











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink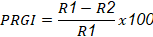
Introduction
The strawberry crop (Fragaria x ananasa) is of great importance to Mexico, which currently is third in terms of amount of strawberry produced in the world, with a production value of over 15,400 million pesos (SIAP, 2024). Nevertheless, there are pathogens that can reduce production and cause economic losses such as the wilting disease, caused by species of the genus Neopestalotiopsis, one of which was recently introduced and can result in production losses of over 70 %. Symptomatology includes rotting of the root, crown and foliar spots (Rebollar-Alviter et al., 2020), and fruits can present lesions of approximately 2-4 mm that extend to the point where they cause rotting and mummification (Goldenhar and Pate, 2021). Due to the shortness of information on the pathogen, control strategies on the field have been studied, such as cultural practices and chemical and biological control (Rebollar-Alviter et al., 2020). It has been pointed out that for the control of this pathogen, antagonistic agents can be used of the genera Trichoderma, Bacillus and Pseudomonas (Zhang et al., 2024;Amrutha and Vijayaraghavan, 2018). In particular, the antagonistic effect of Trichoderma is due to the action of enzymes, which can degrade the cell wall, chitin, proteins and lipids (Pandey et al. 2022). In addition, it produces secondary metabolites such as polyketides, non- ribosomal peptides, pyrones, antibiotic and others, which function as growth inhibitors, and can also act as a mycoparasite, and compete for space and nutrients (Mukherjee et al., 2012). In the case of control with agrochemicals, although there are products authorized for use on strawberry (COFEPRIS, 2024), whether as a synthetic molecule or plant extracts, the information on the control of Neopestalotiopsis is not precise. In this sense, the goal of this work was to evaluate the inhibiting effect of Trichoderma sp. strains on the growth of this Neopestalotiopsis sp. strain isolated from strawberry and the control with commercial fungicides used in some parts of the Guanajuato Bajio. The information generated displays the importance of the use of Trichoderma and commercial fungicides effective in the reduction of growth of this pathogen on the production of strawberry.
Materials and Methods
Reactivation of the Trichoderma sp. strains
The Trichoderma sp. strains were provided by the Agricultural Microbiology Laboratory of the Transformation Area of the Bajio Experimental Field (CEBAJ) of the National Forestry, Agricultural and Livestock Institute (INIFAP) and previously isolated from different substrates and places in Mexico (Table 1). The reactivation was performed in Potato Dextrose Agar (PDA) at 26±2 °C.
Table 1 Identification data, isolation substrate and origin of the Trichoderma sp. strains used in the biological control of
| Identification of strains | Substratum | Origin |
|---|---|---|
| 8L9 | Leachate | Talea de Castro, Oaxaca, México |
| 1752 | Agricultural soil | Santiago Coltzingo, Puebla, México |
| 1S1 | Compost | Palma Sola, Veracruz |
| THP | Agricultural soil | San José de Pantoja, Valle de Santiago, Guanajuato, México |
| 675, T1, T2, T3, T4, T5, T6, T7 | Agricultural soil, banana growing soils | Piedra Parada, Veracruz |
Isolation of Neopestalotiopsis sp. in strawberry and morphological characterization
In July, 2022, Neopestalotiopsis sp. was isolated from symptomatic strawberry plants, var. San Andres, from the CEBAJ orchard greenhouse. Leaves, fruits and stems were gathered and placed in plastic bags for transportation to the laboratory. Pieces of tissue measuring approximately 1 cm2 were cut and disinfected with NaClO 5 % for 60 seconds. They were washed three times with sterile water and the plant material was left to dry on a sterile paper towel. They were later planted in discardable Petri dishes containing PDA and amoxicillin (30 mg 1000 µL-1) and incubated at 26±2 °C. Once the pathogen sporulated, an acervulus was taken using a dissection needle and suspended in 1000 µL of sterile water. Later, the solution was homogenized in a vortex and 100 µL were taken for plating by spread technique. The incubation was carried out at 26±2 °C for 24 h. After this time and under a stereoscopic microscope, germinated conidia were placed in a PDA medium to obtain monosporic cultures, which were incubated at 26±2 °C. The morphological characterization of Neopestalotiopsis sp. was performed based on the description at a genus level with the characteristics described by Maharachchikumbura et al. (2012) and Maharachchikumbura et al. (2014).
