










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias forestales
versão impressa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.6 no.32 México Nov./Dez. 2015
Articles
Molecular identification of the Botryosphaeria sp. complex related to cankers and drying of buds in Eucalyptus sp.
1Maestría en Ciencias Forestales. Universidad Autónoma Chapingo.
2Laboratorio de Biología Molecular Departamento de Fitotecnia. Universidad Autónoma Chapingo.
Phytosanitary problems drastically affect the production or the plants of Eucalyptus sp. from the damages that they bring. Botryosphaeria sp. is associated with cankers and drying buds, dieback, fruit rot, collar rot and damping off in seedlings, which is especially important in commercial plantations that use these species for lumber in short cycles. In order to characterize and identify at the molecular level isolates of this fungus, in the work described below is estimated, first, the genetic variability in diseased samples with genomic markers of the ISSR type, and secondly, the ITS region was sequenced and compared. The ISSR markers were analyzed with multivariate techniques to group the isolates based on their genetic similarity, of what turned out that most of the samples were very similar to each other at the genome. The results of the sequencing technique IST regions indicated the presence of Neofusicoccum sp., Neofusicoccum parvum , Diplodia corticola, Lasiodiplodia pseudotheobromae, Neofusicoccum eucalyptorum and Neofusicoccum sp. It is noteworthy that at the nucleotide level of the sequenced region, the quantity of polymorphisms among the isolates studied and those that were used as outgroups obtained from GenBank, was minimal.
Key words: DNA; cankers; ISSR; MEGA; ITS sequence, UPGMA
Los problemas fitosanitarios afectan de manera drástica al cultivo o a la planta de eucalipto, por los daños que le producen. Botryosphaeria sp. está asociado a cancros y secamiento de yemas, muerte descendente, pudrición de frutos, pudrición del cuello y damping-off en plántulas, lo que es particularmente importante en las plantaciones comerciales de Eucalyptus para obtener madera en ciclos cortos. Con el objetivo de caracterizar e identificar a nivel molecular aislamientos de este hongo, en el trabajo que se describe a continuación se estimó, por un lado, la variabilidad genética en muestras enfermas con marcadores genómicos tipo ISSR, y por otro, se secuenció y comparó la región ITS. Los ISSR fueron analizados con técnicas multivariadas para agrupar a los aislamientos con base en su similitud genética, de lo que resultó que la mayoría de las muestras fueron muy similares entre sí, a nivel de genoma. Los resultados de la técnica de secuenciación de las regiones ITS indicaron la presencia de Neofusicoccum sp., Neofusicoccum parvum, Diplodia corticola, Lasiodiplodia pseudotheobromae, Neofusicoccum eucalyptorum y Neofusicoccum sp. En relación con los nucleótidos de la región secuenciada, la cantidad de polimorfismos entre los aislamientos estudiados y aquellos que se usaron como outgroups, obtenidos del , fue mínimo.
Palabras clave: ADN; cancros; ISSR; MEGA; secuencias ITS; UPGMA
Introduction
The use of forest exotic species offers, in most cases, the advantages over native species, in regard to the growth rate and cutting shifts, the latter which tend to be shorter; both criteria are essential to develop viable forest projects from the financial point of view. Eucalyptus species are used to get timber in short cycles but plantations are affected by pathogens and insects; Botyosphaeria fungus is causing severe diseases in such trees.
In Mexico there are commercial plantations of fast-growing eucalyptus (Eucalyptus camaldulensis Dehnh., E. globulus Labill., E. grandis W. Hill, E. nitens Deane & Maiden, E. pellita F. Muell., E. urophylla S. T. Blake and the E. urophylla x grandis (E. urograndis) hybrid (Velázquez et al., 2013) established in the Estado de México, Guerrero, Michoacán, Oaxaca, Tabasco and Veracruz.
From this group of species, E. nitens and E. globulus are outstanding in environments at higher altitude prone to frost and E. urophylla in the humid tropics (Conafor, 2012). In the years 2010-2012, the first two taxa were used in plantations in the state of Michoacán, where mortality was recorded in trees aged three to four years old; the organisms involved in the dead or diseased tissues were fungi belonging to the Botryosphaeriacea (Botryosphaeriales, Ascomycetes) family (De la Mora et al., 2014). In southern Veracruz, in plantations of E. grandis and E. urophylla the existence of B. rhodina (Berk & M. A. Curtis, 1970) was noted as the causing agent of dieback; the damaged trees showed root entanglement, so it is presumed that the fungi played a role of opportunistic pathogen (Cibrián, 2013).
