










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de fitopatología
versão On-line ISSN 2007-8080versão impressa ISSN 0185-3309
Rev. mex. fitopatol vol.39 no.2 Texcoco Mai. 2021 Epub 03-Nov-2021
https://doi.org/10.18781/r.mex.fit.2101-5
Scientific articles
Antifungal activity of Trichoderma harzianum and T. koningiopsis against Fusarium solani in seed germination and vigor of Miahuateco chili seedlings
1 Centro de Agroecología, Benemérita Universidad Autónoma de Puebla (BUAP); Edificio VAL 1, Km 1,7 carretera a San Baltazar Tetela, San Pedro Zacachimalpa, C.P. 72960, Puebla, México
2 Centro de Investigaciones en Ciencias Microbiológicas. Instituto de Ciencias (ICUAP), Benemérita Universidad Autónoma de Puebla (BUAP); Edificio VAL 1, Km 1,7 carretera a San Baltazar Tetela, San Pedro Zacachimalpa, C.P. 72960, Puebla, México;
3 Laboratorio de Patología Vegetal. Instituto de Investigaciones Agropecuarias y Forestales, Universidad Michoacana de San Nicolás de Hidalgo (UMSNH). Carretera Morelia-Zinapécuaro Km 9.5, Tarímbaro, Michoacán, México. C.P. 58880.
Chili is the second largest vegetable in Mexico. The objective of the investigation was to evaluate the in vitro and in vivo antagonistic activity of Trichoderma harzianum (T-H4) and T. koningiopsis (T-K11) versus Fusarium solani (MX-MIC 798) in the germination and establishment of seedling of chili Miahuateco. The dual culture technique was used to determine the percentage of radial growth inhibition (PRGI) of the MX-MIC 798 strain. In addition, the germination percentage in Miahuateco chili seeds was analyzed in nursery, as well as the mortality of seedlings and degree of severity at 40 days after transplantation (dat) in Santa María Tecomavaca, Oaxaca, through biocontrol and chemical control (Mancozeb 80®). The T-H4 strain presented the highest PIRG antagonic level (53.3%) in vitro and class II on Bell’s scale, also obtained 82% germination in Miahuateco chili seeds in nursery and 48% of field mortality; So, he matched chemical control and beat T. koningiopsis T-K11. The antifungal activity of Trichoderma spp., They offer an alternative for the biocontrol of the wilt and necrosis as a root in the cultivation of Miahuateco chili caused by F. solani MX-MIC 798.
Key words: Antagonism; mycelial growth; PRGI; mortality; Capsicum
El chile es la segunda hortaliza de mayor producción en México. El objetivo de la investigación fue evaluar la actividad antagónica in vitro e in vivo de Trichoderma harzianum (T-H4) y T. koningiopsis (T-K11) versus Fusarium solani (MX-MIC 798) en la germinación y establecimiento de plántula de chile Miahuateco. Se utilizó la técnica de cultivo dual para determinar el porcentaje de inhibición de crecimiento radial (PICR) de la cepa MX-MIC 798. Además, se analizó el porcentaje de germinación en semillas de chile Miahuateco en vivero, así como la mortalidad de plántulas y grado de severidad a los 40 días después del trasplante (ddt) en Santa María Tecomavaca, Oaxaca, a través de biocontrol y control químico (Mancozeb 80®). La cepa T-H4 presentó el nivel antagónico PICR más alto (53.3%) in vitro y clase II en la escala de Bell, asimismo obtuvo 82% de germinación en semillas de chile Miahuateco en vivero y 48% de mortalidad en campo; de manera que igualó al control químico y superó a T. koningiopsis T-K11. La actividad antifúngica de Trichoderma spp., ofrecen una alternativa para el biocontrol de la marchitez y necrosis en raíz del cultivo de chile Miahuateco causada por F. solani MX-MIC 798.
