











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroduction
Coffee is an important crop, and its world production is based on only two species: Coffea canephora P. (robusta) and Coffea arabica L. (arabica), of which C. arabica is the dominant species because of the volume of global production it represents (63 %) and because it is associated with better quality coffee. World production in 2017 and 2018 was approximately 164,810 60-kg bags, where arabica had a 2.2 % increase in production, and robusta, 11.7 % (International Coffee Organization [ICO], 2019).
C. arabica is an allotetraploid (2n = 4x = 44) and autogamous species, which was derived from a spontaneous hybridization between two closely related diploid species: Coffea eugenioides S. Moore ( Yuyama et al., 2016) and C. canephora (Lashermes et al., 1999). C. canephora (2n = 2x = 22) is a diploid and alogamous species harboring high diversity (Musoli et al., 2009). The propagation history of C. arabica, combined with its autogamy, has led to narrow genetic diversity within cultivars (Steiger et al., 2002). Likewise, coffee production is limited by several diseases and pests, drought, heat stress, salinity, and climate change (Breitler et al., 2018; dos Santos et al., 2011; Silva et al., 2006). As a result, C. arabica breeding programs are focused on overcoming these obstacles with the use of divergent and wild cultivars, but are limited by the wide time ranges required (Sant’Ana et al., 2018; Tran, Ramaraj, Furtado, Slade, & Henry, 2018).
In this context, new methods that generate variability and reduce the development time of new cultivars are required to address the challenges presented by climate change to coffee cultivation. These methods include the use of traditional breeding schemes, genetic engineering, and induced mutations (Dada et al., 2018). Gamma-ray-induced mutation is one of the potentially useful applications in agriculture (Majeed, Muhammad, Ullah, & Ali, 2018). Gamma radiation from radioactive cobalt (60Co) has high penetrating potential; however, it can be used to irradiate whole plants and their sensitive parts (Oladosu et al., 2016).
In economically important crops (Oryza sativa, Zea mays L., Solanum tuberosum and Phaseolus vulgaris L.), Singh and Datta (2010) note that one of the important physical agents is gamma radiation, which is used to improve agronomic traits and productivity. In these cases, seeds are commonly used to generate mutations because they can tolerate this physical condition (Amir et al., 2018). Kiong, Ling, Lai, Hussein, and Harun (2008) observed that seed germination rate depends on the level of chromosomal damage caused by increasing irradiation doses.
Although mutation induction techniques for plant breeding are successfully applied in a wide range of crops, their use is less frequent in coffee (Dada et al., 2018). Therefore, the objective of this study was to irradiate C. arabica seeds of the Geisha, Oro Azteca and Marsellesa varieties with gamma rays to determine the median lethal dose (LD50) and evaluate their physiological response in terms of germination, survival, plant height (PH), stem diameter (SD), height to first leaf pair (HFLP) and leaf area (LA).
Materials and methods
Plant material, experiment location and experimental design
Seeds of three C. arabica varieties (Geisha, Oro Azteca and Marsellesa) were obtained from the National Coffee Germplasm Bank operated by the Universidad Autónoma Chapingo’s (UACh) Centro Regional Universitario Oriente (CRUO), located in Huatusco, Veracruz, Mexico (19° 10’ 27” N and 96° 57’ 50” W, at 1,345 m a. s. l.). Seed irradiation was performed at the Instituto Nacional de Investigaciones Nucleares (ININ) located in Salazar, State of Mexico, Mexico. The irradiator used was the Transelektro LGI-01, with a dose rate of 752.76 Gy·h-1. The 60Co gamma ray doses used were 0, 100, 200, 300, 400 and 500 Gy, on 700 seeds of each variety harvested in the 2017-2018 cycle, which had 10 % moisture. The control seeds of each genetic material were not irradiated.
The irradiated seeds were established in a glass-covered greenhouse with a steel structure and lateral ventilation on May 15 and 16, 2018, in Montecillo, Texcoco, State of Mexico. The average temperature was 29.0 °C and relative humidity was 27.10 %. The seeds were sown manually in wooden beds (1.5 x 3 x .25 m) with soil. Furrows were established at distances of 5 cm between rows and 1.5 cm between seeds. The beds were checked regularly for water requirements.
