











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El tomate (Solanum lycopersicum), cuyo centro de domesticación es México, en la última década se ha colocado como la cuarta hortaliza en producción en el mundo, y es la más consumida después de la papa (Solanum tuberosum L.) (Blancard et al., 2011; FAOSTAT, 2015); sin embargo, en los últimos años se han desarrollado nuevas enfermedades. Dentro de los patógenos que pueden causar pérdidas económicas se encuentran nemátodos, bacterias (Hernández-Romano et al., 2019), virus, viroides, pero son los hongos, por su gran diversidad de especies y hospedantes, los más importantes a nivel mundial (Agrios, 2005).
Uno de los hongos fitopatógenos es Colletotrichum coccodes, que ha sido reportado en algunas regiones del mundo por afectar principalmente a solanáceas y cucurbitáceas (Tsror y Johnson, 2000). En el caso de las solanáceas, en papa ocasiona el síntoma denominado “punto negro”, enfermedad que a principios de los 1990s era considerada de poca importancia; sin embargo, se reporta que ha causado grandes pérdidas en países como Israel y EUA (Barkdoll y Davis 1992; Tsror et al., 1994). C. coccodes causa el síntoma de antracnosis en tomate. Los síntomas iniciales aparecen en frutos maduros como manchas de color marrón claro, y a medida que la enfermedad avanza, las lesiones se agrandan y se observa un hundimiento. El hongo infecta a frutos a través de aberturas naturales; además, el mismo proceso se lleva a cabo a través de heridas ocasionadas por arena que transporta el viento (Piterková et al., 2009).
Cuando los frutos son verdes aún, el patógeno puede permanecer inactivo (infección quiescente) sobre el fruto hasta que éste alcanza la madurez fisiológica (Alkan et al., 2012). Dentro de las lesiones se pueden observar masas de conidios, aunque el hongo también puede atacar raíces, tallos y hojas. Los daños en frutos pueden afectar el pH en los tejidos, lo cual puede ser importante en términos económicos (Alkan et al., 2012; Dillard, 1992). Una de las características morfológicas importantes del hongo es la producción de esclerocios, los cuales pueden sobrevivir en residuos de cosecha o en el suelo, incluso por más de cinco años, y ser fuente de inóculo en campos donde no se realiza rotación de cultivos (Dillard y Cobb, 1998). La identificación precisa de un patógeno es esencial para reducir los efectos devastadores de la enfermedad mediante la aplicación de un manejo integrado del cultivo para disminuir los daños ocasionados por la enfermedad (Agrios, 2005; Blancard, 2011). Por lo anterior, el objetivo del presente trabajo fue determinar el agente causal de la antracnosis en frutos de tomate mediante técnicas morfológicas y moleculares.
MATERIALES Y MÉTODOS
Recolecta de materiales y aislamiento de simbiontes
En el año 2015, en el estado de Morelos, México, se observaron frutos de Solanum lycopersicum con incidencia del 7 % de antracnosis; de estos materiales se recolectaron muestras con síntomas como lesiones hundidas y en algunos casos con signos del patógeno. En 60 frutos recolectados se realizaron cortes de tejido epidermal (incluídos tejido sano y enfermo), éste fue lavado con agua corriente y posteriormente desinfestado con hipoclorito de sodio 0.5% durante 5 min, después se enjuagó dos veces con agua destilada estéril y se dejó secar sobre toallas de papel estériles a temperatura ambiente bajo condiciones asépticas en una cámara de flujo laminar. El tejido se sembró en medio de cultivo PDA (papa-dextrosa-agar) y agar-jugo V8, se incubó a 27 + 2 ºC bajo lámparas de luz blanca hasta el desarrollo de los hongos. La purificación de los cultivos se realizó mediante obtención de cultivos monoconidiales en medio de cultivo PDA de 7 a 10 d bajo las mismas condiciones de luz
Pruebas de patogenicidad
Se utilizaron 54 frutos sanos de tomate, los cuales se desinfestaron con solución de hipoclorito de sodio 0.5 % durante 5 min. Posteriormente, se sumergieron en agua destilada estéril durante 5 min, se colocaron sobre toallas de papel estéril y se dejaron secar a temperatura ambiente. De cada aislamiento se inocularon tres frutos con discos de 5 mm de diámetro que contenían al patógeno de 8 d de edad contados después de la siembra en medio de cultivo PDA. Se realizaron dos tipos de inoculación: 1) con herida (punción con aguja) y 2) sin herida. Los frutos se colocaron en charolas de unicel dentro de bolsa de plástico para proporcionar condiciones de humedad de 90% y temperatura ambiente de 25 ± 2 ºC; posteriormente, cada 48 h se midió el diámetro de la lesión. Finalmente, de los 60 frutos colectados se obtuvieron ocho aislamientos con clave 1A, 2A, 2B, 3, 4, 6A, 6B y 7. Los aislamientos se conservaron en glicerol al 30 % y se almacenaron en congelación para su posterior uso.
