











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El estado de Quintana Roo cuenta con selvas bajas caducifolias, medianas subcaducifolias y subperennifolias (Miranda y Hernández, 1963; Pennington y Sarukhán, 2005), en las cuales se realiza silvicultura desde hace aproximadamente 60 años (Negreros-Castillo et al., 2014). Se estima que el aprovechamiento forestal de estas selvas además de las reservas naturales son los factores que permiten la permanencia y ganancia de superficie arbolada en el estado (Ellis, Hernández y Romero-Montero, 2017), por lo que se debe procurar su continuidad de manera adecuada y sostenible (Negreros-Castillo y Mize, 2013).
La caoba (Swietenia macrophylla King) y el cedro (Cedrela odorata L.) son las especies más valoradas en Quintana Roo y el avance en el conocimiento de su ecología y silvicultura ha sido sustancial (Snook, 2005; Negreros-Castillo y Mize, 2013; Negreros-Castillo et al., 2014). Bursera simaruba (L.) Sarg., Lysiloma latisiliquum (L.) (Benth.) y Piscidia piscipula (L.) Sarg. son otras especies aprovechadas en el estado, cuyos volúmenes maderables llegan a ser tan importantes como los de caoba y cedro, por lo que es idóneo ampliar el conocimiento ecológico y técnico sobre ellas para conservar la composición, la estructura y el funcionamiento de las selvas en que se desarrollan (Negreros-Castillo y Martínez-Salazar, 2011; Negreros-Castillo et al., 2014).
Para aprovechar cualquier especie leñosa es necesario conocer su tasa de crecimiento, así como los múltiples factores que en ella inciden, por ejemplo: el ambiente, la fisiología o la morfología. Estimar el crecimiento también tiene implicaciones en: prospección del cambio climático (Way y Oren, 2010), competencia ecológica (O’Connor, Lynch, Falk y Swetnam, 2015), captura de carbono (Vanderwel, Zeng, Caspersen, Kunstler y Lichstein, 2016), dinámica de poblaciones (Fassnacht et al., 2015) y restauración ecológica (Fahey y Lorimer, 2013).
En la península de Yucatán los antecedentes de crecimiento sobre estas especies son los trabajos de Mize, Centeno y Negreros-Castillo (1997)), en selvas subperennifolias de Campeche, que concluyen que los incrementos de fuste no fueron diferentes entre tres tipos de suelo (Litosol, Rendzina, Vertisol) ni entre edades del bosque (18 años, 24 años, 29 años y 30 años). Para L. latisiliquum las investigaciones de López-Torres y Tamarit-Urias (2005), también en Campeche, así como las de García y Rodríguez (1993) y Negreros-Castillo y Martínez-Salazar (2011) en Quintana Roo mencionan que la apertura del dosel y cortas intermedias de mejoramiento inducen mayores incrementos en diámetro.
La complejidad del proceso del crecimiento vegetal no puede adjudicarse únicamente a una variable, sino a una serie de factores abióticos y bióticos (Baker, Swaine y Burslem, 2003). Algunos estudios señalan que el clima ocuparía el primer lugar, las características del micrositio el segundo y, por último, la naturaleza e intensidad del disturbio que regeneró al bosque (Toledo et al., 2011). Sin embargo, la estructura, la composición de otras selvas y las características bioecológicas de cada especie no permitirían extrapolar este orden.
Pocos estudios han establecido la relación clima-crecimiento en la Península de Yucatán (Interián-Ku, Vaquera-Huerta, Valdez-Hernández, García-Moya y Romero-Manzanares, 2014; Estrada-Medina, Santiago, Graham, Allen y Jiménez-Osornio, 2013) y los resultados indican que el elemento precipitación ocupa un lugar primario en este proceso. Investigaciones de mayor profundidad sobre la temperatura, la humedad, la radiación solar, etc., sobre el crecimiento arbóreo son incipientes en México (Yáñez-Espinosa, Terrazas y López-Mata, 2006; Ricker, Gutiérrez-García y Daly, 2007). La importancia de incluir a la temperatura y/o a la humedad radica en que intervienen en el inicio de la foliación y la consecuente activación de la actividad del cambium (Lloyd y Farquhar, 2008; Way y Oren, 2010).
Por lo anterior, es probable que el crecimiento de otras especies de las selvas de Quintana Roo esté relacionado no únicamente con la lluvia y su distribución, sino que otros elementos pueden tener una influencia de la misma o mayor magnitud.