The experiment was established according to a completely randomized factorial design with two factors (variety, with three levels, and irradiation dose, with six levels), resulting in 18 treatments with three replicates (225 seeds per replicate).
Evaluated parameters
To determine the effect of the treatments, the variables evaluated were: germination (%), survival (%) and morphological variables of each variety. Data recording for germination was carried out 20 days after sowing (das). Survival and the rest of the variables were determined at 120 das. For the morphological variables, 10 plants were randomly selected per experimental unit, in which PH (cm), HFLP (cm), SD (mm; at 1 cm distance between the base of the root and the first leaf of the plant) and LA (cm2; with the ImageJ program) were measured.
Statistical analysis
The statistical model used was Y ijk = µ + A i +Bj + AB ij + ɛ ijk ; where Y ijk is the value of the response variable corresponding to replicate k of level i of A (variety) to level j of B (dose [Gy]), µ is the overall mean, A i is the effect of level i of A, B i is the effect of level j of B, AB ij is the AxB interaction (corresponding to level i of A and level j of B) and ε ijk is the experimental error corresponding to replicate k of level i of A to level j of B. Using SAS version 9.4 statistical software (SAS Institute, 2002), an analysis of variance was performed for each variable and treatment means were compared using Tukey’s test (P ≤ 0.05). The data obtained from the survival percentage were used to determine the median lethal dose (LD50) of each variety, which was estimated using a simple linear regression model.
Results and discussion
Germination and plant survival percentage
The three varieties showed a negative effect on germination starting at the 100 Gy dose (Figure 1). A 60 % decrease in germination was observed for the Geisha variety with respect to its control, as well as a significant decrease starting at doses of 200 and 400 Gy for the Oro Azteca and Marsellesa varieties, respectively. These results agree with those reported by Quintana et al. (2019), who observed a drastic decrease in the germination of Tipica variety coffee seeds at doses of 100 and 150 Gy. In other species, Kumar, Nepolean, and Gopalan (2003) showed that Phaseolus lunatus seed germination decreased with increasing gamma irradiation in a range of 200-1,000 Gy. Cheema and Atta (2003) showed that the seed germination of three different varieties of Oryza sativa decreased with an increasing irradiation dose. Therefore, exposure to high doses of gamma radiation often inhibits the germination of gymnosperm and angiosperm seeds (Thapa, 2004).
