos no se asociaron con el aborto de frutos. Las pérdidas en la producción de frutos se estiman entre un 40 y 50 %, debido al cese irreversible del cuajado en las plantas enfermas y su eventual muerte regresiva (dieback). Al medir el ángulo de inserción de los pedúnculos de flores y frutos desde el ápice del tronco, las plantas infectadas presentaron un ángulo de inserción menor. Las plantas enfermas tenían pocas hojas en la copa (Figuras 1A y 1B), y algunas de éstas mostraban necrosis marginal en la lámina. Debido a la pérdida de follaje y la necrosis de las hojas apicales nuevas, las plantas parecen un lápiz con la punta hacia arriba en el campo, semejante al síntoma de la etapa final de plantas con la enfermedad del cogollo racimoso (PBT). Sin embargo, el látex estuvo presente en diferentes tejidos, incluyendo los frutos, contrario al síntoma típico del PBT (Davis et al., 1996).
Las diferentes pruebas de ELISA realizadas (PRSV, PapMV, CMV, TSWV, y el grupo de los potyvirus) en las plantas sintomáticas indicaron resultados negativos para las
70 muestras de papaya analizadas (Cuadro 2). Únicamente los controles positivos, utilizados para cada virus evaluado, mostraron un cambio de color en la solución del sustrato luego del tiempo de incubación, siguiendo los protocolos de Agdia Inc. (datos no mostrados).
Cuadro 2 Resumen de las pruebas realizadas y resultados obtenidos de las muestras foliares de plantas de Carica papaya, sintomáticas y asintomáticas, recolectadas en Costa Rica para determinar si algún virus está asociado a la enfermedad del “pecíolo rayado”.
| Región | ELISA | MET | RT-PCR | |||
|---|---|---|---|---|---|---|
| (positivo/ n evaluados) | (positivo/ n evaluados) | (positivo/ n evaluados) | ||||
| S | A | S | A | Sz | A | |
| Norte | 0/35 | 0/4 | 4/4 | -- | 60/61 | 0/28 |
| Atlántica, Guápiles | 0/18 | 0/3 | 2/2 | -- | 48/50 | 1/18 |
| Atlántica, Guácimo | 0/17 | 0/3 | 2/2 | -- | 42/47 | 0/21 |
| Valle Central | -- | -- | -- | 0/1 | -- | 0/2 |
| Total | 0/70 | 0/10 | 8/8 | 0/1 | 150/158 | 1/69 |
S o A= Muestras de plantas sintomáticas (S) o asintomáticas (A); ELISA: evaluaciones realizadas para: Potyvirus papayanuli, Potexvirus papayae, Orthotospovirus tomatomaculae, Cucumovirus CMV, y grupo de Potyvirus; MET: presencia de partículas en forma de bala en el núcleo celular; RT-PCR: amplicón de 900 pb obtenido; z: muestras evaluadas incluyó las 32 (8 y 24) de plantas sintomáticas recolectadas para las pruebas de validación usando los iniciadores Rhab-R /Rhab-F; --: No evaluado.

Figura 1 Alteraciones morfológicas observadas en plantas de Carica papaya con la enfermedad de “pecíolo rayado” en Costa Rica. A) Una planta sintomática (flecha) con los pecíolos curvos hacia abajo, reducción del número de hojas en el meristemo apical, y cese del cuajado de frutos. Adyacente (levemente atrás a la derecha) dos plantas sanas mostrando abundante desarrollo de frutos y la típica arquitectura foliar; B) Vista detallada de la sección apical de una planta sintomática que muestra reducción foliar en el meristemo, curvatura de pecíolos, y escaso cuajado de frutos, al fondo (izquierda) una planta sana con producción normal de frutos; C) Vista detallada de la sección superior de una planta sana; D) Pecíolos de una planta enferma con estrías de color púrpura; E) Comparación entre pecíolos de plantas sintomáticas (arriba) y sanas (abajo).