Palabras clave: Antagonismo; crecimiento micelial; PICR; mortalidad; Capsicum
The genetic diversity of the different types of chilies in Mexico has been scarcely studied, and a large variety have been found to be native, such as the Poblanos (Toledo-Aguilar et al., 2016). In the year 2018, the Poblano chili (Capsicum annuum) was found to have been planted in an area of approximately 17,729.80 ha, with a production of 391,133.26 t and an income of $ 3’038,997.37 pesos (SIAP, 2020). The Poblano chili has great gastronomic, economic and social importance, since it is a basic ingredient of traditional dishes and has a culinary influence on other foods worldwide, being one of the cornerstones of development or providers of jobs and income in the areas (Rodríguez et al., 2007; Pérez-Carrasco et al., 2017).
In the town of Santa María Tecomavaca, in the state of Oaxaca, Mexico, the Miahuateco chili is grown, which is a variety of Poblano chili and the current production in the region is jeopardized by the presence of fungal diseases. The genus Capsicum presents a higher fungal incidence in the seedling stage. These diseases are caused by pathogens such as Phytophthora sp., Rhizoctonia sp. and Fusarium spp., which lead to the loss of over one third of the production of the crop. The main symptom is the premature death of the plants, since it causes the obstruction and imbalance in the functioning of the vascular bundles, defoliation, color changes, curling of the foliage, damage to reproductive structures, early maturation and irregular rot of the root and necrosis in the stem (Rivera-Jiménez et al., 2018; Guenoun et al., 2019). Fusarium oxysporum and F. solani have been isolated from the rot of the crown and the base of the stem of mature Poblano chili plants in nursery. In addition, F. solani is the causal agent of wilting and necrosis in the root and stem of the Chilaca chili in Queréndaro, Michoacán, Mexico (Rivera-Jiménez et al., 2018; Reyes-Tena et al., 2019).
The products of chemical synthesis have been used for decades to control these diseases (Gan and Wickings, 2017). However, their use is related to the generation of resistance in phytopathogens, damages to the environment and to human health (Chen et al., 2014; Andrade-Hoyos et al., 2019). In the light of this situation, biological control is considered an efficient practice and environmentally feasible for the development of sustainable agriculture (Pérez-Torres et al., 2018).
The genus Trichoderma contains the most relevant antagonistic species for the biocontrol of phytopathogenic fungi, used mainly by organic farmers and in good agricultural practices for the management of diseases and as a compost enhancer (Romero-Arenas et al., 2017; Launio et al., 2020). Its use and success in agriculture is due to action mechanisms such as competition for space, mycoparasitism, antibiosis (Nawrocka et al., 2018) and the production of volatile compounds (Hernández-Melchor et al., 2019). In previous investigations, T. harzianum T-H4 presented a higher percentage of inhibition (78.3%) against Phytophthora cinnamomi than T. koningiopsis T-K11 in vitro (Andrade-Hoyos et al., 2020). Likewise, the strain T-H4 presented 63.6% of PICR and class III of the Bell scale against Fusarium sp. (Morales et al., 2020). Consequently, the aim of this investigation was to evaluate the antagonistic activity of two strains of Trichoderma (T-H4 and T-K11), versus F. solani MX-MIC 798 in the germination of seeds and vigor of Miahuateco chili seedlings for the town of Santa María Tecomavaca, Oaxaca, Mexico.
Materials and methods
Antagonism of T. harzianum and T. koningiopsis against F. solani in dual cultures. The evaluation of the antagonism was carried out with strains T-H4 of T. harzianum and T-K11 of T. koningiopsis, both isolated from the root of Persea americana, the sequences of which are found in the National Center for Biological Information (NCBI) data base, with accession numbers MK779064.1 and MK791648.1, respectively. Likewise, F. solani strain MX-MIC 798, with accession number MK532475 was used, which was donated by the Plant Pathology Laboratory of the Agricultural and Forestry Research Institute of the Universidad Michoacana. All the biological material is stored in laboratory 204 of the Eco-Campus Valsequillo of the Institute of Science of the Benemérita Universidad Autónoma de Puebla (BUAP).