Objetivos
Determinar el crecimiento en diámetro de Bursera simaruba, Lysiloma latisiliquum y Piscidia piscipula en una selva secundaria subperennifolia y correlacionarlo con la precipitación, la temperatura y la humedad relativa.
Materiales y métodos
Área de estudio
El sitio de estudio se localiza en el ejido Andrés Quintana Roo del municipio Felipe Carrillo Puerto, Quintana Roo (19° 58’ 5.2” N, 87° 53’ 12.2” O). Geomorfológicamente es una planicie caracterizada por bancos de yeso de varios metros de espesor intercalados entre rocas calizas cristalinas y margas blancas o amarillas (Pozo, Armijo y Calmé, 2011). Los suelos, según la clasificación maya, son los denominados tzequel (litosoles) con una profundidad menor de 15 cm con afloración de rocas de color negro con presencia intercalada de yaaxhom (vertisoles gleicos o gleisoles) de profundidad variable y de color negro a gris y kankab (rendzinas) de color rojo a negro con profundidades variables (Organización de Ejidos Productores Forestales de la Zona Maya, S.C. [Oepfzm], 2000; Organización Mundial de las Naciones Unidas para la Agrigultura y la Alimentación [FAO], 2007). El clima es de tipo Aw1, cálido subhúmedo con lluvias en verano, con temperatura media anual de 25.8 °C y una precipitación media anual de 1204.5 mm (García, 1987).
El tipo de vegetación es selva mediana subperennifolia que conserva de 50% a 75% su follaje durante la época seca, con algunas lluvias en la temporada seca (Miranda y Hernández, 1963). La altura del dosel varía de 15 m a 25 m dependiendo del tipo de suelo y especie arbórea presente. Los árboles dominantes son: Metopium Browne Jacq. Urb., Manilkara zapota (L.) P. Royen, Bursera simaruba (L.) Sarg. y Lysiloma latisiliquum (L.) (Benth.); otras arbóreas: Nectandra spp. Rol. ex Rottb., Byrsonima bucidaefolia Standl., Caesalpinia gaumeri Greenm., Piscidia piscipula (L.) Sarg., Piscidium sartorianum (O. Berg) Nied., Vitex gaumer Greenm.y Beaucarnea ameliae Lem. y las palmas: Thrinax radiata Lodd. ex Schultz. &Schult., Coccothrinax readii H. J. Quero, Pseudophoenix sargenti H. Wendl. ex Sarg. (Miranda y Hernández, 1963; Pennington y Sarukhán, 2005).
Muestreo de árboles y medición del diámetro
En dos unidades de muestreo de 1000 m2 (20 m × 50 m), ambos con una temporalidad de sucesión de 30 años y cuyo uso anterior fue el ganadero, se eligieron: 73 árboles (> 2.5 cm DAP): 23 de Bursera simaruba, 25 de Lysiloma latisiliquum y 25 de Piscidia piscipula. Todos los individuos seleccionados presentaron fuste recto. Fueron clasificados por categoría de diámetro: 5 = 2.5 cm - 7.49 cm; 10 = 7.5 cm - 12.49 cm; 15 = 12.5 cm - 17.49 cm; 20 = 17.5 cm - 22.49 cm (López-Ayala, Valdez-Hernández, Terrazas y Valdez-Lazalde, 2006).
A todos los árboles se les colocaron bandas dendrométricas de acero inoxidable fabricadas en el lugar (Cattelino, Becher y Fuller, 1986). De enero de 2010 a mayo de 2011 se realizaron medidas bimestrales.
La precipitación, temperatura promedio y la humedad relativa (Tabla 1) para el periodo de medición se obtuvieron de la estación de Felipe Carrillo Puerto (19° 34’ 32” N, 88° 02’ 42” O) del Servicio Meteorológico Nacional.