Fungi of this family are widespread in the world and affect a diverse group of Dicotyledoneae, Monocotyledoneae and gymnosperms (Smith et al., 2001; Mohali et al., 2006; Pérez et al., 2009, Pérez et al., 2010). Botryosphaeria and Neofusiccocum genera are endophyte pathogens, opportunists, inducing cankers on stems and branches, dieback, fruit rot, collar rot and damping off in seedlings (Mohali et al, 2006; Slippers and Wingfield, 2007).
Originally 13 species were determined for the Neofusicoccum genus number grows as additional studies are made (Sakalidis et al., 2011). The molecular techniques based on DNA markers and sequencing are very supportive of the taxonomy of organisms. ISSR markers in particular have been used with great success by the results they offer and the relative ease with which they are obtained. For purposes of molecular identification, sequence comparisons or regions of genes associated with phylogeny (for example, ITS zones) it has revolutionized the current taxonomy of many eukaryotic organisms and are widely used (Jin et al, 2003; Awasthi et al, 2004).
The aim of this study was to characterize, compare and identify molecularly isolates of Botryosphaeria sp., associated with cankers and drying of isolated buds of Eucalyptus sp., due to the importance of these fungi in the forest, urban, restoration, agroforestry and commercial plantations.
Materials y Methods
Sample collection
The assays were made with 14 Eucalyptus samples with canker symptoms, from which only those with Botryosphaeria sp. were taken into account. Nine came from plots of Forestaciones Operativas de México S. A. de C V. (REXCEL) (EFOMSA) company and five from Forestales PROTEAK company during 2013 and 2014 (Table 1).
Isolation and purification of the fungal isolates
Samples were disinfected with commercial 3 % sodium hypochlorite for 5 minutes; they were rinsed three times in sterile distilled water for 5 min each; they were dried with sterile absorbent paper and three sections of each sample were sown in potato-dextrose agar 2 % (PDA, Bioxon) culture medium at 25 ± 2 °C (considered as replications, which made a total of 42 samples) which were maintained for 10 days. Purification of each isolate characteristics of Botryosphaeria sp. was performed by hyphae tip, for which, once the mycelium developed in PDA, a section of about 1 mm was sown in Petri dishes with water agar (AA, for its acronym in Spanish) medium and kept for 24 h at the same temperature. AA is a poor culture medium in which the mycelium grows in a very thin form, and in order to grow the mycelia in it was to achieve greater accuracy when taking the fraction of the hyphae. At the same time, another section of it about 1 mm was transferred to Malt Extract-Agar 2 % (MEA) culture medium and kept so for 20-30 days for its development in temperature conditions similar to those mentioned before.
Morphological characterization
From the isolates developed in MEA culture medium, the mycelium color and the type of fungus growth were evaluated. In the AG-C I 114, ED-I PM 43 and FR-M isolates at day 25 the presence of black globular pycnidia covered by gray mycelium were observed but without the formation of conidia. These samples were processed according to Slippers and Wingfield (2007), who stated that in order to induce the formation of conidia in pycnidia the fungus should be grown in the AA or MAE medium supplemented with pine needles or small pieces of tissue from the host at 25 °C. The FR-M sample (plot 4) was used for the identification of such structures through slides from a Carl Zeiss compound microscope with the HardWare
Setup, Microscope: Leica DM2500, Test None. Image-Source: Leica DCF/DVM/ICD/IC3D Camera (Firewire A/B) program. The genus identification was performed with Barnett and Hunter (1998) and Sivanesan (1984) keys and the species was identified based on its morphology according to Smith et al. (2001).
Molecular characterization and identification
In order to compare the genetic variability of the 42 studied fungi samples ISSR type DNA fingerprints were obtained. DNA was extracted from the monoconidial cultures purified in MEA with the CTAB I protocol from the recommendations of Weising et al. (2005). The DNA quality was confirmed in 1.2 % agarose gel in TAE 1X buffer (Tris-basis, glaciar acetic acid, EDTA 0.5 M pH 8.0). The high molecular weight DNA was visualized through a Universal Hood II (Bio-Rad) model transiluminator and its concentration was quantified in a NanoDrop Lambda Bio 12 model (Perkin Elmer) spectrophotometer.
