











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El camarón blanco (Litopenaeus vannamei, Bonne, 1931) es la especie que más se cultiva a nivel mundial, por lo que ha superado a la pro ducción pesquera. La industria del cultivo de camarón busca producir biomasa con máximos rendimientos al costo mínimo; en este aspecto, el uso de alimentos balanceados altamente digestibles puede mejorar la producción de camarón e incrementar las utilidades, además de re ducir considerablemente la contaminación ambiental. En este sentido, el precio del alimento es uno de los factores que más repercuten sobre los costos variables de producción en las granjas acuícolas, donde este insumo llega a representar entre 40 y 60% (Reyes, 1998). Por lo tanto, la calidad y el costo del alimento son factores críticos para la rentabi lidad de una granja camaronera. La determinación de la digestibilidad aparente de los nutrientes y alimentos en organismos acuáticos puede ser afectada por una estimación errónea de la cantidad de los nutrien tes defecados, en particular, cuando estos nutrientes se lixivian durante el tiempo que las heces permanecen en el agua antes de ser recolec tadas. Este efecto puede generar una subestimación de la cantidad de nutrientes en las heces y por lo tanto, una sobreestimación del valor de la digestibilidad aparente (Ricque et al., 2006).
Considerando lo anterior, existen muchos estudios sobre la digesti bilidad aparente de nutrientes en camarones. En algunos de ellos se considera la lixiviación de nutrientes en los pellets para ajustar los co eficientes de digestibilidad aparente (Ricque et al., 2006; Cruz-Suárez et al., 2009; Nieto-López et al., 2011, Villarreal-Cavazos et al., 2014). Sin embargo, en términos de lixiviación de nutrientes en las heces y con ajustes en los cálculos de digestibilidad aparente, sólo existen dos estudios: Fenucci et al. (1982) realizaron el primero, con camarón azul del Pacífico (Penaeus stylirostris Stimpson, 1874); mientras que el se gundo lo llevaron a cabo Smith y Tabrett (2004) con camarón tigre ne gro (Penaeus monodon Fabricius, 1798). En cuanto al camarón blanco del Pacífico, no existe información disponible en este tema, por lo que el objetivo del presente estudio fue evaluar dos tiempos de colecta de heces y determinar sus efectos sobre la pérdida de materia seca (PMS), proteína cruda (PPC) y aminoácidos (PAA) de las heces de juveniles de Litopenaeus vannamei .
Materiales y métodos
Elaboración de dieta experimenta. La dieta de referencia consistió en un alimento balanceado tipo comercial (Tabla 1), formulado y fabricado por la compañía Alimentos Costamar. El alimento fue molido y tamizado a 250 µm, para después ser mezclado con un marcador inerte (1% de óxido de cromo, Impex Continental lote 52-0305) y 1% de aglutinante (alginato de sodio, Aldrich-180947). La dieta se reconstituyó de acuer do con la metodología descrita por Villarreal-Cavazos et al. (2011). Los ingredientes se incorporaron en una mezcladora KitchenAid durante 10 min, y luego de agregar agua tibia (30%), durante 15 min se procesó la mezcla en un molino de carne (Torrey) con un dado con perforaciones de 1.6 mm de diámetro. El tiempo requerido para el procesado de la dieta fue de 40 min/kg-1, alcanzando temperaturas de 75-80ºC. Para secar los extrudidos se utilizó un horno ventilado a 100ºC durante 8 min y permanecieron a temperatura ambiente durante una noche para finalmente almacenarlos a 4ºC hasta su uso.
Tabla 1 Composición proximal y contenidos de aminoácidos (% base seca) de la dieta experimental para camarón blanco del Pacífico.

DR: dieta de referencia; AAE: aminoácidos esenciales; AAT: aminoácidos totales.
Animales y acuarios experimentales. En este estudio se utilizaron 15 juveniles de camarón blanco del Pacífico (L. vannamei) con una talla promedio de 4.9 ± 1.0 g por cada acuario.
Los organismos provinieron de la compañía mexicana Maricultura del Pacífico (Mazatlán, Sinaloa). Se utilizaron 4 acuarios experimentales de 120 l en un sistema de recirculación de agua marina sintética con capacidad de 9 toneladas métricas. Éste contó con un sistema de recir culación “air lift” (por acuario), un filtro biológico de contacto rotativo, un filtro de carbón activado, filtros de cartucho de 50 micras, un filtro de perlas BBF2 (Aquaculture Systems Technology LLC, New Orleans LA, EUA), un filtro de luz ultravioleta y tres fraccionadores de espuma.
Diseño experimental. La prueba se basó en dos tratamientos repre sentados por diferentes tiempos de permanencia de las heces en agua. Cada tratamiento experimental consistió de cuatro acuarios replicados (réplicas biológicas) distribuidos de la siguiente forma: se destinaron 2 para el tiempo 1 y los dos restantes para el tiempo 2. Al día siguiente los acuarios se alternaron de tratamiento; de tal manera que cada tiem po de colecta obtuvo 4 réplicas biológicas. En cada caso se utilizó un diseño completamente aleatorio y cada análisis químico se realizó por cuadruplicado (réplicas analíticas).