Tabla 1 Precipitación, temperatura y humedad relativa para la zona de estudio durante el periodo de medición
| Fecha | Precipitación (mm) | Temperatura (°C) | Humedad relativa (%) |
|---|---|---|---|
| enero 2010 | 19.79 | 22.17 | 79.96 |
| marzo 2010 | 10.4 | 23.82 | 71.71 |
| mayo20 10 | 345.13 | 28.27 | 82.64 |
| julio 2010 | 392.5 | 27.38 | 91.1 |
| septiembre 2010 | 124.38 | 27.62 | 88.68 |
| noviembre 2010 | 11.18 | 23.73 | 82.27 |
| enero 2011 | 31.98 | 22.89 | 84.17 |
| marzo 2011 | 122.4 | 24.94 | 80.22 |
| mayo 2011 | 1.77 | 28.74 | 74.97 |
Ecuaciones de crecimiento
Para modelar el crecimiento acumulado en función del tiempo, se empleó el siguiente modelo polinomial de tercer orden (Kiviste, González, Alboreca, y González, 2010):
La ecuación fue evaluada a través de técnicas de regresión no lineal en el programa Curve Expert (Hyams, 2005). Se empleó el método de mínimos cuadrados generalizados, en los cuales se comprobaron los supuestos de normalidad y homogeneidad de varianzas (Guerra, Cabrera y Fernández, 2003). Los indicadores de ajuste fueron: cuadrado medio del error (CME), coeficiente de determinación (R2), y nivel de significancia estadística (p ≤ 0.01) de los parámetros de cada ecuación (Guerra et al., 2003).
Análisis estadístico
Para comparar el crecimiento entre categorías diamétricas se realizó un análisis de varianza (ANDEVA) (Gotelli y Ellison, 2004) y una prueba de comparación múltiple (Bonferroni) (Gotelli y Ellison, 2004). La precipitación, la temperatura y la humedad relativa se correlacionaron con los incrementos en diámetro (Pearson) (Gotelli y Ellison, 2004). El paquete estadístico empleado fue Stata versión 9.1 (Stata Corp, 2001).
Resultados
Las curvas de incremento general de las tres especies mostraron valores altos de R2 y niveles de confianza estadística aceptables. Lysiloma latisiliquum fue la que tuvo mayor aumento en diámetro, seguida por Piscidia piscipula y Bursera simaruba. La curva de L. latisiliquum tuvo la mayor pendiente respecto a las otras dos. En P. piscipula se aprecianmomentos de menor incremento, lo cual se relaciona con la época de lluvias (verano de 2010). B. simaruba presentó la curva con la menor pendiente, aunque con ritmo sostenido durante todo el estudio (Fig. 1).
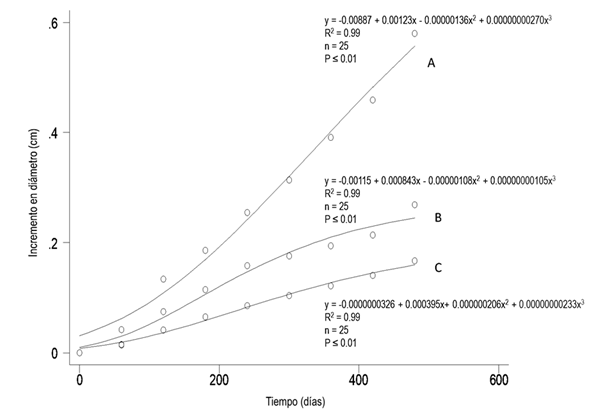
Figura 1. Curvas de crecimiento acumulado. A. Lysiloma latisiliquum. B. Piscidia piscipula. C. Bursera simaruba. Los círculos son los incrementos promedio por cada medición, la línea representa la curva ajustada.
La tabla 2 muestra los incrementos que presentaron las tres especies, confirmándose que Lysiloma latisiliquum fue superior en máximo, promedio y mínimo.
Tabla 2 Incrementos anuales, máximo, promedio, mínimo y desviación estándar de tres especies de la selva mediana subperennifolia de Quintana Roo, México.
| Especie | Incremento (cm) | Desviación estándar | ||
|---|---|---|---|---|
| Máximo | Promedio | Mínimo | ||
| Bursera simaruba | 0.1669 | 0.0615 | 0.0137 | 0.0537 |
| Lysiloma latisiliquum | 0.5793 | 0.1884 | 0.0416 | 0.1820 |
| Piscidia piscipula | 0.2688 | 0.1044 | 0.0149 | 0.0859 |
Entre las categorías diamétricas de Bursera simaruba, la 20 mostró los mayores incrementos, seguida de 15, 10 y 5. En dicha categoría se presentaron los individuos con los mayores aportes de incremento en diámetro de la muestra. Los valores estadísticos del modelo mostraron valores de R2 medianos en las categorías 5 y 10 y valores bajos en en las categorías 15 y 20 (Fig. 2).

Figura 2 Curvas de incremento acumulado de las categorías diámetricas de Bursera simaruba. A = categoría 5; B = categoría 10; C = categoría 15 y D = categoría 20. Los círculos son los incrementos por cada medición de cada árbol, la línea representa la curva ajustada.