Development of ISSR Markers
The (AC)8YG, (GA)8YC, (GACA)4 (GGAT)4 and (GATA)4 initiators were used to describe the isolations. The PCR reactions were carried out in a 25 μL final volume. The reaction mix included 1X Buffer, MgCl2 2.5 mM, DNA Taq polymerase 1.5 U (Promega), dNTP 200 μM, Initiator 20 pM, DNA 100 ng and water.
Thermocycling conditions
The samples were amplified in the Gene Amp PCR System 9700 (Applied Biosystems) thermal cycler under the following program: one cycle at 95 °C, 3 min; 35 cycles [95 °C, 30 sec; 56 °C, 45 sec; 72 °C, 2 min] and final extension cycle at 72 °C for 10 min. The alignment temperature for the primers (GACA)4, (GGAT)4 and (GATA)4 was 48 °C. For separating the amplified fragments, a vertical chamber C.B.S. Scientific CO® MVG-216-33 model with a power supply [Corporations® E-C Apparatus, Model EC-105] was used. The acrylamide gel was 6 %, and was run at 245 volts for 2.5 hours, approximately. 2 μL of the molecular 100 bp DNA (Promega) weight marker and 3 μL of 1 kb O'GeneRuler™ marker (Fermentas) were used for bands location purposes.
From the DNA profiles observed in the acrylamide gels, the percentage of mono and polymorphic bands was determined and with the data, a binary matrix was made. A dendrogram was built with the Free-Tree program 0.9.1.50 version and the Nei and Li/Dice coefficient by the UPGMA clustering method; the bootstrapping resampling was with 100 replicates. To display the dendrogram the Tree View 1.6.6 program was used. Wherein the genetic distance of the samples tested was observed.
Sequencing of the ITS region
For molecular identification, a representative of the 14 initial isolates of the seek Eucalyptus isolates was used because the profiles of the 3 samples considered as replications were the same. The ITS region (internal transcribed spacers) was amplified with the 5HP universal primers (5'-GGA AGG AGA AGT CGT AAC AAG G-3') and NL 4 (5'-GGT CCG TGT TTC AAG ACG G-3'). The PCR reaction conditions were: Buffer 1X, MgCl2 2.5 mM, Taq DNA polymerase 1.5 U (Promega), dNTP 200 μM, 10 pM primer each, DNA 100 ng and water. The thermocycling program consisted of one cycle at 95 °C for 4 min; 35 cycles [94 °C for 1 min; 58 °C, for 1 min; 72 °C for 2 min] and a cycle of final extension at 72 °C for 10 min. Amplifications were visualized on 1.2 % agarose gels. Images were documented with the support of the Quantity One (Bio-Rad) software.
ITS products were cleaned with EXO-SAP prior to sequencing. After obtaining the sequences they were edited with BioEdit and FinchTV programs and aligned with ClustalW. They were then compared to some reported in the GenBank (outgroup) using the BLAST program (http://www.ncbi.nlm.nih.gov/). Alignment of all sequences was performed in 6.05 MEGA program and the phylogenetic tree constructed using the statistical method of Maximum Parsimony (MP) using 1 000 bootstrap replicates. The sequences obtained were uploaded to the NCBI database.
Results and Discussion
Phenotypic characterization
The mycelium of some of the isolates (AG-C I 55, I 143 ED-PM, MAR, MI-T I 2, VC1, FR-M and AG-C) showed white fluffy appearance at day 5 and at the 10th, it turned from dark grey to black, as well as airy and abundant; these samples recorded greater growth speed. The AG-C I 114, JA-S, RA-A, VC2, VC3, VC4 and VC5 samples were slower when colonizing the Petri boxes; The color of the mycelium was black and it was stuck to the culture medium.