El tratamiento 1 (T1) consistió en realizar tres colectas de heces. La primera iniciaba una hora después de la alimentación, y las dos co lectas adicionales fueron realizadas con un intervalo de una hora entre ellas. La recolección de heces fue por sifoneo a una cubeta de 2 litros. Posteriormente, las heces se trasladaron por gravedad a canastillas de unicel (500 ml) donde fueron seleccionadas (pipetas plásticas de suc ción de 2 ml). Se eliminaron los restos de alimento, heces amarillas y cafés, colectando únicamente las heces verdes, que permanecían en su canastilla de unicel con un poco de agua marina en refrigeración a 4°C durante 3 horas.El mismo procedimiento se repitió para la segunda colecta co rrespondiente a la misma alimentación (se utilizó una canastilla de unicel por cada colecta). Al terminar con la tercera colecta, las heces fueron lavadas (dos veces) con agua destilada para eliminar el exceso de sales, ya que éstas pueden interferir con los análisis posteriores, y finalmente se concentraron en un frasco de plástico con tapa y se congelaron a -20°C hasta su procesamiento.
En el tratamiento 2 (T2) la recolección de las heces de la primera ración comenzó 15 minutos después de la primera alimentación y siguió con intervalos de 15 minutos. La segunda ración de heces se recolectaron con el mismo intervalo. La recolección de heces se realizó por succión directa del acuario utilizando una micropipeta de succión. Se seleccionaron sólo las heces de color verde, se lavaron inmediata mente en agua destilada para ser concentradas sin agua en un frasco de plástico con tapa, el cual permaneció en hielo durante las colectas y después fue congelado inmediatamente a una temperatura de -20ºC.
La dieta experimental fue ofrecida ad libitum. Se inició con el 10% de la biomasa de cada acuario, cantidad que fue dividida en dos racio nes por día (50% de la ración diaria cada una); la primera se ofreció a las 9:00 y la segunda a las 13:00 horas. Se realizaron 3 colectas por cada alimentación. La cantidad mínima de heces frescas recolectadas fue de 8 g por replicado y por tratamiento. Las heces recolectadas fue ron almacenadas en congelación a -20°C para después ser liofilizadas, molidas y almacenadas en congelación hasta su uso.
Análisis químicos. La composición bromatológica de la dieta experi mental y las heces fueron determinadas utilizando los siguientes mé todos: determinación de humedad, método 930.15; determinación de proteína cruda, método 990.03; ceniza, método 942.05; fibra, método 962.09B (A.O.A.C. 1997); lípidos, Soxhlet (Tecator, 1997), y el extracto libre de nitrógeno fue calculado por diferencia. La pérdida de nutrientes en agua marina también se estimó acorde a la técnica reportada por Villarreal-Cavazos et al. (2014). El contenido de cromo fue determinado mediante el método descrito por Bolin et al. (1952), modificado por Cruz-Suárez et al. (2009). Las determinaciones de AA se realizaron a través de los procedimientos descritos por Llames y Fontaine (1994) y Fontaine (2003).
Coeficientes de digestibilidad aparente. Los coeficientes de diges tibilidad aparente de materia seca, proteína y aminoácidos (CDA) de la dieta fueron calculados usando la ecuación reportada por Villarreal-Cavazos et al. (2014).
Análisis estadístico. Los valores se analizaron con una prueba t de Student para establecer posibles diferencias entre los tratamientos evaluados, con un nivel de significancia p <0.05. Las pruebas esta dísticas se realizaron mediante el programa computacional SAS (2008) versión 9.1.3.