La categoría 20 de Lysiloma latisiliquum presentó los mayores incrementos, seguida por 15, 5 y 10. Los valores de R2 fueron bajos debido a que en las cuatro categorías se presentaron individuos con incrementos destacados en la muestra (Fig. 3).

Figura 3 Curvas de incremento en diámetro de las categorías diámetricas de Lysiloma latisiliquum. A = categoría 5; B = categoría 10; C = categoría 15 y D = categoría 20. Los círculos son los incrementos por cada medición de cada árbol, la línea representa la curva ajustada.
Los incrementos en la categoría 15 fueron mayores a los de 20, 10 y 5 en Piscidia piscipula. Los valores de R2 fueron medianos dado que se presentaron individuos con incrementos destacados en las categorías 15 y 20 (Fig. 4).
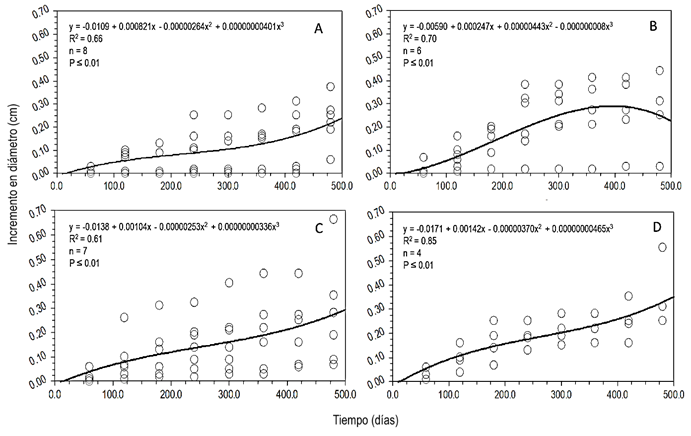
Figura 4 Curvas de incremento en diámetro de las categorías diámetricas de Piscidia piscipula. A = categoría 5; B = categoría 10; C = categoría 15 y D = categoría 20. Los círculos son los incrementos por cada medición de cada árbol, la línea representa la curva ajustada.
En la tabla 3 se muestran los incrementos promedio y totales por categoría diamétrica de las tres especies. Los incrementos de las categorías de Lysiloma latisiliquum superan a los de Piscidia piscipula y de Bursera simaruba en ese orden. La categoría con los mayores incrementos en las tres especies fue la 20 y aquellos incrementos con los menores valores correspondieron a las categorías 5, 10 y 15 para B. simaruba, L. latisiliquum y P. piscipula, respectivamente. El análisis de varianza, mostró que en las tres especies se encontraron diferencias significativas entre las categorías diamétricas de las tres especies, dichas variaciones fueron más significativas en Bursera simaruba, seguidas por Piscidia piscipula y por último en Lysiloma latisiliquum (Tabla 4).
Tabla 3 Incrementos en diámetro por categoría diamétrica de tres especies de la selva mediana subperennifolia de Quintana Roo, México.
| Especie | Categoría diamétrica (cm) | ||||
|---|---|---|---|---|---|
| 5 | 10 | 15 | 20 | Promedio | |
| Bursera simaruba | 0.0535 | 0.0764 | 0.0720 | 0.1197 | 0.0804 |
| Lysiloma latisiliquum | 0.1958 | 0.1848 | 0.3304 | 0.3009 | 0.2530 |
| Piscidia piscipula | 0.0949 | 0.1664 | 0.1344 | 0.1679 | 0.1409 |
Tabla 4 Análisis de varianza de tres especies de la selva mediana subperennifolia de Quintana Roo, México.
| Especie | Fuente de variación | SC | GL | MS | F | Prob > F |
|---|---|---|---|---|---|---|
| Bursera simaruba | Entre grupos | 0.140257091 | 3 | 0.046752364 | 6.39 | 0.0004 |
| Intragrupos | 1.61594959 | 221 | 0.007311989 | |||
| Total | 1.75620668 | 224 | 0.007840208 | |||
| Lysiloma latisiliquum | Entre grupos | 0.923860397 | 3 | 0.307953466 | 2.80 | 0.0408 |
| Intragrupos | 24.2889905 | 221 | 0.109904934 | |||
| Total | 25.2128509 | 224 | 0.11255737 | |||
| Piscidia piscipula | Entre grupos | 0.207918553 | 3 | 0.069306184 | 4.18 | 0.0066 |
| Intragrupos | 3.66105275 | 221 | 0.01656585 | |||
| Total | 3.86897131 | 224 | 0.017272193 |
SC = Suma de cuadrados, GL = Grados de libertad, MS = Cuadrados medios, F = Valor de F; Prob > F = Nivel de confiabilidad.