Las partículas virales en las plantas sintomáticas de papaya fueron observadas mediante el MET. Numerosas partículas con forma de bala fueron vistas dentro de los núcleos distendidos (Figuras 2A y 2C), “gemando” a través del espacio perinuclear (Figura 2B), y dentro de vesículas dispersas en el citoplasma celular, pero solamente en los tejidos de plantas enfermas. El promedio del diámetro y largo de las partículas virales (n=100) fue de 70 nm y 260 nm, respectivamente. Ninguna inclusión de tipo viral o partículas de otros virus fueron detectadas en las muestras.
En los análisis RT-PCR preliminares a ocho muestras sintomáticas para determinar la presencia de rhabdovirus de plantas o de dichorhavirus se obtuvo amplicones del tamaño esperado (900 pb) solo con el par de iniciadores para rhabdovirus de plantas (Rhab-R/Rhab-F); ninguna amplificación fue observada en plantas sanas. Subsecuentemente, un grupo adicional de muestras de 24 plantas sintomáticas consistentemente produjeron el amplicón de 900 pb correspondiente al rhabdovirus, mientras que ninguna amplificación fue detectada en muestras de papaya sana (control negativo). Por tanto, un rhabdovirus de plantas fue asociado con las plantas sintomáticas: inicialmente en un grupo de ocho muestras, luego en un grupo de 24 muestras, y finalmente en 95 % (150/158) de las muestras de papaya sintomáticas recolectadas en este estudio (2015-2016). En contraste, solamente el 1.4 % (1/69) de las muestras asintomáticas evaluadas fueron positivas (Cuadro 2).
Las secuencias de nucleótidos (Sanger) de 12 de los amplicones con mayor densidad obtenidos, confirmaron la asociación con un virus de plantas relacionado con los alphanucleorhabdovirus y disponible en GenBank. Los resultados de BLASTn para estas secuencias indicaron una similitud del 95.6 y 96.8 % con el papaya alphanucleorhabdovirus 1 (PNRV-1), GenBank Acc. Nos. MN203193 y MN203194, respectivamente. El análisis filogenético realizado a la raza costarricense relacionada al PNRV-1, mostró que ambos virus se agruparon en el clúster principal que contiene a todos los nucleorhabdovirus incluidos, y dentro del clado de los alphanucleorhabdovirus (Figura 3).
Síntomas similares a los reportados en Costa Rica, fueron descritos hace 45 años, en plantaciones de papaya en otros países en la cuenca del Caribe. Lastra y Quintero (1981) informaron de la necrosis apical de la papaya en Venezuela (estado de Zulia), inicialmente las plantas presentaban un arrugamiento foliar general, seguido por clorosis de hojas jóvenes, marchitez de las hojas ya desarrolladas, defoliación, y necrosis, llevando a la muerte de la planta. Wan y Conover (1981) también describieron la enfermedad del decaimiento necrosante de la papaya en Florida - EE. UU. (condados: Dade, Monroe y Sarasota), y detallaron la lista de síntomas asociados, como la re-curvatura o arqueo de los pecíolos hacia abajo, clorosis del follaje nuevo, pecíolos más cortos y rígidos (con algo de corrugación) con estrías grises, entrenudos cortos (generando una corona redondeada con apariencia racemosa), con inflorescencias estaminadas cortas y rígidas, aborto de flores pistiladas, cese del cuajado de frutos, sin aborto de frutos. Presencia de látex cuando se cortan los frutos o las plantas. En la fase final de la enfermedad se presenta abscisión de las hojas, necrosis marginal de las hojas nuevas y necrosis descendente del tallo. Además de estos reportes, síntomas de necrosis apical de la papaya se reportaron en Cuba, en las provincias de La Habana, Holguin y Villa Clara (Mejías et al., 1987; Hernández-Pérez et al., 1988), y México (estados de Oaxaca, Veracruz y Tabasco) (Becerra et al., 1999).