For the evaluation of mycelial growth, fragments of F. solani (MX-MIC 798), T. harzianum (T-H4) and T. koningiopsis (T-K11), 5 mm in diameter, were planted in Petri dishes with PDA (Potato Dextrose Agar), each in triplicate and incubated in the dark at 28 °C for 10 days. The mycelial diameter was measured every 12 h to estimate the growth speed (cm), which was calculated with the function of linear growth y=mx + b (where ‘y’ is distance, ‘x’ is time and ‘b’ is the constant factor) and it was expressed in centimeters per day (cm d-1) (Zeravakis et al., 2001). The diameter of the mycelial growth was determined with a digital caliper (CD-6 Mitutoyo) taking the same direction in triplicate.
The dual culture technique was used following Andrade-Hoyos et al. (2019) in triplicate to determine the percentage of inhibition of radial growth (PICR) for each trial evaluated, for 10 days using the formula PICR= [(R1-R2/R1) x 100], where R1 is the diameter of the control (average of the radial growth of the triplicates in each strain) and R2 is the diameter of the tested organism. To complement the evidence of the antagonism, each trial was compared and classified using the scale established by Bell et al. (1982) (Table 1).
Nursery tests to analyze the percentage of germination and rotting of seedlings. Miahuateco chili seeds were used, which were provided by farmers from Santa María Tecomavaca in the state of Oaxaca. The trial was held under controlled conditions in the nursery of the Eco-Campus Valsequillo, ICUAP-BUAP. Before the seeds germinated, they were disinfested with sodium hypochlorite at 0.3% (v/v) for 10 min, rinsed three times with sterile water and dried with sterile paper (Illa et al., 2019). The pathogen F. solani was inoculated in a salt suspension at a concentration of 1x108 conidia mL-1 with a Neubauer chamber (PAUL MARIENFELD), in 400 seeds for each treatment under study; once the pathogen was soaked and the seeds were dry, T. harzianum and T. koningiopsis were inoculated. This was carried out with a suspension of teliospores at a concentration of 1x108 conidia mL-1 for two separate treatments, each one with 400 seeds (Reyes-Tena et al., 2019).
Table 1 Scale of antagonisms proposed by Bell et al. (1982).
| Clase | Características |
|---|---|
| I | Crecimiento de Trichoderma spp., que cubrió toda la superficie del medio y redujo el crecimiento micelial del patógeno. |
| II | Crecimiento de Trichoderma spp., que cubrió al menos 2/3 partes del medio. |
| III | Trichoderma spp., y fitopatógeno crecieron 1/2 y 1/2 la superficie del medio, uno no se sobrepuso al otro. |
| IV | Hongo patógeno creció al menos 2/3 partes del medio y resistió a la invasión de Trichoderma spp. |
| V | Crecimiento del patógeno que cubrió toda la superficie del medio. |
Once the seeds were inoculated with the fungi, the seeds were placed in germinating trays with Peatmoss and Agrolita (1:1 v/v) after sterilizing it at 121 °C and 15 lb of pressure (Mannai et al., 2018). The seeds were planted using a standard mechanical procedure; the humid substrate was placed to take up three quarters of the total cavity, and one seed was placed in each cavity at a depth of 1 cm (Castillo-Aguilar et al., 2015). Mancozeb 80® was used as a chemical treatment, following the manufacturer’s recommendations for the production of the seedling (1 kg of seed for every 5 g of product), along with the use of a control treatment, which consisted of seeds that were disinfested in a solution of sterile distilled water and sucrose at 3%, without fungal activity. Finally, the trays were wrapped with black plastic for 10 days and the temperature was kept between 10 and 27 °C during germination (Castillo-Aguilar et al., 2015). The percentages of seed germination were calculated using the following formula: seed germination (%) = (number of germinated seeds/total number of seeds) × 100 (Al-Fadhal et al., 2019). The data were taken when the control treatment presented a germination rate of ≥ 90%.