Pathogenicity tests
To verify the pathogenicity of the isolation, the fungus was planted and left to grow for five days. Using a hole puncher, five discs, 5 mm in diameter, were taken from the invaded medium and placed on the crown of the strawberry. This procedure was carried out in 15 pots with healthy strawberry plants of the San Andres variety. Additionally, two discs were placed on leaves and stem to verify the pathogenicity of the fungus. The plants were watered every 48 h and covered with a polyethylene bag to reach a relative humidity
> 90 %. The symptomatology that arose after inoculation was observed (ai).
Antagonistic activity of Trichoderma on Neopestalotiopsis sp.
To determine the antagonistic activity, the dual culture method described by Correa et al. (2007) was used, which indicates a pathogen disc and another of the antagonistic fungus on opposite sides, 1 cm from the edge of the Petri dish (90 x 15 mm). The test was performed with eight repetitions in an antibiotic-free PDA medium. Discs measuring 5 mm in diameter were used, containing mycelium from the pathogen and strains of Trichoderma sp. previously grown on PDA for five days. The pathogen was given a growth advantage of three days. The incubation was carried out at 26±2 °C. The radial growth of the pathogen was measured every eight hours until 120 h. The percentage of radial growth inhibition (PRGI) was obtained from the growth of each pathogen in the dual culture, along with their respective controls, using the formula used by Suárez et al. (2008):
where PRGI=percentage of radial growth inhibition; R1= radial growth (mm) of the pathogen without Trichoderma, R2= radial growth (mm) of the pathogen with Trichoderma.
Additionally, the antagonistic ability of the chosen isolates selected was evaluated according to the scale by Bell et al. (1982), where Class 1: the antagonistic agent completely colonizes the phytopathogen and completely covers the surface of the medium; Class 2: the antagonistic agent colonizes two thirds of the culture medium, limiting the growth of the phytopathogen; Class 3: the antagonistic agent and the pathogen each colonize half of the surface, with the growth of each one being similar; Class 4: the phytopathogen colonizes two thirds of the surface of the culture medium and limits the growth of the antagonistic agent; Class 5: the phytopathogen completely colonizes the culture medium and grows on the antagonistic agent. The PRGI was analyzed with an analysis of variance (ANOVA) with a totally randomized design. The means comparison was performed using Tukey’s test (p< 0.05). The data were analyzed using the R Program, version 4.4.0.
Mycoparasitism
The presence of parasitism of the Trichoderma strains towards Neopestalotiopsis was observed to the dual microculture technique was used. In the trials, 15 Trichoderma strains were evaluated, in opposition to the pathogen Neopestalotiopsisis and three repetitions were carried out. The microcultures were prepared on slides on which a layer of medium (16 g bacteriological agar + 10 g PDA for 1 L), free of antibiotics, was applied. For the confrontations, discs 5 mm in diameter of the pathogen’s mycelium and Trichoderma previously grown on PDA for five days were used. On one end of the slide was a pathogen disc with a difference of 24 h, since the growth of the pathogen displayed a slower growth in comparison with Trichoderma. In the equidistant end, a disc of Trichoderma was placed. The preparations were incubated at 26±2 °C. When both fungi came into contact (approximately 24 h), the slide was removed from the Petri dish and using transparent scotch tape, a sample of mycelium was taken and a drop of cotton blue was added to be able to observe under the microscope and determine the type of parasitism of the Trichoderma strains.
Growth inhibition test Neopestalotiopsis with commercial fungicides
The effect of inhibition of the growth of pathogens was determined in five commercial products: the contact fungicide Captan (Captan 50®), the systemic fungicide Tebuconazole (Folicur®), the systemic fungicide Carbendazim (Agrosys®), the Cinnamon and Neem Extract (Nymeria®), peracetic acid (Microclean®) and a citrus- based organic fungicide for sale locally. Each fungicide was evaluated in three concentration (Table 2), which were chosen considering the recommendations by the manufacturer in the technical details; a minimal concentration was used, along with an intermediate one and the highest one recommended. Five repetitions were carried out for every concentration plus one control. The tests were carried out in Petri dishes with PDA free of antibiotics. The fungicides were incorporated into the medium when it reached a temperature of 45-50 °C. At the center of each dish, a 5 mm disc of the Neopestalotiopsis growth, previously grown for 5 days in PDA medium was placed. Every 8 h, the diameter of growth was determined for 120 h. The control was established in PDA without fungicide. The tests were carried out in five repetitions. For the statistical analysis, the data were arranged in a totally randomized design. These data were analyzed in an ANOVA using the R program, version 4.4.0. The multiple means comparison was carried out using Tukey’s method (p≤0.05).