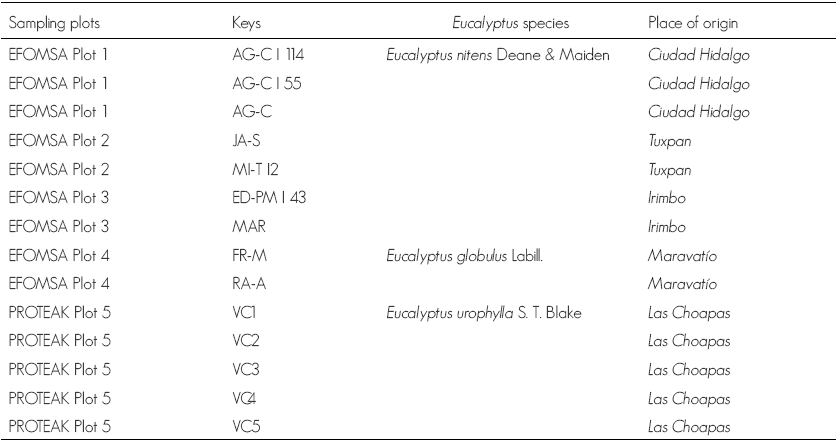
At 25 days, in the FR-M sample dark brown to black pseudothecia were observed immersed in the EMA culture medium; and the presence of pycnidia, conidia and mature asci with ascospores (Figure 1). These morphological characteristics are consistent with those reported by Smith et al. (2001) to Botryosphaeria eucalyptorum Crous, H. J. M. Smith et Wingf. sp. in nov.
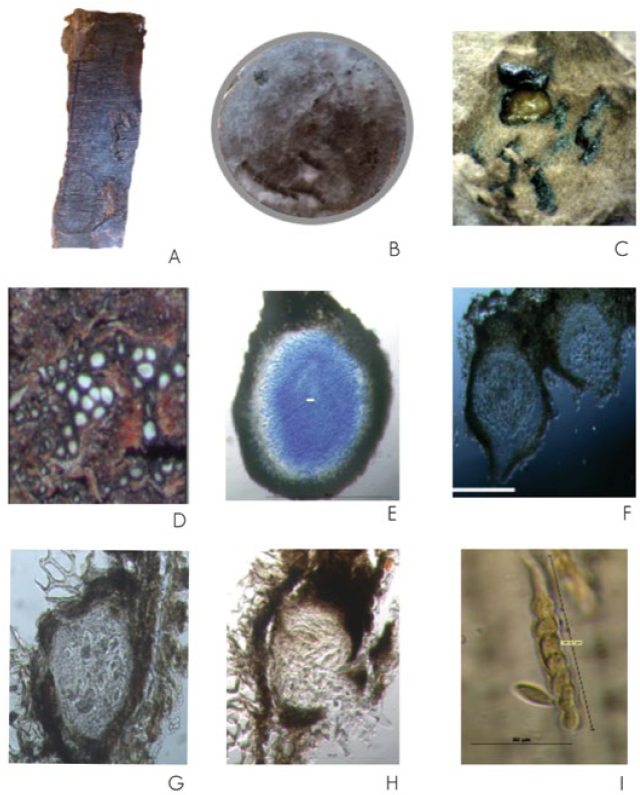
Figure 1 A) Part of the seek tree that was used; B) Neofusicoccum sp. in MEA at 25 days; C) Look of the ascomes; D) Longitudinal cut of the pycnidia; E) Pycnidium and conidium; F-H) Pseudothecia; I) Asca with eight ascospores observed at 60X with a compound microscope.
Another feature that has been considered for the classification of the Botryosphaeria genus were the teleomorphic (sexual) and anamorphic (asexual) forms. But up to date, the taxonomy of the species of this genus is mainly dependent on the anamorphic, because some of the traits used for identification as the color of the conidia, the presence of septa, or stromal morphology as they have great plasticity within the genus (Denman et al., 2000).
ISSR
All the primers that were used were able to generate DNA fingerprint profiles; (AC)8YG and (GACA)4 in particular were the most informative as they amplified a greater number of bands and showed higher polymorphism compared to the rest of the others; therefore, they were selected to make the corresponding clusters (Figure 2). The opposite condition occurred for (GA)8YC,(GGAT)4 and (GATA)4 which amplified a few bands, and thus, it was decided not to include them in the data analysis. With the two selected primers fragments of different molecular weight were viewed and for counting the DNA bands only those of 300 bp up to 1 500 bp were chosen, as they had the best consistency and acrylamide gel reproducibility. It is worth noting that the PCR reactions and the acrylamide gels were run at least thrice in an independent way, without observing variations in the results.