Caracterización morfológica
Se registraron variables como color de la colonia, tipo de micelio, crecimiento en la colonia en la caja Petri, morfología y tamaño de los conidios, apresorios, setas y presencia o ausencia de gútulas en los ocho aislamientos con tres repeticiones por cada uno. Se midió el diámetro de la colonia cada 48 h hasta que el hongo llenó la caja por completo. Para la medición de conidios se realizaron preparaciones permanentes en glicerol 50 %. Con ayuda del microscopio compuesto y del programa Motic Images Plus 2.0 (Group Co., Ltd) se midió el ancho y largo de 100 conidios de cada uno de los aislamientos. Para observar los apresorios se realizaron preparaciones temporales de cada aislamiento, se colocó una masa de micelio con conidios en agua destilada estéril sobre un portaobjetos, los cuales se mantuvieron en cámara húmeda durante 24 h a 27 + 2 ºC.
Caracterización molecular
Una vez obtenidos los aislamientos puros en cultivo monospórico, se inició el análisis molecular con la extracción de ADN genómico de los aislamientos fúngicos identificados morfológicamente como pertenecientes al género Colletotrichum, para lo cual se utilizó el kit de extracción PlantDNeasy® Minikit (Quiagen) de acuerdo con las instrucciones del fabricante. La reacción PCR se llevó a cabo con los oligonucleótidos ITS4 (5´-TCCTCCGCTTATTGATATGC-3´) e ITS5 (5´-GGAAGTAAAAGTCGTAACAAGG-3´) (White et al. 1990). Las condiciones consistieron en desnaturalización inicial de 5 min a 94 ºC, 40 ciclos de 30 s a 94 ºC, 30 s a 52 ºC, 30 s a 72 ºC y amplificación final por 7 min a 72 ºC (Damm et al., 2009). Los productos de la PCR se procesaron en el laboratorio de Macrogen Inc. (Corea del Sur) para su secuenciación
Análisis de la información
En el caso de la caracterización morfológica se aplicó análisis de varianza y para la comparación de medias se utilizó la prueba de Tukey (P ≤ 0.05) mediante el programa SAS versión 9.0 para Windows
Para la caracterización molecular los resultados obtenidos se compararon en la base de datos del Centro Nacional para la Información Biotecnológica (NCBI por sus siglas en inglés) con la herramienta BioEdit v7.0.9. (Hall, 1999). Las secuencias que presentaron similitud se utilizaron para la construcción de un árbol filogenético con el programa MEGA X (Kumar et al., 2018
RESULTADOS Y DISCUSIÓN
Pruebas de patogenicidad
Los primeros síntomas del hongo se observaron 3 d después de la inoculación (ddi), éstos fueron más visibles en frutos inoculados con herida artificial, mientras que en frutos sin herida sólo en algunos casos se observaron síntomas (Figura 1). A los 8 ddi todos los aislamientos fueron patogénicos al ocasionar lesiones de 0.6 a 3.5 cm de diámetro. En todos los casos el patógeno causó lesiones hundidas y presentó características particulares como masas negras (acérvulos), rajaduras o rompimiento de la epidermis entre los 4 y 6 d. En las pruebas de patogenicidad no hubo diferencia en cuanto a los días de aparición de síntomas (3 ddi) y diámetro de lesión (0.5 a 3.5 cm) a los 8 ddi. Estos resultados son comparables con lo reportado por Alkan et al. (2012), quienes encontraron que al inocular frutos de tomate con C. coccodes mediante herida artificial los primeros síntomas aparecen a los 4 ddi con lesiones de 0.4 a 3.5 cm de diámetro a los 7 ddi.
En cuanto a síntomas como lesiones circulares, hundidas y con aparición de signos del patógeno desde los 3 ddi, éstos fueron similares a lo reportado por otros autores (Alkan et al., 2012; Dillard y Cobb, 1998; Gleason y Edmunds, 2006; Li, 2013).
Estadísticamente no se encontraron diferencias significativas en patogenicidad entre aislamientos, pero sí las hubo en cuanto al método de inoculación (Cuadro 1); cuando la inoculación se realizó a través de una herida artificial todos los frutos inoculados presentaron lesión y ésta fue de mayor diámetro, a diferencia de la inoculación sin herida, donde se observó que sólo siete de 24 frutos presentaron síntomas. Aunque el patógeno puede infectar frutos en ausencia de heridas mediante la formación de apresorios, el ataque de insectos, las aberturas naturales y las heridas facilitan la infección del hongo (De Lapeyre de Bellaire, 2000; Gleason y Edmunds, 2006; Prusky, 1996; Rodríguez et al., 2009)

Características morfológicas
En cuanto al desarrollo de los hongos en medio de cultivo PDA y agar-jugo V8, se pudo observar que siete aislamientos desarrollaron micelio blanco aéreo, formación de esclerocios y anillos concéntricos (Figura 2). El aislamiento 2B sólo desarrolló micelio blanco y no presentó formación de esclerocios ni anillos concéntricos. Con respecto a la velocidad de crecimiento del patógeno en medio cultivo, se observó que éste fue más lento en el presente estudio en comparación con lo reportado por Cano et al. (2004), quienes observaron mayor diámetro de crecimiento por día en medio PDA y PCA (papa-zanahoria- agar), aunque sus aislamientos fueron incubados a 25 ºC durante 12 h de oscuridad alternados con 12 h de luz cercana a la ultravioleta, mientras que para esta investigación se utilizó medio PDA y agar-jugo V8 y se incubó a 27 + 2 ºC con lámparas de luz blanca.