Table 2 Fungicides evaluated in Neopestalotiopsis sp. growth inhibition.
| Fungicide | Dosis (L-1) |
|---|---|
| Captan | 2 g (A), 3 g (B), 4 g (C) |
| Tebuconazol | 1 mL (D), 2.5 mL (E), 3.75 mL (F) |
| Carbendazim | 4 mL (G), 5 mL (H), 6 mL (I) |
| Cinnamon and Neem Extract | 5 mL (J), 10 mL (K), 15 mL (L) |
| Peracetic acid | 0.25 mL (M), 0.50 mL (N), 0.75 mL (O) |
| Citrus-based organic fungicide | 5 mL (P), 7.5 mL (Q), 10 mL (R) |
Note: Letters in parentheses will be used to refer to the treatment applied.
Results and Discussion
Identification of symptoms in strawberry plants
Symptoms were found in plants according to reports by Rebollar-Alviter et al. (2020): blight in leaves, regressive death of sprouts, rotting and deformation of fruits, stunting of plants, as well as rotting of the root and crown (Figure 1). These symptoms are similar among Neopestalotiopsis species. In this regard, Baggio et al. (2021) mention that the tissues most affected by N. rosae are those of the root and neck of the root, although there is a low incidence on fruits and leaves. Likewise, authors such as Obregón et al. (2018) indicated that N. clavispora shows symptoms on leaves, which consist of reddish necrotic interveinal areas that begin at the edges and progress as the tissue becomes senescent. According to the authors, the root and crown are also affected, ultimately leading to the death of the plant. The species N. clavispora causes blight on the calyx and the receptacle of strawberries (Shi et al., 2022).

Identifying the causal agent
After growing for eight days, the colonies in PDA medium presented a cottonlike structure, a white mycelium which slowly turned to a salmon color and concentric edges. When the culture was six days old, black spore masses formed (Figure 2). After 12 days, black globose acervuli were produced on the growth medium and on the tissues the stromata presented an irregular shape (Figure 2). The conidia presented three dark central cells and two basal hyalines with three apical appendices (Figure 2). The morphology of the pathogen coincided with the characteristics of the genus described by Maharachchikumbura et al. (2014). However, according to some characteristics pointed out by Maharachchikumbura et al. (2012), the Neopestaloptiopsis isolate in this study could be from N. rosae, a species that was confirmed to be present in strawberry crops in Mexico (Rebollar-Alviter et al., 2020).

Figure 2 Growth of Neopestalotiopsis sp. in a PDA medium after two weeks of inoculation. A) Development of colonies in a culture medium with black acervuli, B and C) Fungal conidia.
Patogenicity of Neopestaloptiosis sp.
The plants inoculated with the isolate presented symptoms 24 h ai (Figure 3) in which small, maroon spots sized 2 mm were initially observed. After 56 h ai, leaf blight was observed, along with stem necrosis and weakening of the plant (Figure 3). On the 168th hour after inoculation, the damage was highly advanced, approximately 70 % of the stem presented necrosis and total softening. The spots present in the leaves covered over 80 % of the foliar surface (Figure 3). The symptomatology coincided with the one documented by Rebollar Alviter et al. (2020) and Gilardi et al. (2019) in strawberry crops, both in leaves and fruits.
Antagonistic activity of Trichoderma on Neopestalotiopsis
The statistical analysis indicated significant differences (p < 0.05) in the growth inhibition between the evaluated strains. According to the means comparison, three inhibition groups were found, strain T1 displayed the greatest PRGI (70 %), while strains T675, T2 and T10 displayed the least inhibition (Figure 4). In general, the PRGI remained between 60 and 70 %, so the strains can be considered to have the potential to control the pathogen. Regarding this, Yao et al. (2023), after an analysis of different studies, indicate that a strain of Trichoderma can be considered to have the potential in the effective control of the disease when growth inhibition surpasses 50 %.

Figure 3 Symptoms on strawberry plants caused by Neopestalotiopsis sp. A) Blight on leaves, B) Necrosis on the stem and C) Necrosis in neck and leaves.