Figure 2 DNA fingerprints from Botryospaheria isolates obtained with the (GACA)4 primer separated in 6 % acrylamide gel. Line 1 and 44 molecular 100 bp weight marker and 1 ksb Fermentas™.
With data derived from the gels and based on the molecular weight of the fragments, a binary matrix was made, which was coded with digit 1 for present bands and 0 for absent bands. In order to establish genomic groups with ISSR electrophoretic patterns and correlate them visually with the different observed mycelia in the isolates, a radial shape dendrogram was built (Figure 3), in which, mainly, four groups are recognized: group 1 is represented by MAR- Neofusicoccum sp., VC2-Neofusicoccum sp. and VC1-Diplodia corticola A. J. L. Phillips, A. Alves & J. Luque 2004; group II, which contains VC3-N. parvum (Pennycook & Samuels) Crous, Slippers & A. J. Phillips, 2006, FR-M Neofusicoccum sp. and VC5 Neofusicoccum sp.; group III consisting of JA-S Neofusicoccum sp., AG-C I 55 N. parvum, AG-C I 114 Neofusicoccum sp., VC4 -Neofusicoccum sp., ED-PM I 43-N. parvum., MI-T I 2 N eucalyptorum (Crous, H. Sm. ter & M. J. Wingf.) Crous, Slippers & A. J. Phillips, 2006, and AG-C N. parvum; and group IV, which includes only RA-A-Lasiodiplodia pseudotheobromaeA. J. L. Phillips. A. Alves & Crous 2008. As shown, the color of the mycelia isolates within each group varies, even within the same species it may be different as in the case of Neofusicoccum sp.

Figure 3 Dendrogram constructed from the ISSR profiles obtained with the (GACA)4 and (AC)8YG primers, which exhibits the genomic relations of the Botryospheria genus. The variability in color and growth shape of the mycelia of the different isolates are shown.
The profiles of the DNA fingerprints of the different isolates revealed a high genomic relationship between them. In the corresponding dendrogram species are not clearly divided, maybe due to the amount of fragments taken in the analysis (a total of 24). However, in the three major groups (I, II and III), as can be seen, mix several species of the genus. It is important to highlight that the ISSR technique, that has been considered in various research studies as a strong one, allows a fast estimation, of the inter and intraspecific variability of the organisms that are compared, since this kind of markers is based upon the amplification of the DNA fragments located between two regions of identical microsatellites, which are highly preserved among species.
Sequence analysis
The 5HP and NL4 primers amplified fragments near to 800 bp that belong to the ITS regions in discussion. Sequences were compared to some others registered in the data base of the gene bank of NCBI (National Center for Biotechnology Information, www.ncbi.nih.gov) by means of the BLAST program (Basic Local Alignment Search Tool, http://www.ncbi.nlm.nih.gov/BLAST/). The comparison with the reference accessions showed that most isolates belong to Neofusicoccum sp.; but also D. corticola, L. pseudotheobromae, N. eucalyptorum and N. parvum. were identified. The accession number designated by NCBI to each of the 14 sequences of the organisms of this study are in Table 2.
Table 2 Identification of sequences.
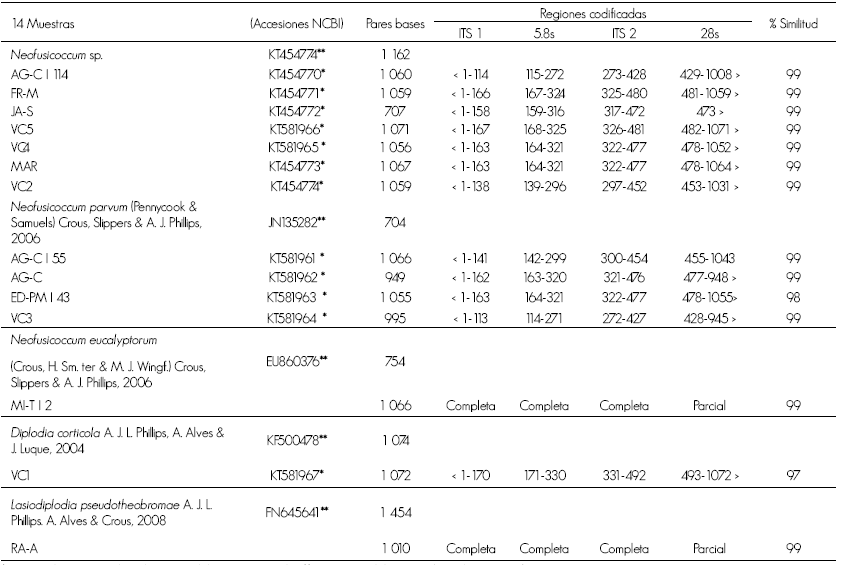
* Accession number of the present isolates in this study; **Accesions of the GenBank used as reference.