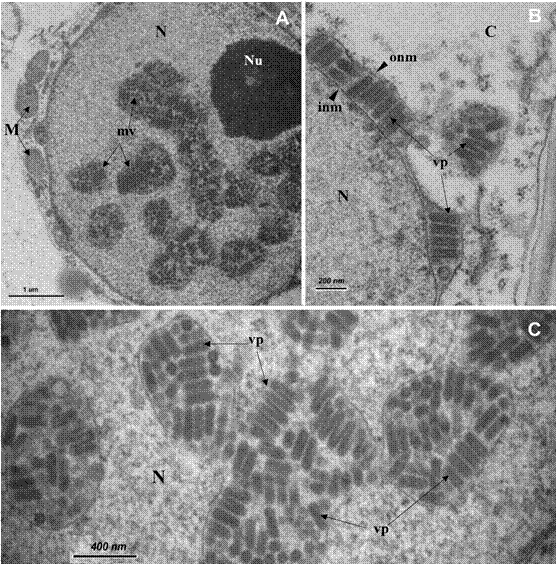
Figura 2 Núcleo de una célula de parénquima de una planta sintomática de Carica papaya con la enfermedad de “pecíolo rayado” observada con el microscopio electrónico de transmisión. A) Partículas con forma de bala dentro de vesículas membranosas en el núcleo celular; B) Detalle de las partículas vistas en el espacio perinuclear; C). Detalle de las partículas virales dentro de las vesículas membranosas dispersas en el núcleo. C= citoplasma, M= mitocondria, N= núcleo, Nu= nucleolo, inm= membrana nuclear interna, mv= vesículas membranosas llenas de partículas virales, onm= membrana nuclear externa, vp= partículas virales.
A pesar de que los virus PRSV y PapMV son dos fitopatógenos frecuentes que infectan al cultivo de la papaya en muchos países, ni ellos u otros virus evaluados por ELISA fueron detectados en las muestras recolectadas en las áreas de producción de las regiones en el norte y la zona atlántica de Costa Rica. En este país, la raza PRSV-W se ha informado infectando cucurbitáceas (Rivera et al., 1993) y otras plantas hospederas (Sánchez et al., 1998), pero no la raza PRSV-P. Asimismo, PapMV no se ha detectado en el país. Tanto el CMV, TSWV y algunos potyvirus se han encontrado en diferentes hospederos vegetales, pero no en papaya (Garita et al., datos sin publicar).
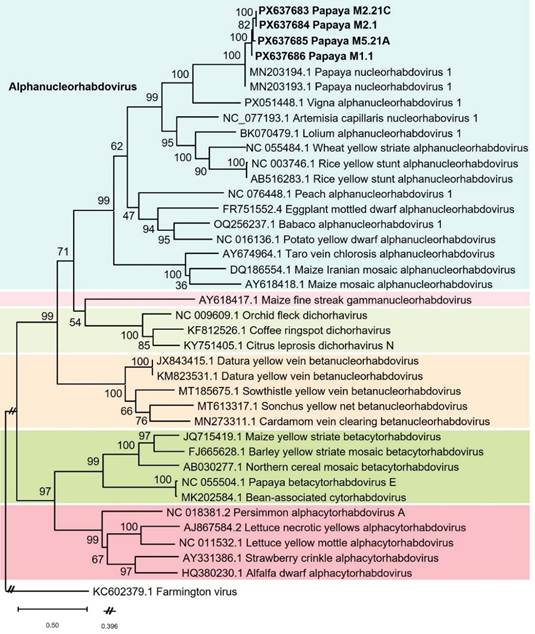
Figura 3 Árbol filogenético construido con las secuencias parciales (900 nt) de la ARN polimerasa dependiente del ARN viral (gen L) de un alphanucleorhabdovirus tentativo obtenido de Carica papaya en Costa Rica (GB Acc. No. PX637683 a PX637686) junto con 31 diferentes rhabdovirus de diferentes plantas (34 secuencias) y el virus Farmington (GB Acc. No. KC602379) como grupo externo, todas ellas recuperadas del GenBank. Éstas se alinearon con el algoritmo ClustalW utilizando BioEdit v.7.7.1, y la filogenia se infirió mediante el método de Máxima Verosimilitud en Mega 12 (modelo “General de Tiempo Reversible” (GTR) con una distribución gamma de tasas de sustitución con sitios invariantes) y un “Bootstrap” de 1000 réplicas.
Las partículas con forma de bala observadas en las muestras de papaya de Costa Rica, y su localización dentro del núcleo celular (Figura 2) semejan los rhabdovirus con forma de bala que infectan plantas (Jackson et al., 2005). Los núcleos hipertróficos conteniendo partículas en forma de balas fueron semejantes a los informados previamente, en muestras de papaya sintomática, de Venezuela (Lastra y Quintero, 1981), Florida - EE. UU. (Wan y Conover, 1981), Cuba (Mejías et al., 1987) y México (Becerra et al., 1999).