Mortality and rate of severity in plants 40 dat. We used 1,500 Miahuateco chili seedlings 15 days after emergence, provided by local farmers. The trial was conducted under open field conditions in the town of Santa María Tecomavaca, between parallels 17° 51’ and 18° 03’ latitude N, and meridians 96° 57’ and 97° 15’ longitude W, at an altitude between 400 and 2600 masl. The predominant climates in the area are dry warm (40.5%) and semidry warm (17.5%) (INEGI, 2017). The ground was prepared by local farmers three months before planting. The field was plowed to a depth of 50 cm in order to reduce compaction and favor draining, along with two harrows-turns to a depth of 30 cm to ventilate the soil. The transplanting date was March 27, 2018, at a depth of de 3 cm. Fertilization was carried out with 90-80-80 of N, P and K after 15 days (dat). The experimental design was random block with three repetitions and the population density was six plants per m2. During the development of the crop, the field was irrigated every five to six days, before the soil humidity reached 50% of the capacity of the field (Vázquez-Casarrubias et al., 2011).
F. solani (MX-MIC 798) was inoculated at the neck of each chili plant (100 seedlings per treatment distributed into five random blocks) with 1 mL of solution at a concentration of 1x108 conidia. After 36 h, the plants were inoculated with strains of Trichoderma (T-H4 and T-K11) at the same concentration as the pathogen (1x108 conidia mL-1), for each strain, applied to the root for all treatments. For the chemical treatment, Mancozeb 80® was applied on the third day, following the recommendations provided by the manufacturer (480 g 200 L-1 of water per ha). The control treatment was only given sterile water with no fungal activity. The plants were kept on the field for 40 days. The severity of the disease was calculated in the infected section in relation with the total root length (Perveen et al., 2010) and classified on the scale of 0 to 3, where 0 = 0 to 25% of severity, 1 = 26 to 50%, 2 = 51 to 75% and 3 = 76 to 100% (Bokhari and Perveen, 2013).
Finally, after week 6 in the field, reisolation was carried out. Small symptomatic root sections were taken (approx. 1 × 1 cm), sterilized with sodium hypochlorite at 1% for 30 s and rinsed three times with sterile distilled water. The segments were then placed in a PDA medium and incubated in the dark at 28 °C for 10 days. The cultures that developed were purified with monosporic or hyphae tips and the structures were identified using taxonomic keys by Barnett and Hunter (1998) in a microculture system using an optical microscope (Carl Zeiss, Jena, Germany) at a magnification of 1000x (Samson et al., 2014), and were stored in a 20% glycerol solution at -84 °C (Morales et al., 2020).
Statistical analysis. The data were analyzed with an ANOVA (two-ways) in the IBM SPSS Statistics statistical package, version 25. Growth speed and development rate were the response variables with three repetitions and they were validated in duplicate in a totally random statistical design. Means were compared with the method by Tukey-Kramer with a level of probability of p≤0.05. PICR, the percentage of germination, mortality (%) and the severity of the disease were expressed as percentages and transformed with an angular arccosine √x+1. They then underwent an analysis of variance (two-way), along with the variables of total height, root height and rootless plant; the Tukey-Kramer test was used to determine the significant differences between treatments with a significance of p≤0.05. In turn, the PICR was compared based on Student’s t test for paired samples (p≤0.05) using the same statistical package.
Results and discussion
Evaluation of the antagonism of T. harzianum and T. koningiopsis against F. solani in dual cultures. Areas of interaction appeared between T. harzianum (TH-4) and T. koningiopsis (T-K11) against F. solani (MX-MIC 798), where parasitism was observed after 132 h. The rate of development and speed of growth presented significant differences (p≤0.05), where T. harzianum obtained the highest value (Table 2) with 0.972±0.008 mm h-1 and 1.3238±0.0024 cm d-1, respectively. Morales et al. (2020) reported a higher speed of growth than in the present study for strain TH-4 with 1.86± 0.22 cm d-1 and 1.67 ± 0.01 mm h-1 for the development rate. F. solani presented the lowest speed of growth (0.4718±0.00063 cm d-1). A study carried out by Andrade-Hoyos et al. (2019) reaffirm the results obtained in the present investigation, in which different species of Trichoderma spp. have superiorities over the mycelial growth of F. oxysporum (0.22 mm). The reduction in the growth rate of dual cultures is an indicator of the antagonistic ability of Trichoderma (Guigón-López et al., 2010). The percentage of inhibition of radial growth (PICR) of F. solani exerted by T. koningiopsis and T. harzianum presents significant differences (p<0.0001) from day eight (Table 2), when the strain T-H4 was 1.33% higher than T. koningiopsis T-K11 and a class II classification (Figure 1 A and B), according to the scale established by Bell et al. (1982).