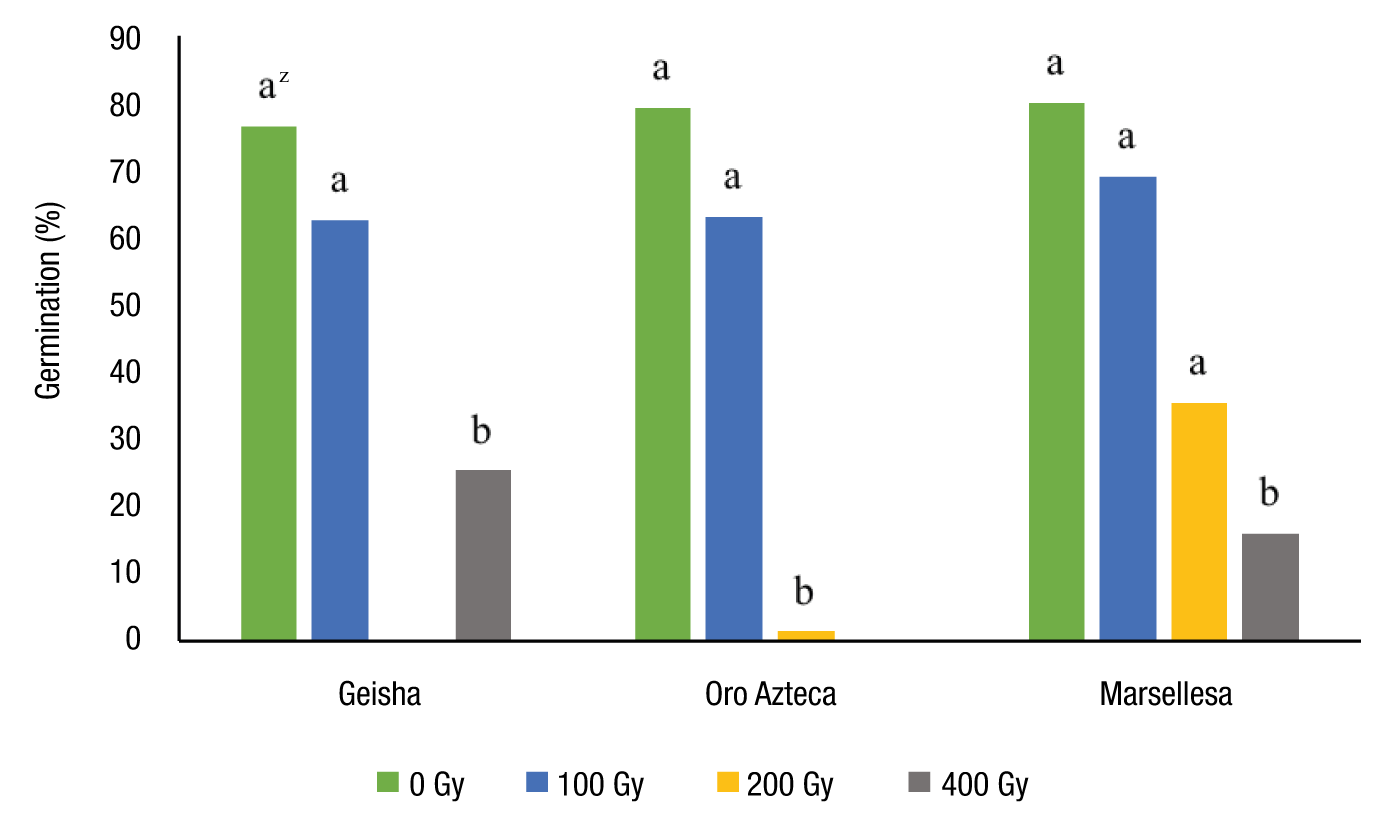
Figure 1 Effect of gamma radiation (60Co) on the germination percentage of the Geisha, Oro Azteca and Marsellesa varieties of C. arabica. zBars with the same letter do not differ statistically (Tukey, P ≤ 0.05).
The survival percentage was significantly affected (P ≤ 0.05) by the gamma radiation (Figure 2). There was a significant decrease starting at the 100 Gy dose in all three varieties compared to their respective controls. A similar trend was described by Ramírez, González, Camejo, Zaldívar, and Fernández (2006), who observed that doses higher than 200 Gy decreased the survival percentage of S. lycopersicum L. seedlings. These authors observed that, although radicle emission and cotyledon emergence occurred a few days after germination and with slow growth, the seedlings finally died. Similarly, it was shown that with doses of 600 Gy in Jatropha curcas L. seeds, plant survival decreased significantly (Sonsgri, Suriharn, Sanitchon, Srisawangwong, & Kesmala, 2011).

Figure 2 Effect of gamma radiation (60Co) on the survival percentage of the Geisha, Oro Azteca and Marseillaise varieties of C. arabica. zBars with the same letter do not differ statistically (Tukey, P ≤ 0.05).
A general indicator defining the range of mean ionizing radiation (IR) doses is a short-term activation period preceding the onset of inhibition of physiological processes. Mean IR doses range from 20 to 100 Gy (irradiation of seeds) or from 5 to 50-70 Gy (irradiation of plant vegetative parts); therefore, doses above these levels are called high (Gudkov, Grinberga, Sukhova, & Vodeneeva, 2019). Broadly speaking, mutagenic agents used to induce hereditary changes are divided into physical and chemical groups, of which IRs are the most widely used physical mutagens, and their ionizing components within the electromagnetic spectrum include cosmic rays, gamma rays (γ) and X rays. Because of this, gamma rays are the most commonly used physical mutagens, which are emitted in the decay process of the cobalt (60Co) or cesium (137Cs) radioisotopes (Mba, 2013).