Some authors have evaluated the effect of Trichoderma for the control of Neopestalotiopsis. One of the first studies was the one by Mora-Morales et al. (2020), who evaluated the antagonistic in vitro effect of T. harzianum on Pestalotiopsis (Neopestalotiopsis sp.), and their results showed a PRGI of 71.11 %. Other authors such as Amrutha and Vijayaraghavan (2018) evaluated the antagonistic effect of T. asperellum against Neopestalotiopsis clavispora isolated from strawberry, and determined an average inhibition of 66 % in the growth of the pathogen in in vitro tests, as well as a reduction of 74 % of the disease. The authors indicated that, in order to manage the disease with biological and chemical products, it is necessary to consider the agroclimatic conditions for a greater efficiency in the control. In another study carried out by Liu et al. (2024), in which they evaluated the antagonistic effect of volatile metabolites and fermentation metabolites produced by T. asperellum on the growth of N. clavispora, they determined inhibitions of 79.67 and 69.84 %, respectively. They also identified that the antagonistic effect was due to the competition for the substrate, the production of antimycotics and parasitism. Additionally, they indicated that the use of T. asperellum promoted the growth in the strawberry plant, its height, the root length, the total fresh weight, the root weight, the fresh weight of the stem and the dry weight of the root. On the other hand, Pandey et al. (2022) determined that T. reesei influenced the in vitro growth of Pseudopestalotiopsis theae. Their results displayed an inhibition of 81.2 %. When they took the experiment to plants in greenhouse conditions, the severity of the disease was between 67.5 and 75 %. In this sense, the authors indicated that T. reesei was able to produce volatile and non- volatile metabolites with an inhibiting effect. In addition, in plants inoculated with the antagonist, they found an accumulation of defense enzymes such as polyphenol oxidase, peroxidase, phenylalanine ammonia-lyase, β-1, 3-glucanase, and chitinase. Likewise, at 45 days, these plants displayed an increase in height, stem diameter, and the weight of shoots and roots.






Figure 4 Percentage of inhibition of root growth for Neopestalotiopsis sp. obtained from the confrontation with Trichoderma strains (p≤0.05).
Antagonism of the Trichoderma strains
According to the scale by Bell et al. (1982), antagonism levels II and III were found (Table 3, Figure 5), i.e., in type II, the Trichoderma strains colonized two thirds of the culture medium, limiting the growth of the phytopathogen; in type III, Trichoderma and the pathogen each colonized half of the surface; growth was similar among strains (Table 3). Diverse authors have indicated that Trichoderma has different antagonism mechanisms, highlighting mycoparasitism, competition over space and food and antibiosis (Álvarez et al., 2022). One of the main mechanisms is mycoparasitism, in which structures may take the shapes of hooks, curl or an appressorium around the host, allowing it to penetrate into the pathogen and degrade it (Mukherjee et al., 2022; Álvarez et al., 2022). This mechanism comes with the secretion of enzymes such as chitin to degrade the cell wall of the pathogen (Yao et al., 2023; Companioni-González et al., 2019).
This study found the mycoparasitism mechanisms: adhesion and curling of the Trichoderma strains to the hypha of the pathogen, as well as the hyphal lysis, the formation of hooks and penetration of the antagonist into the pathogen (Figure 6).

Figure 5 Confrontations of Trichoderma strains against Neopestalotiopsis sp. where T = Trichoderma and P = pathogen. The number found on the top right corner corresponds to the identification of the Trichoderma strain. T1, T5 and T7 are observed to maintain a level II antagonism, while T4, T8 and T10 maintain a level III.

Figure 6 Mechanisms of parasitism in Trichoderma sp against Neopestaloptiopsis sp. A) Curling, B) Formation of hooks and C) Adhesion and lysis.
Table 3 Level of antagonism according to Bell et al. (1982) and mycoparasitim mechanisms.
| Nivel de antagonismo | Cepa | Mecanismo del micoparasitismo |
|---|---|---|
| II | 1S1, T175, T675, T5 | Adhesion, curling |
| II | 8L9, T3 | Curling, formation of hooks |
| II | THP | Lysis, curling |
| II | T1, T2 | Adhesion |
| II | T3 | Curling, formation of hooks |
| II | T6 | Adhesion, curling, formation of hooks |
| II | T7 | Adhesion, penetration hyphal |
| III | T4, T8 | Curling |
| III | T9, T10 | Curling, formation of hooks |
Evaluation of control of Neopestalotiopsis sp. using commercial fungicides
According to the statistical analysis, significant differences were found between treatments. The means comparison indicated that the systemic fungicide Tebuconazole, the cinnamon and neem extract and peracetic acid, in all concentrations used, inhibited the growth of Neopestalotiopsis sp. by 100 % (Figure 7). Captan, the contact fungicide, inhibited the growth of the pathogen by 81, 83 and 86 %, followed by the citrus-based organic fungicide with 52, 60 and 63 %; and the systemic fungicide Carbendazim inhibited it by 28, 42 and 52 % in regard to the control.