The present investigation coincides with Sundaramoorthy and Balabaskar (2013), who found an inhibition of 53% in the mycelial growth of F. oxysporum f. sp. lycopersici. Likewise, Jagraj et al. (2018) reported a PICR of 55.6% for F. oxysporum against T. koningiopsis, for strains isolated from tomato plants. Michel-Aceves et al. (2019) obtained a class II antagonism on F. oxysporum with T. harzianum, which were similar results to those obtained in this investigation.
Table 2 Rate of development, growth speed, percentage of inhibition of radial growth of Trichoderma and F. solani under in vitro conditions.
| Nombre | Tasa de desarrollo (mm h-1)y | Velocidad de crecimiento (cm d-1)y | PICRz | Clase Antagonismo |
|---|---|---|---|---|
| T. harzianum | 0.972±0.08a | 1.3238±0.024a | ||
| T. koningiopsis | 0.912±0.012b | 1.2369±0.063b | ||
| F. solani | 0.391±0.019c | 0.4718±0.006c | ||
| F. solani vs. T. harzianum | 53.33±0.20a | II | ||
| F. solani vs. T. koningiopsis | 52±0.002b | III | ||
yDifferent letters mean significant difference between treatments according to Tukey-Kramer for p≤0.05.
zMeans with different letters indicate significant differences with Student’s t test (p≤0.05).
Suárez et al. (2008) presented 12 isolations of T. harzianum confronted with F. solani, where they obtained a PICR between 60 and 70% of the antagonism. Nagendran et al. (2016) reported that 15 isolations of Trichoderma spp. reached an inhibition of over 50% on the mycelial growth of F. solani, in which strain Phyto-4 obtained the highest inhibition (66.1%). Likewise, Sánchez-García et al. (2017) reported a PICR of 59.1% for F. solani against T. harzianum (Tri-4), which are higher results than those obtained in the present investigation. This indicates that T. harzianum may have a higher rate of incorporation of nutrients and a higher metabolism than F. solani, as well as different mechanisms for the secretion of hydrolytic enzymes, including cellulases, chitinases, glucanases, xylases and proteases, which may be involved in biocontrol mechanisms, helping it make better use of nutrients from the medium and deprive the pathogen of the resources (Michel-Aceves et al., 2005). T. harzianum is also capable of inhibiting the growth of phytopathogens, since it produces numerous antibiotics, such as trichodermine, suzukacillin, alameticin, dermadin, penicillin, trichothecenes, trichorzianines and others (Andrade-Hoyos et al., 2020).
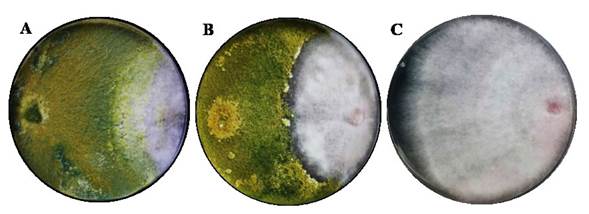
Figure 1 Antagonism of Trichoderma with F. solani on the scale byBell et al. (1982) after 132 h in dishes with PDA medium, incubated at 28 °C for 10 days, (A) class II antagonism of T. harzianum (T-H4) and F. solani (MX-MIC 798), (B) class III antagonism of T. koningiopsis (T-K11) and F. solani (C) F. solani without interaction.
Mycoparasitism is another action mechanism displayed by the genus Trichoderma. In this investigation, we found granulation from the strain MX-MIC 798 induced by T. harzianum, as well as the breaking of septa and strangulation (Figure 2). This highlights the eminent mycoparasitic ability of strain TH-4. Duarte-Leal et al. (2017) found mycoparasitism between hyphae in T. asperellum (Ta.25), in which they observed lysis in hyphae of F. oxysporum f. sp. ciceri (F-50), as reported in the present investigation. Other studies, carried out by Hyder et al. (2017), refers to the enzyme degradation of the cell wall of phytopathogenic fungi during the mycoparasitic action, which causes lysis and thus the disorganization of the cytoplasmic content. For T. koningiopsis, this investigation was unable to observe the mycoparasitic mechanism or breaking of the septae.