RESULTADOS
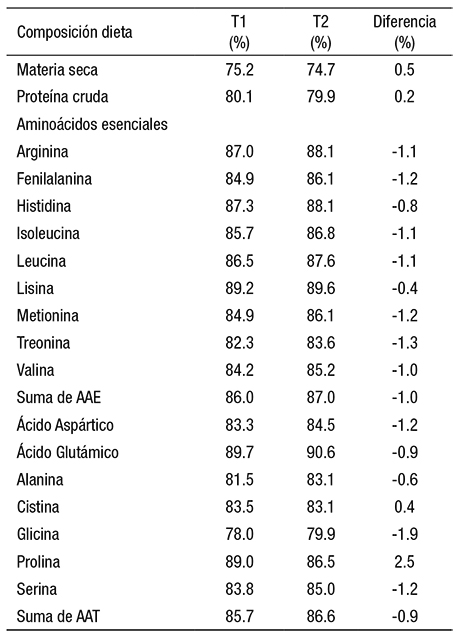
La composición química de la dieta experimental se presenta en la Tabla 1, donde se observa que el contenido de proteína cruda fue de 34.8%, el de lípidos, 9.6%, el de fibra cruda, 3.2% y el de cenizas, 11.8%; adicionalmente, se muestra el contenido de aminoácidos. Los resultados de proteína cruda en las heces oscilaron entre 28.9 (T1) y 29.0% (T2). Las concentraciones de óxido de cromo promedio fueron de 4.1% (T1) frente a 4.0% (T2). Adicionalmente, se determinó el contenido de aminoácidos de las heces recolectadas en diferentes tiempos, donde se encontró que las concentraciones de aminoácidos entre los tratamientos mostraron di ferencias inferiores al 10%, con excepción de arginina, metionina, alanina y ácido glutámico (11%), que fueron más altas para el T1, en general; sólo la prolina registró una menor concentración para el T1 (20%) que el T2. La suma de aminoácidos fue de 16.1% (T1) contra 14.8% (T2) (Tabla 2). Los CDA de materia seca (Tabla 3) fueron estadísticamente iguales, con 75.2% (T1) vs 74.7% (T2) y el mismo resultado se presentó en PC 80.1% (T1) frente a 79.9% (T2), es decir, no se presentaron diferencias estadísticas entre los tratamientos para estos parámetros. Los CDA de AA no presentaron diferencias significativas.
Tabla 2 Contenido de Materia seca, proteína cruda Cr2O3 y aminoácidos (%base seca) en heces de camarón blanco del Pacífico colectadas después de 3 horas de permanencia en agua (T1) versus colecta inmediata (T2).

AAE: aminoácidos esenciales; AAT: aminoácidos totales.
Discusión
Los contenidos de proteína cruda y óxido de cromo en heces para am bos tratamientos resultaron estadísticamente iguales. Esta información concuerda con el estudio realizado por Fenucci et al. (1982), quienes reportan pérdidas no significativas de proteína cruda (inferiores al 5%) y diferencias en la concentración de Cr2O3 (inferiores al 4%). La au sencia de diferencias significativas indica que no hubo una pérdida de nutrientes, ya que él Cr2O3 es insoluble e indigestible, lo anterior se ob serva en heces de juveniles de Penaeus stylirostris colectadas después de 15, 120 y 360 minutos de inmersión en agua marina.
Por otro lado, nuestros resultados de lixiviación de materia seca en heces coinciden con los resultados reportados por Smith y Tabrett (2004), quienes observaron que en el camarón tigre negro Penaeus monodon, las pérdidas de materia seca fueron del 3% en las heces después de 2, 3 o 6 h, sin ser significativas (pérdidas calculadas como la variación porcentual de la relación MS/ Cr2O3 con respecto a su valor de 1 h). Esto es muy parecido a los hallazgos de este estudio con una variación mínima del 1%. No obstante, los autores reportan pérdidas progresivas de 9, 15 y 31% para proteína cruda en heces, lo que puede estar asociado con la permeabilidad de la membrana peritrófica entre ambas especies de camarón (L. vannamei y Penaeus monodon). De acuerdo con Martin et al. (2006), la membrana peritrófica es un saco que envuelve las heces, que se encuentra constituido principalmente de quitina y es producido en el intestino medio. En el estudio de Wang et al. (2012) se ha documentado, en L. vannamei, el contenido de las proteínas de la membrana peritrófica relacionadas con la digestión, sistema inmune, antioxidantes y quitinasa; esta última, asociada con la porosidad o permeabilidad de la membrana peritrófica.
Por otra parte, los CDA de MS y PC no presentaron diferencias sig nificativas entre los dos tratamientos de este experimento. Los CDA de aminoácidos registraron diferencias cercanas al 1% entre los trata mientos, lo que demuestra que los contenidos de proteína cruda, ma teria seca y aminoácidos en las heces de Litopenaeus vannamei no se modifican por el efecto de la lixiviación de nutrientes en las heces en un lapso menor o igual a 3 horas.
De este modo, el uso alterno de los replicados 1 y 2, contra 3 y 4 réplicas para los tratamientos 1 y 2, asegura una adecuada identifica ción de los animales experimentales y elimina las fuentes de variación de los resultados. Así demostraron Alexandre et al. (2014) que la mem brana peritrófica sirve de soporte a estas enzimas. Por lo anterior, al impedir el consumo de heces con el tratamiento de colecta inmediata (camarones sometidos al T2), bien podría haberse presentado una dis minución en la eficiencia digestiva. Alternar la aplicación del tratamien to de colecta inmediata a los dos grupos replicados, permite repartir de manera equitativa este eventual efecto secundario sobre la eficiencia digestiva, y en última instancia, sobre la concentración de nutrientes en las heces. Los resultados del presente estudio indican que el tiempo de colecta de las heces generadas por él camarón blanco del Pacífico no tiene un efecto sobre la pérdida de nutrientes durante un periodo igual o menos a tres horas.