Los rhabdovirus de plantas (familia Rhabdoviridae) tienen genomas ARN simple banda de sentido negativo, infectan una amplia variedad de hospederos, tanto especies de mono- como de dicotiledóneas, y causan importantes enfermedades en los cultivos. Tradicionalmente, estos rhabdovirus de plantas se clasificaron como cytorhabdovirus y nucleorhabdovirus, basado en la organización del genoma, la filogenia, y el sitio de replicación intracelular. El género Nucleorhabdovirus recientemente fue dividido en los géneros: Alphanucleorhabdovirus, Betanucleorhabdovirus, Gammanucleorhabdovirus, y Deltanucleorhabdovirus (Simmonds et al., 2024; Walker et al., 2022; Dietzgen et al., 2020). La tecnología de secuenciación masiva de alto rendimiento (High-throughput sequencing, HTS), también conocida como secuenciación de nueva generación (NGS), ha permitido aumentar el número de virus de plantas de estos cuatro nuevos géneros, así como el ámbito de sus plantas hospedantes (Bejerman et al., 2021; 2025).
Las asociaciones entre enfermedades de papaya y partículas semejantes a las de rhabdovirus fueron informadas décadas antes de que las herramientas moleculares estuvieran disponibles. Recientemente, mediante la HTS se ha logrado la caracterización genómica de rhabdovirus infectando papaya, lo que permitirá una ubicación taxonómica más clara y la realización de estudios epidemiológicos. En plantaciones comerciales de papaya en Ecuador, un cytorhabdovirus prevalente fue caracterizado y nombrado papaya virus E (PpVE) por Medina-Salguero et al. (2019), y luego designado Cytorhabdovirus papayae (Bejerman and Dietzgen 2019).
Asimismo, el papaya nucleorhabdovirus 1 (PNRV-1) es un alphanucleorhabdovirus tentativo encontrado mediante análisis con metagenómica de muestras de papaya de Chiapas, México (GenBank Acc. No. MN203193- MN203195). Este rhabdovirus fue detectado en infecciones mixtas con otros virus, tanto en plantas con síntomas de PRSV y asintomáticas (Alcalá-Briseño et al., 2020). Otro presunto nucleorhabdovirus (babaco nucleorhabdovirus-1, BabRV-1, GenBank Acc. No. OQ256237) ha sido detectado recientemente infectando babaco (Vasconcellea × heilbornii) mediante metagenómica en Ecuador (Reyes-Proaño et al., 2023), pero éste no parece estar relacionado a los encontrados en México y Costa Rica, la comparación del PNRV-1 y el BabRV-1 no muestra una similitud significativa, de acuerdo al algoritmo BLAST. De acuerdo con los criterios de demarcación para Alphanucleorhabdovirus establecidos por el Comité International para la Taxonomía de Virus (ICTV), que considera identidad del genoma completo (umbral <75 %) y diferencias en el nicho ecológico, incluyendo hospederos y vectores, la alta similitud de nucleotídica (95.6-96.8 %) con el PNRV-1 del segmento parcial del gen L obtenido, además del hospedero que comparten (C. papaya), sugiere que éstos podrían corresponder al mismo virus. Sin embargo, la obtención de la secuencia genómica completa del segmento L o, idealmente, el genoma completo del aislamiento costarricense es necesario para confirmar definitivamente su identidad.
El análisis realizado con la raza de Costa Rica mostró que la raza mexicana (PNRV-1) y la costarricense están relacionadas y agrupan en el clúster principal conteniendo los alphanucleorhabdovirus (Figura 3). Desafortunadamente, las secuencias de los nucleorhabdovirus tentativos, previamente detectados por MET en papaya en Venezuela, Florida (EE. UU.), México o Cuba no están disponibles en el GenBank para poder compararlos. Sin embargo, los síntomas descritos en esos reportes y la asociación de partículas semejantes a rhabdovirus (forma de bala), sugiere que esos informes y el nucleorhabdovirus acá reportado para Costa Rica podrían corresponder a un antiguo fitopatógeno de papaya en esta región geográfica. La secuenciación de muestras de los diferentes países donde se reportaron estos síntomas contribuiría a realizar un análisis apropiado.