In the present study, the use of different doses of gamma radiation with 60Co showed that C. arabica seeds of the Geisha, Oro Azteca and Marsellesa varieties are very susceptible to the effects of this mutagenic agent at high doses. For the Geisha and Oro Azteca varieties, the effect was lethal starting at a dose of 200 Gy. The decrease in germination and the almost zero survival at high doses of radiation can be attributed to alterations at the cellular level (caused at the physiological or biochemical level), including damage to chromosomes.
The impact of IR on a plant depends on the species, variety, developmental stage and physiological state of the plant (Kim et al., 2011). When doses received by plants exceed the indicated limits, their growth and development are inhibited (Gudkov et al., 2019). IR can also damage DNA indirectly through the products of radiolysis, causing a cascade of reactive molecules. Reactive oxygen species (ROS) resulting from water radiolysis produce negative effects at high doses, as unusual concentrations of ROS are produced in cells, which causes the alteration of the cellular redox balance (Caplin & Willey, 2018). The production or overproduction of •OH and other ROS immediately triggers defense systems against oxidative stress by modulating the activities of detoxifying enzymes (e.g. peroxidases, ascorbate peroxidase, superoxide dismutase, and glutathione reductase) (Esnault, Legue, & Chenal, 2010). These radicals do not kill cells, but instead produce genetic abnormalities (Koyama et al., 1998).
Plant height, height to first leaf pair and stem diameter
In the factorial analysis of variance, statistical significance was detected in the variety and dose factors, where at least one level of each factor had a significant effect on the variables PH and SD (Table 1). PH in the Marseillaise variety was significantly affected (P ≤ 0.05) by gamma irradiation with the 100 Gy dose, although this variety was the only one that presented a response to the 400 Gy dose (Table 2). In the HFLP and LA variables, no significant differences were found between variety and dose with respect to the control.
Table 1 Mean squares of the analysis of variance of the morphological variables of the three varieties of Coffea arabica.
| SV | DF | HFLP (cm) | PH (cm) | SD (mm) | LA (cm2) |
|---|---|---|---|---|---|
| Variety | 2 | 0.05 | 6.66* | 0.00 | 3.10 |
| Dose (Gy) | 5 | 1.52* | 10.6** | 0.14* | 2.48 |
| Interaction | 10 | 0.40 | 0.73 | 0.02 | 1.83 |
| Error | 36 | 0.46 | 1.76 | 0.03 | 6.45 |
| Total | 53 | ||||
| CV (%) | 16.24 | 20.51 | 11.07 | 47.16 |
SV = source of variation; DF = degrees of freedom; HFLP = height to first leaf pair; PH = plant height; SD = stem diameter; LA = leaf area; CV = coefficient of variation. *, ** = significant at P ≤ 0.05 and P ≤ 0.01, respectively.
Table 2 Response of Coffea arabica seeds of the Geisha, Oro Azteca and Marsellesa varieties to different doses of gamma radiation (60Co).
| Varieties | Dose (Gy) | PH (cm) | HFLP (cm) | SD (mm) | LA (cm2) |
|---|---|---|---|---|---|
| Geisha | 0 | 7.09 ± 1.17 az | 4.27 ± 0.52 a | 1.58 ± 0.20 ab | 5.40 ± 3.04 a |
| 100 | 5.83 ± 1.04 ab | 4.26 ± 0.92 a | 1.66 ± 0.33 a | 4.99 ± 2.40 a | |
| 400 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Oro Azteca | 0 | 6.59 ± 1.38 a | 4.35 ± 0.70 a | 1.58 ± 0.10 ab | 5.88 ± 2.80 a |
| 100 | 5.93 ± 1.18 ab | 3.71 ± 0.50 a | 1.58 ± 0.15 ab | 4.99 ± 2.01 a | |
| 200 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Marsellesa | 0 | 6.56 ± 1.45 ab | 4.32 ± 0.73 a | 1.54 ± 0.16 ab | 5.21 ± 1.82 a |
| 100 | 5.32 ± 1.32 b | 3.97 ± 0.89 a | 1.66 ± 0.21 a | 5.30 ± 2.61 a | |
| 200 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 400 | 6.30 ± 1.27 ab | 4.2 ± 0.43 a | 1.38 ± 0.22 ab | 4.40 ± 1.30 a | |
| P-value | |||||
| Variety | 0.0259 | 0.9012 | 0.7571 | 0.6190 | |
| Dose | 0.0033 | 0.0437 | 0.0114 | 0.6810 | |
| Variety x dose | 0.6619 | 0.4300 | 0.3984 | 0.7530 |
HFLP = height to first leaf pair; PH = plant height; SD = stem diameter; LA = leaf area. Means of three replicates ± standard deviation. zMeans with the same letter within each column do not differ statistically (Tukey, P ≤ 0.05).