In Michoacán State, the morphological and molecular identification of N. parvum and N. eucalyptorum (B. eucalyptorum was recorded in isolates of E. nitens and E. globulus linked to dieback. The genetic sequences of N. eucalyptorum (NCBI numbers of accession: KC479184 and KC4799188) and N. parvum (KC479185, KC479186 and KC479187) showed a similitude of 100 % with those of the actual study, which is the first report of these two species in Mexico which are responsible of the disease (De la Mora et al., 2014). Therefore, the actual study confirms the existence of isolated N. parvum and N. eucalyptorum of damaged eucalyptus trees with canker.
On the other hand, Lasiodiplodia theobromae (Pat.) Griffon & Maubl., 1909 and N. parvum have been found in mango fruits, and the symptom has been linked with dieback and rottenness of the pedunculus of mango in Mexico (Sandoval et al., 2013). Such study is the first survey of species of the Botryosphaeriacea family in this product. L. theobromae, N. mangiferum and N. parvum are regularly related to diseases in the producing regions of mango in the world (Sakalidis et al., 2011). L. pseudotheobromae was reclassified by Alves et al. (2008) as a cryptic species of L. theobromae and they took it as a new species; ever since, it has been recognized in hosts with commercial value such as Citrus spp., Coffea sp., Mangifera spp. and Rosa sp. (Zhao et al., 2010).
As it has been mentioned before, there are difficulties for the right identification and separation among species of Botryosphaeria. Molecular methods are very useful to support this ending, mainly through DNA sequencing and the characterization based on molecular markers have approached to the knowledge of the genomic variability of the organisms in question. But the morphological becomes necessary to complete the identification of such species (Denman et al., 2000; Phillips et al., 2008), and based upon their apparent traits, new ones have been recognized. The tendency of this increase comes from the finding and description of the species of the genus found in different hosts in regard to another already studied (Phillips et al, 2007).
The existence of so many species has been questioned, and, hence, that the number of them could be even smaller, in particular when comparing the DNA sequences. This viewpoint is based on the fact that the mutations of nucleotides are responsible for the huge biological polymorphism that favors evolution; and, since mutations usually occur at random, they can have or not impact upon the final structure of the organism. However, when mutations affect protein coding genes, they provoke morphological polymorphisms (Bruce et al., 2002); an example would be the variation in the color and shape of the mycelium of Botryosphaeria, and maybe it is the major problem that has originated the idea of the so different taxa, even though the DNA sequences suggest the opposite, as it happened with the actual study.
Phylogenetic analysis
The phylogenetic tree constructed with ITS sequences linked the closest isolates based on its similarity (Figure 4). In the phylogram two groups can be observed whose difference is around 0.03 relative distance; the largest group is divided into two subgroups: the first contains VC3, VC4, VC2, FR-M, AG CI 114, JA-S, VC5, MAR, ED PM I 43 and AG C which were grouped with the JN135282 and KJ657705 accessions corresponding to N. parvum and N. sp., respectively; the second subgroup with isolations AG C155 and MI T I 2 grouped with EU860376 accession which belongs to N. eucalyptorum; and finally the smallest group made up of the VC1 and RA A isolates that clustered with FN645641 corresponding to L. pseudotheobromae.