Entre las categorías diamétricas de Bursera simaruba la 20 presentó mayor crecimiento que el resto. Para Lysiloma latisiliquum la diferencia se encontró entre las categorías 10 y 20. Piscidia piscipula presentó los mayores crecimientos en las categorías de 10 y 20, mientras que los menores se dieron en las categorías 5 y 15 (Tabla 5).
Tabla 5 Prueba de comparación múltiple de Bonferroni para categorías diamétricas de tres especies de la selva mediana subperennifolia de Quintana Roo, México.
| Especie | Categoría diamétrica | |||
|---|---|---|---|---|
| 5 | 10 | 15 | 20 | |
| Bursera simaruba | 0.0535a | 0.0764a | 0.0720a | 0.1197b |
| Lysiloma latisiliquum | 0.1958a | 0.1848b | 0.3304a | 0.3009c |
| Piscidia piscipula | 0.0949a | 0.1664b | 0.1344a | 0.1679c |
Valores con letras distintas son estadísticamente diferentes (P≤ 0.05)
La precipitación y la humedad relativa solo se correlacionaron con el grosor del fuste de la categoría diamétrica 10 en Bursera simaruba y Piscidia piscipula, respectivamente. La temperatura fue el elemento que presentó correlaciones positivas y significativas para las tres especies, principalmente con Piscidia piscipula y Bursera simaruba (Tabla 6).
Tabla 6 Correlaciones paramétricas (Pearson) de los incrementos en diámetro de tres especies de la selva mediana subperennifolia de Quintana Roo, México y los elementos del clima.
| Especie | Categoría diamétrica | Precipitación | Sig. | Temperatura | Sig. | Humedad relativa | Sig. |
|---|---|---|---|---|---|---|---|
| Bursera simaruba | 5 | -0.1367 | ns | 0.6453 | ns | -0.3072 | ns |
| 10 | 0.6604 | * | 0.7699 | * | 0.2033 | ns | |
| 15 | 0.3531 | ns | 0.7259 | * | 0.0362 | ns | |
| 20 | 0.5804 | ns | 0.6018 | ** | 0.5028 | ns | |
| Todas | 0.5036 | ns | 0.8269 | ** | 0.2309 | ns | |
| Lysiloma latisiliquum | 5 | -0.1066 | ns | 0.6168 | ns | -0.0035 | ns |
| 10 | -0.0206 | ns | 0.2021 | ns | -0.102 | ns | |
| 15 | 0.343 | ns | 0.5944 | ns | 0.1606 | ns | |
| 20 | 0.0181 | ns | 0.6366 | ns | -0.1823 | ns | |
| Todas | 0.1159 | ns | 0.7018 | * | -0.0458 | ns | |
| Piscidia piscipula | 5 | 0.123 | ns | 0.822 | ** | -0.2145 | ns |
| 10 | 0.5791 | ns | 0.2869 | ns | 0.7437 | * | |
| 15 | 0.3307 | ns | 0.8154 | ** | -0.0347 | ns | |
| 20 | 0.4816 | ns | 0.8754 | ** | -0.0352 | ns | |
| Todas | 0.5472 | ns | 0.9637 | *** | 0.2304 | ns |
Sig. Significancia;***P ≤ 0.001; **P ≤ 0.01; *P ≤ 0.05; ns = no significativo
Discusión
El orden de los incrementos entre especies en el presente estudio coincidió con lo encontrado por Mize et al. (1997) en Campeche (1000 mm/año - 1200 mm/año) donde Lysiloma latisiliquum fue la especie con mayor aumento en el grosor de su fuste y Bursera simaruba con el menor. Dicho estudio se realizó en rodales de diferentes edades (18 años, 24 años, 29 años, 30 años y > 30 años) de una selva secundaria subperennifolia con tipos de suelo similares (litosoles, rendzinas, luvisoles) a los del presente trabajo.