Se observó la presencia de setas, conidios cilíndricos, hialinos, sin septos, en ocasiones con la presencia de crecimiento citoplasmático o gútulas con dimensiones de 3.16 a 4.16 µm de ancho y de 12.5 a 17.2 µm de largo. Se observaron apresorios ovalados, elípticos y lobulados (Figura 3). Estas características corresponden a la especie C. coccodes, de acuerdo con las claves de Sutton (1980). En cuanto a las características morfológicas de los conidios, éstas son similares a las reportadas por Cano et al. (2004). En otro estudio se reportaron características similares en medio de cultivo PDA, aunque el aislamiento correspondió al patógeno de pimiento (Capsicum annuum) (Stoyanova et al., 2013).
Identificación filogenética
Al comparar los resultados de los ocho aislamientos de Colletotrichum con la base de datos del GenBank se encontró 100 % de similitud con Colletotrichum coccodes, por lo que se tomó un aislamiento representativo y se realizó una secuencia consenso que fue depositada en el GenBank con número de accesión (KX632089) y se comparó con nueve secuencias del Banco de Genes del CNBI (Figura 4). Anteriormente la identificación de los hongos se realizaba mediante claves taxonómicas; sin embargo, en la actualidad se utiliza la reconstrucción filogenética para la identificación a nivel de especie (Guerber et al., 2003).
Cuadro 1 Diámetro de lesiones Colletotrichum coccodes ocho dias después de la inoculación en frutos de tomate (Solanum lycopersicum)
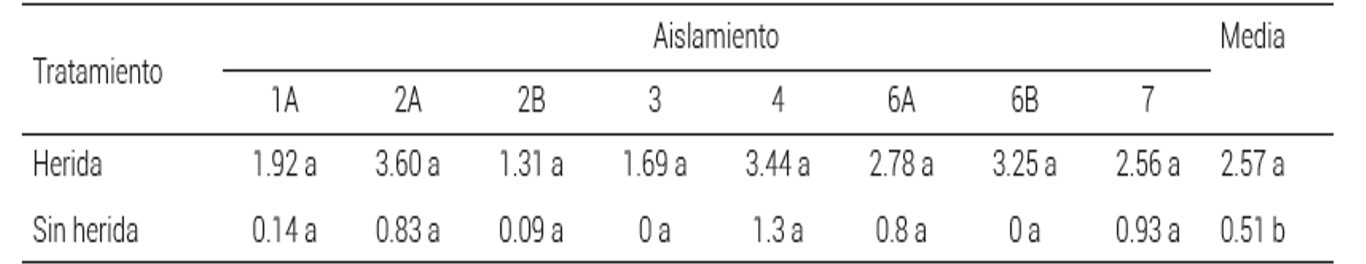
Valores con la misma letra dentro de filas para los aislamientos, son estadísticamente iguales (Tukey, P ≤ 0.05). En la columna de la extrema derecha,valores con diferente letra son estadísticamente diferentes (Tukey P ≤ 0.05).
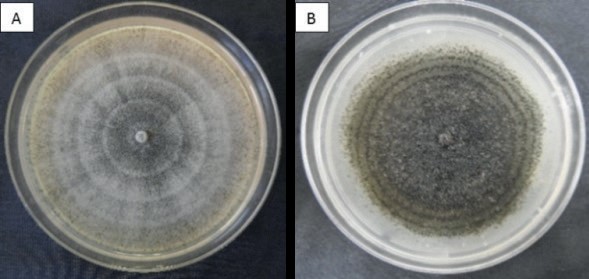
Figura 2. Colonias de Colletotrichum coccodes Crecidas en medios de cultivo: A) agar-jugo V8 y B) papa dextrosa-agar a los 12 dias de edad

Figura 3 A) Acérvulo con setas; B) Conidios hialinos cilíndricos, sin septos gutulados de Colletotrichum coccodes; C) Formación de apresorios melanizados de color café oscuro, de forma globosa a irregular de 24 h. Magnificación 40x

CONCLUSIONES
Con base en la morfología y reconstrucción filogenética se considera a C. coccodes como el causante del síntoma de antracnosis en frutos de tomate. En las pruebas de patogenicidad se observaron lesiones hundidas y signos del patógeno con manifestaciones en forma de masas negras (acérvulos), rajaduras o rompimiento de la epidermis. La aparición de síntomas y tamaño de lesión fue mayor en frutos inoculados mediante una herida. Este es el primer reporte de C. coccodes que afectó a frutos de tomate en México.