Este informe y los realizados previamente en la región, justifican la ejecución de estudios adicionales. Es imperativo completar los postulados de Koch para determinar la relación de los alphanucleorhabdovirus tentativos detectados en papaya con la sintomatología observada. Además, el estudio de asociaciones con otros virus que infectan este cultivo, la identificación de hospedantes alternas y la detección de posibles insectos vectores, ayudarán a entender los aspectos relacionados con la ecología y la epidemiología de esta enfermedad, permitiendo el diseño de estrategias de manejo adecuadas, en beneficio de los productores de papaya.
Conclusiones
Los resultados, obtenidos mediante el MET, mostraron que C. papaya afectada por la enfermedad del “pecíolo rayado” presenta partículas semejantes a las de los nucleorhabdovirus, y de acuerdo con las secuencias nucleotídicas obtenidas, el virus indicado está muy relacionado al papaya nucleorhabdovirus 1 (PNRV-1), un presunto alphanucleorhabdovirus encontrado empleando NGS por Alcalá-Briseño et al. (2020). Nuestra especulación, como mencionamos previamente, es que este virus puede estar relacionado a las detecciones mediante MET en papaya de Venezuela y Florida (EE. UU.) a principios de la década de 1980, y posteriormente en Cuba y México. Sin embargo, se necesita secuenciar plantas sintomáticas de papaya de los países de la cuenca del Caribe, para obtener la evidencia que soporte esta propuesta.
Hasta donde tenemos conocimiento, éste es el primer informe de un alphanucleorhabdovirus infectando papaya en Costa Rica. El virus se encontró asociado a una enfermedad viral emergente con efectos negativos para el cultivo; sin embargo, se requieren estudios adicionales para determinar el papel del virus en la etiología de la enfermedad y las especies de vectores asociadas.
Limitaciones. Las restricciones presupuestarias y el desconocimiento de la identidad del insecto vector (el cual se hipotetiza que es un cicadélido) representaron limitaciones para completar los postulados de Koch. Además, los rhabdovirus de plantas son difíciles de transmitir mecánicamente, requiriendo técnicas como la inoculación mediante punción vascular, la cual demanda pericia técnica y herramientas especializadas (Redinbaugh et al., 2001). Asimismo, es necesario: i) evaluar un número mayor de muestras para los diferentes virus recientemente asociados con enfermedades de la papaya en Latinoamérica para obtener un panorama completo del estatus en Costa Rica, ii) buscar plantas hospederas alternas, e iii) identificar el(los) vector(es). Esta información es necesaria para proponer las recomendaciones de manejo y estrategias para ayudar a los productores de papaya. Por tanto, aún se requiere de investigaciones adicionales para responder varias preguntas relacionadas al tema.
Por otro lado, es esencial obtener el genoma completo o secuencias más largas del alphanucleorhabdovirus consistentemente detectado en muestras de papaya sintomáticas de Costa Rica, ya sea mediante tecnología HTS u otro, para lograr una comparación apropiada con el papaya nucleorhabdovirus 1 reportado en México (Alcalá-Briseño et al., 2020).
Conflicto de interés. Los autores declaran que no existe ningún conflicto de interés con esta investigación.
Financiamiento. Este trabajo fue financiado por los subsidios para investigación brindados por FITTACORI (F24-14 y F17-16) y la Universidad de Costa Rica (B4-509 y A1-801).
Agradecimientos. Los autores aprecian enormemente: i. el apoyo brindado por los productores costarricenses, el PITTA PAPAYA, FITTACORI, y la Universidad de Costa Rica; ii. La gentileza de los investigadores: Ramón Lastra (Venezuela), Enrique Becerra (Mexico), y Ricardo Hernández-Pérez (Cuba), al compartir sus observaciones sobre esta enfermedad de papaya en sus países.
Contribución de los autores. Todos los autores contribuyeron al diseño del estudio, recolecta de muestra, procesamiento, y análisis de los datos. Todos revisaron y editaron el manuscrito, y aprobaron la versión final.