En la investigación de Mize et al. (1997), las tasas anuales de crecimiento también fueron superiores (B. simaruba: 0.38 cm - 0.61 cm; L. latisiliquum: 0.78 cm - 1.0 cm; P. piscipula: 0.40 cm - 0.61 cm) a las encontradas en el presente trabajo. Baker et al., (2003) mencionan que diferencias entre sitios son resultado de una serie concatenada de hechos como el uso de recursos abióticos por parte de la especie, el clima, la apertura y tamaño de claros, así como el vigor genético de cada población.
El comportamiento de B. simaruba en el presente estudio coincide con lo mencionado por López-Ayala et al. (2006) en una selva mediana subcaducifolia de Colima, México respecto al patrón uniforme de la curva de crecimiento y a la relación del aumento en grosor del fuste en función del tamaño de los individuos. Hulshof, Stegen, Swenson, Enquist y Enquist (2012) mencionan que en B. simaruba las diferencias de crecimiento entre tamaños se deben a la disponibilidad de recursos abióticos interanuales y a la variabilidad entre micrositios.
En el caso del sitio de estudio, las bajas tasas de crecimiento - principalmente de B. simaruba - corresponden al de un bosque con sucesión secundaria cuya densidad de individuos requiere de tratamientos silvícolas de liberación que favorezcan la apertura del dosel, debido a ello es que los árboles de la mayor categoría tienen incrementos destacados ya que son aquellos que han alcanzado el dosel.
Lo mencionado por López-Ayala et al. (2006), respecto al papel del clima en el crecimiento de B. simaruba, no coincide con este trabajo debido a que en Colima se encontraron correlaciones negativas con la temperatura y positivas con la precipitación, sin llegar a ser significativos ambos casos. Valdez-Hernández, Andrade, Jackson y Rebolledo-Vieyra (2010), en una selva baja caducifolia de Yucatán, encontraron que B. simaruba presentó sus mayores intensidades de foliación en la época de secas y lluvias, coincidiendo con el aumento de la temperatura. Lo anterior permite inferir que la presencia de follaje permite actividad fotosintética que induciría la activación del cambium vascular y por ende un aumento en el diámetro (Pallardy, 2008).
Los trabajos de López-Torres y Tamarit-Urias (2005) en Campeche (1390 mm año-1) y de Negreros-Castillo y Martínez-Salazar (2011) en Quintana Roo (1300 mm año-1) registraron tasas de crecimiento en diámetro de L. latisiliquum mayores (0.51 cm/año y 0.41 cm/año, respectivamente) a los del presente trabajo. Ambas investigaciones demostraron que las tasas de crecimiento están en función del tamaño progresivo del DAP, lo cual coincide parcialmente con esta investigación. La evidencia científica permite establecer que árboles que se encuentran en una fase de curva ascendente y que no han llegado a la etapa de plena madurez pueden presentar variación entre sus categorías diamétricas, respecto a sus tasas de crecimiento (Vanclay, 1995).
La relación del clima con el crecimiento de Lysiloma sp. fue estudiado por Daubenmire (1972), quien únicamente señalo que no había decrementos en patrón radial por efecto de la precipitación, por lo que este trabajo proporciona un mayor acercamiento sobre la influencia de la temperatura. Valdez-Hernández et al. (2010) encontraron, en dos especies de Fabaceas (Acacia gaumeri S. F. Blake, Apoplanesia paniculata C. Presl.), un patrón de foliación coincidente con el termoperiodo de la zona, lo que muestra la importancia de fenología en el proceso de crecimiento.
Los incrementos de P. piscipula en el presente estudio difieren de lo mencionado por Verduzco (2016) en Yucatán, donde se menciona que no se presentaron diferencias significativas entre dos categorías diamétricas (juveniles: 5 cm - 10 cm, maduros DAP > 20 cm). Baker, Swaine y Burslem (2003) sugieren que variables como el micrositio y el uso particular de recursos por parte de la especie definen los patrones de crecimiento.
De acuerdo con los resultados de Verduzco (2016), P. piscipula engrosó su fuste en función de un gradiente de precipitación distribuido en tres sitios (677.9 mm, 1132.0 mm y 1439.6 mm) lo cual no coincide con lo encontrado en Felipe Carrillo Puerto, Quintana Roo. La evidencia ecofisiológica sobre la especie en la Península de Yucatán menciona que P. piscipula ejerce una estrategia oportunista en el aprovechamiento del agua, con lo cual mantiene el potencial hídrico de forma constante durante la estación seca (Estrada-Medina et al., 2013), lo cual permite inferir que la temperatura es un elemento climático simultáneo en la activación de la actividad del cambium.