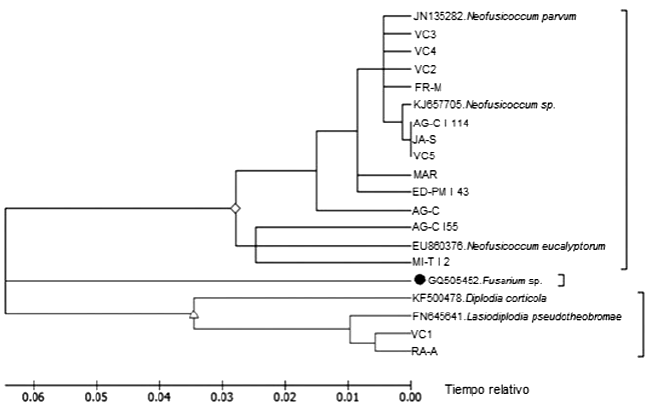
Figure 4 Phylogenetic tree by Maximum Parsimony obtained from sequences of the ITS region of Neofusicoccum sp., D. corticola, L pseudotheobromae, N. eucalyptorum and N. parvum identified in this study. The JN135282, KJ657705, EU860376, GQ505452, KF500478 and FN645641 accessions are representative of the reference species.
Conclusions
The isolates described by the ISSR technique and that belong to the Botryosphaeria genus showed little genomic variability among them, even when five species were involved.
The differences at the nucleotide level that were observed in the studied isolates in regard to the species used as outgroups, were 1-3 %.
The species identified and the incidence of the species associated to cankers and drying of the samples of sick Eucalyptus that were obtained in the 14 analyzed samples were Neofusicoccum sp. 50 %; Neofusicoccum parvum 29 %; Neofusicoccum eucalyptorum, 7 %; Diplodia corticola 7 % and Lasiodiplodia pseudotheobromae 7 %.
Conflict of interests
The authors declare no conflict of interests.
Contribution by author
Dany Gabriel Santiago-Santiago: collection accomplishment, laboratory protocols and writing of the manuscript; Ernestina Valadez Moctezuma and David Cibrián Tovar: funding negotiations to carry out the Project, supervision of the whole work and final version of the manuscript.
Acknowledgements
The first author wants to express his gratitude to the Consejo Nacional de Ciencia y Tecnología for the grant to accomplish his Master of Science degree as well as to the Universidad Autónoma Chapingo for the facilities and financial support received to carry out the actual research study.
REFERENCES
Alves, A., P. W. Crous, A. Correia and A. J. L. Phillips. 2008. Morphological and molecular data reveal cryptic species in Lasiodiplodia theobromae . Fungal Diversity 28: 1-13. [ Links ]
Awasthi, A. K., G. M. Nagarajal, G. V. Naik, S. Kanginakudru, K. Thangavelu and J. Nagaraju. 2004. Genetic diversity and relationships in mulberry (genus Morus) as revealed by RAPD and ISSR marker assays. BMC Genetics 5: 1-8. [ Links ]
Barnett, H. L. and B. B. Hunter. 1998. Illustrated genera of imperfect fungi. The American Phytopathological Society. St Paul, MN, USA. 218 p. [ Links ]
Cibrián, T., D. 2013. Manual para la identificación y manejo de plagas en plantaciones forestales. México: Universidad Autónoma Chapingo- Conafor-Conacyt. México, D.F., México. 230 p. [ Links ]
Comisión Nacional Forestal (Conafor). 2012. Programa de Desarrollo de Plantaciones Forestales Comerciales: A 15 años de su creación. http://www.conafor.gob.mx:8080/biblioteca/ver.aspx?articulo=493 (2 de febrero de 2016). [ Links ]
De la Mora C., J. G., D. Cibrián T. y O. A. Pérez V. 2014. Neofusicoccum eucalyptorum (=Botryosphaeria eucalyptorum ) y N. parvum: patógenos en plantaciones de eucalipto en México. Revista Chapingo. Serie Ciencias Forestales y del Ambiente 20(3): 187-197. [ Links ]
Denman, D., P. C. Crous, J. E. Taylor, K. Ji-Chuan, I. Pascoe and M. J. Wingfield. 2000. An overview of the taxonomic history of Botryosphaeria , and a re-evaluation of its anamorphs based on morphology and ITS rDNA phylogeny. Studies in Mycology 45: 129-140. [ Links ]
Jin, Y., T. He and B. R. Lu. 2003. Fine scale genetic structure in a wild soybean (Glycine soja ) population and the implications for conservation. New Phytologist 159:513-519. [ Links ]