Figure 7 Growth of Neopestalotiopsis sp in a PDA medium with commercial fungicides. Test: control with no product; A-C: Captan 200, 300 and 400 g 100 L-1; D-F: Tebuconazole 100, 250 and 375 mL 100 L-1; G-I: Carbendazim 400, 500 and 600 mL 100 L-1; J-L: Extract of Cinnamon and Neem 500, 1000 and 1500 mL 100 L-1; M-O: Peracetic Acid 25, 50 and 75 mL 100 L-1; P-R: Citrus-based organic fungicide 500, 750 and 1000 mL 100 L- 1. 400 g and 600 mL 100 L-1 (p≤0.05).
Due to the large economic losses caused by Neopestaloptiopsis, studies have been performed to determine the management of the disease. In this regard, Rebollar-Alviter et al. (2020) evaluated the in vitro control of Neopestaloptiosis rosae using Captan and other products such as Azoxystrobin, Fluxapyroxad + Pyraclostrobin, Pyraclostrobin + Boscalid, Iprodion, Difenoconazole, Prochloraz, Cyprodinil + Fludioxonil. The results in terms of inhibition in the growth of the pathogens suggested that Captan, Fludioxonil + Cyprodinil, Difenoconazole and Iprodione had the greatest effect for the control of Neopestaloptiopsis. Likewise, Baggio et al. (2023) held studies to determine the sensitivity of the pathogen to commercial fungicides, and their results indicated that Captan, Azoxystrobin, Fluxapyroxad + Pyraclostrobin, Pyraclostrobin + Boscalid, Iprodiona, Difenoconazole, Prochloraz, Cyprodinil + Fludioxonil.were more effective. Authors such as Acosta-González et al. (2024) pointed out that, when applying fungicide, particularly Pidiflumetofen + Fludioxonil, Ciprodinil + Fludioxonil and Procloraz preventively, they were able to reduce the incidence and severity of the disease, as well as the mortality of plants. The authors indicated that when the fungicides were applied preventively, they obtained an efficiency of 99 to 100 %, although when using them curatively, that is, after the symptoms were expressed, the efficiency was between 0 and 37 %.
On the other hand, products based on the extracts of plants such as garlic, chamomile, rue, cinnamon, marigold, acacia, chili pepper, pepper, agave, rue, castor, prickly poppy, pine, citrus, creosote and others are authorized in Mexico as pesticides for strawberry crops (COFEPRIS, 2024). Therefore, they are an important alternative for the organic management of this crop. The results of this study with the extract of cinnamon and neem showed that they can be applied to inhibit the growth of the pathogen without causing long-term environmental harm. Other authors such as Darapanit et al. (2021) showed that ethanolic extracts based on turmeric (Curcuma longa), ginger (Zingiber officinale), lemongrass (Cymbopogon citratus), hibiscus (Hibiscus sabdariffa) and clove (Syzygium aromaticum) inhibited the growth of Neopestalotiopsis and Pseudopestalotiopsis strains, particularly the turmeric and clove extracts at concentrations of 10,000 mg L-1. In general, plant-based extracts produce secondary metabolites such as steroids, alkaloids, tannins, terpenes, phenols, flavonoids and resins, which exhibit antimicrobial properties (Lengai et al., 2020).
Conclusions
The in vitro control obtained with the 15 Trichoderma strains evaluated was determined by the PRGI in an interval of 63 to 70 %. Strain T1 displayed the greatest percentage of inhibition in the radial growth of the pathogen. The level of antagonism found was types II and III, with parasitism mechanisms of curling, adhesion to the hypha of the pathogen and lysis. In the case of the pathogen growth control with the fungicides, an inhibition of 100 % was determined with Tebuconazole, Cinnamon Extract and Neem and Peracetic Acid. The active ingredients Captan and Carbendazim displayed a growth inhibition of 86 and 52 % with the doses 400 g and 600 mL 100 L-1, respectively. The results of this study serve as a reference for the development of an integrated management approach with preventive and control applications using Trichoderma sp. and organic products as an alternative to chemical molecules, which contributes towards the reduction of primary inoculum and provides protection to plants once established in the field.