The growth rate and antagonism tests are tools to predict and determine the biocontrolling ability. According to Morales et al. (2020), these studies help select efficient antagonistic strains for evaluation under field conditions and obtain the best results.
Percentage of germination and rot caused by F. solani in Miahuateco chili seedlings in the nursery. The germination of the seeds from the control treatment was displayed after 12 days with 91% and significant differences (p=0.0023) in regard to the remaining treatments (Table 3), in which the effect of the inoculation from F. solani (MX-MIC 798), which affected the neck of the root, causing rot and death of the plant, as well as the lowest percentage of germination (30%). In this investigation, the Miahuateco chili seeds inoculated with strain MX-MIC 798 treated with T. harzianum T-H4 showed a germination rate of 82%, even higher than the value reported by Ezziyyani et al. (2004), who obtained a percentage of germination of 60% in bell pepper seeds treated with T. harzianum after 10 days. In untreated plants inoculated with F. solani, root rot reached level 4.0 and a mortality of 70% (Table 3) in the scale proposed by Ezziyyani et al. (2004). Madhavi et al. (2006) reported a percentage of germination of 90.3% in chili seedlings (C. annuum) inoculated with T. harzianum-m1 and F. solani under nursery conditions, similar results to thos found in the present investigation.
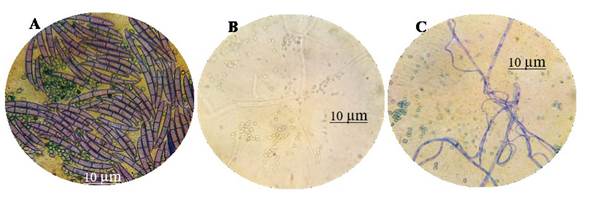
Figure 2 Mycoparasitism of T. harzianum (T-H4) on F. solani (MX-MIC 798) in PDA medium, incubated at 28 °C for 10 days; A) interaction of T. harzianum conidia vs. F. solani septated macroconidia; B) granulation and breaking of septa of the strain MX-MIC 798 induced by T. harzianum; C) strangling of F. solani hyphae by strain TH-4.
The microbial antagonists reduced rot caused by F. solani under nursery conditions in Miahuateco chili seeds significantly (22%), whereas T. harzianum presented the lowest percentage of plant mortality (18%), higher than strain T-K11, which obtained 26%. The group treated with Mancozeb 80® reduced seed mortality by 28% and presented no significant differences with T. koningiopsis; the control treatment accounted for a feasibility of 91% in seed germination (Table 3).
The Mancozeb 80® chemical treatment presented high mortality results, which could indicate the creation of resistance by F. solani to this fungicide. Some fungicides used, such as triazoles, tend to develop resistance to the fungal populations due to genic factors (accumulative with time), since they only have one place of action (Leroux and Walker, 2011; Ziogas and Malandrakis, 2015). Chen et al. (2014) reported that Fusarium fujikuroiuses a resistance mechanism generated by the mutation of β2tub by the fungicide carbendazim, making its management difficult, although it opens the possibility of generating new control measures with antagonistic agents.
Mortality and rate of severity in plants 40 dat. The Miahuateco variety chili seedlings that were inoculated with F. solani at the moment of transplanting displayed rot in the roots 40 days after inoculation. Long reddish lesions were found, which covered 10% of the total surface of the main root and reddish-brown spots were also observed in the section of the stem found under the soil surface, along with a considerable loss of lateral roots. This coincided with descriptions by Agrios (2006).