Mohali, S., B. N. Slippers and M. J. Wingfield. 2006. Two new Fusicoccum species from Acacia and Eucalyptus in Venezuela based on morphology and DNA sequence data. Mycological Research 110 (4):405-413. [ Links ]
Pérez, C. A., M. J. Wingfield, B. Slippers, N. A. Altier and R. A. Blanchette. 2009. Neofusicoccum eucalyptorum , an Eucalyptus pathogen on native Myrtaceae in Uruguay. Plant Pathology 58: 964-970. [ Links ]
Pérez, C. A. , M. J. Wingfield , B. Slippers , N. A. Altier and R. A. Blanchette. 2010. Endophytic and canker-associated Botryosphaeriaceae occurring on non-native Eucalyptus and native Myrtaceae trees in Uruguay. Fungal Diversity 41: 53-69. [ Links ]
Phillips, A. J. L., A. Alves, S. R. Pennycook, P. R. Johnston, A. Ramaley, A. Akulov and P. W. Crous. 2008. Resolving the phylogenetic and taxonomic status of dark-spored teleomorph genera in the Botryosphaeriaceae. Persoonia 21: 29-55. [ Links ]
Phillips, A. J. L. , P. W. Crous and A. Alves . 2007. Diplodia seriat a, the anamorph of "Botryosphaeria " obtusa . Fungal Diversity 25: 141-155. [ Links ]
Sakalidis, M. L., G. E. St. J. Hardy and T. I. Burguess. 2011. Use of the Genealogical Sorting Index (GSI) to delineate species boundaries in the Neofusicoccum parvum-Neofusicoccum ribis species complex. Molecular Phylogenetics and Evolution 60(320): 333-344. [ Links ]
Sakalidis, M. L. , J. D. Ray, V. Lanoiselet, G. E. St J. Hardy and T. I. Burgess. 2011. Pathogenic Botryosphaeriaceae associated with Mangifera indica in the Kimberley Region of Western Australia. European Journal of Plant Pathology 130: 379-391. [ Links ]
Sandoval. Z. M., A. D. Nieto, S. J. Sandoval I., O. D. Téliz, S. M. Orozco y V. H. Silva-Rojas. 2013. Hongos asociados a pudrición del pedúnculo y muerte descendente del mango (Mangifera indica L.). Agrociencia 47: 61-73. [ Links ]
Sivanesan, A. 1984. The bitunicate Ascomycetes and their anamorphs. Lubrecht & Cramer, Ltd. Port Jervis, NY, USA. 701 p. [ Links ]
Slippers, B and M. J. Wingfield . 2007. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: diversity, ecology and impact. Fungal Biology Reviews 21: 90-06. [ Links ]
Smith, H., P. W. Crous, M. J. Wingfield , T. A. Coutinho and B. D. Wingfield. 2001. Botryosphaeria eucalyptorum sp. nov., a new species in the B. dothidea -complex on Eucalyptus in South Africa. Mycologia 93(2): 277-285. [ Links ]
Velázquez M., A., A. M. Fierros G., A. Aldrete, G. A. Gómez, C. S. Férnández, H. De los Santos y H. Ramírez C. 2013. Situación actual y perspectivas de las plantaciones forestales comerciales en México. Comisión Nacional Forestal. Guadalajara, Jal., México. 429 p. [ Links ]
Weising, K., H. Nybom, M. Pfenninger, K. Wolff and G. Kahl. 2005. DNA fingerprinting in plants: principles, methods and applications. CRC press. Boca Raton, FL, USA. pp. 235-276. [ Links ]
Zhao, J. P., Q. Lu, J. Liang, C. Decock and X. Y. Zhang. 2010. Lasiodiplodia pseudotheobromae , a new record of pathogenic fungus from some subtropical and tropical trees in southern China. Cryptogamie Mycologie. 31: 431-439. [ Links ]
Zizumbo V., D. P. Colunga G. M., E. P. de la Cruz, P. Delgado V. and P. Gepts. 2005. Population structure and evolutionary dynamics of wild-weedy domesticated complexes of common bean in a Mesoamerican region. Crop Science 45:1073-1083. [ Links ]
Received: September 24, 2015; Accepted: November 12, 2015
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