The results obtained coincide with those reported by Quintana et al. (2019), who observed a significant decrease in PH and SD in Típica variety coffee plants irradiated with 100 Gy. On the other hand, Dada et al. (2018) demonstrated that in C. arabica plants of the Mundo Novo, Kents and Geisha varieties, irradiated with 25 Gy, there was a significant reduction in PH, and in the Kents variety the effect was lethal. Marcu, Damian, Cosma, and Cristea (2013) report that in seeds of Zea mays L., with doses of 200 Gy, there was a decrease in shoot and root length. Likewise, in seeds of Glycine max L. Merrill, Alikamanoglu, Yaycili, and Sen (2011) observed that plant length decreased with doses of 200 Gy.
Leaf area and determination of LD50
The three varieties analyzed showed no significant differences in LA with any of the gamma radiation treatments (Table 2). In seeds of Phaseolus vulgaris L., Arena, de Micco, and de Maio (2014) report that LA decreases with high doses. In Típica variety coffee plants, Quintana et al. (2019) observed a significant reduction in leaf length and width starting at the 100 Gy dose. Dada et al. (2018) showed that in coffee plants of the Mundo Novo, Kents and Geisha varieties, irradiated with 25 Gy, there was a significant decrease in number of leaves.
In coffee, gamma irradiation has been used mainly to kill insects in stored beans (Ahmad, Tharappan, & Bongirwar, 2003). Therefore, in experiments with seeds, the LD50 should be estimated beforehand and an appropriate range of exposure to gamma irradiation should be established. The LD50 corresponds to the amount of absorbed radiation with which 50 % of the population that has been exposed survives; in this range, the appearance of useful mutations in genetic improvement programs is favored (Morela, González, & Castro, 2002).
Bolívar-González, Valdez-Melara, and Gatica-Arias (2018) conducted research related to the generation of genetic variability in coffee by chemical mutagenesis in embryogenic suspension cultures of C. arabica var. Catuaí, incubated at different doses of sodium azide (NaN3) and ethylmethanesulfonate (EMS). These authors showed that the LD50 for NaN3 was 5 mM for 15 min and for EMS it was 185.2 mM for 120 min. They also observed a significant reduction in cell viability as the concentration of NaN3 and EMS increased.
In the present study, the LD50 for survival of C. arabica var. Geisha plants was estimated at 70 Gy, for Marsellesa it was 85 Gy and for Oro Azteca it was 90 Gy. Due to the above, gamma irradiation doses between 70 and 100 Gy can induce genetic variation in C. arabica. Therefore, this study represents an advance in the development of protocols to generate and genetically improve arabica coffee varieties so that they are resistant to different types of abiotic stress, as well as tolerant to pests and diseases, especially coffee leaf rust caused by the pathogen Hemileia vastatrix.
Conclusions
Gamma irradiation with 60Co had a direct negative effect on the growth and development of C. arabica seedlings in the Geisha, Oro Azteca and Marseillaise varieties. Due to the high radiosensitivity of C. arabica to gamma irradiation with 60Co, for future research the dose range should be less than 100 Gy. Radiation with doses between 70 and 100 Gy can be used in C. arabica breeding programs to promote mutations that favor the obtaining of economically important variants.