Table 3 Effect of the antagonistic activity of T. harzianum and T. koningiopsis on the germination and rot caused by F. solani in Miahuateco chili seeds, under nursery conditions.
| Tratamientos | Germinación(%)y | Severidadz | Podredumbre causada por F. solani (%) | |
|---|---|---|---|---|
| F. solani (MX-MIC 798) | 30d | 4 | 80 | |
| F. solani vs. T. harzianum (T-H4) | 82b | 1 | 25 | |
| F. solani vs. T. koningiopsis (T-K11) | 74c | 2 | 30 | |
| F. solani vs. Mancozeb 80® | 72c | 2 | 30 | |
| Tratamiento control | 91a | 0 | 0 | |
zEvaluation performed on the gradual scale of 0 (no symptoms) to 5 (dead plant) proposed byEzziyyani et al. (2004).
yDifferent letters mean significant differences between treatments according to Tukey-Kramer for p≤0.05.
The field results showed that the treatments with antifungal activity were effective to reduce mortality by wilting and root rot in Miahuateco chili plants under induced infection by 48 and 51% respectively, and it reduced the severity of the disease 40 dat by 50% (Table 4). Strain T-H4 presented the lowest percentage of plant mortality (48%), yet it displayed no significant differences with the Mancozeb 80® fungicide, which accounted for 47% of the mortality. Similarly, Thangavelu et al. (2004) showed that T. harzianum (Th-10) efficiently controlled the fusarial wilting of the banana tree, comparable to chemical control. Nagendran (2016), found the highest percentage (36.9%) of the reduction of the disease with strain Phyto-6, and also displayed a lower severity in witling, as shown by the present investigation. T. koningiopsis T-K11 obtained a mortality of 51%, presenting significant differences with the control group (23%), which obtained the lowest mortality.
During their development, the plants infected by F. solani presented symptoms of yellowing in the basal part, yellowing of the nervations in 30% of leaves, as described by Black et al. (1993) in chili plants (C. annuum) that survived the infection on the field. The analysis of variance for the total plant height displayed highly significant differences (p= 0.0002) between treatments. Mancozeb 80® obtained the greatest height (25.6±2.78 cm), followed by plants inoculated with T. harzianum (24±1.98 cm) and T. koningiopsis (21±1.81 cm). The lowest height appeared recurrently in plants inoculated with F. solani with 16±0.28 cm, as shown in Figure 3A. Agrios (2006) mentions that plants affected by Fusarium sp. reduce their growth, due to the ability of the pathogen to colonize roots, preventing the adequate nutrition of the plant.
The genus Trichoderma can parasite pathogenic fungi and produce antibiotics. It also presents positive effects in plant growth, yield, absorption of nutrients, fertilizer use efficiency and the systemic resistance of plants to diseases (Hyder et al., 2017; Andrade-Hoyos et al., 2020). In a study carried out by Cole and Zvenyika (1988), they noticed that the infection caused by F. solani in tobacco plants could be controlled when applying T. harzianum in disease management procedures. Likewise in a study by Yedidia et al. (1999), when inoculating T. harzianum in early growth, where the absorption of nutrients in the plant improved. These investigations agree with studies by Tsegaye-Redda et al. (2018), who mention that Trichoderma sp. has the ability to antagonize the phytopathogens present in the soil, and also induces the promotion of plant growth and protects plants from biotic and abiotic stress. The studies by Dubey et al. (2007) have been confirmed in T. harzianum, where seed germination, root health and sprout length have improved and the frequency of healthy plants has increased, as confirmed by the present investigation.
Forty dat with the strain MX-MIC 798 that led to the development of necrotic lesions in the roots and stems of evaluated plants, we proceeded to re-isolate in order to satisfy Koch’s postulates. The fungal cultures developed abundant aerial mycelia, with a cotton-like texture and white in color with purple tones (Figure 3B), which appeared more intense in the middle of the culture medium (Rentería-Martínez et al., 2018). The duration of the microculture until chlamidospores were visible was 14 days; for macroconidia, seven days; and for phialides and microconidia, four days. Oval-shaped septated mycelia and abundant microconidia were observed, with an average size of 2.4 x 6.3 μm in length by 2 x 4 μm in width (Figure 3C), along with the typically crescent-moon shaped macroconidia, one to four septa (Figure 3F), an average size of 2.4 x 4.1 μm in width and 11.8 x 36.4 μm long (Barnett and Hunter, 1998). Likewise, chlamidospores were also observed (Figure 3E), 8.4-10.5 μm in diameter and with particular morphometric characteristics reported by Montealegre et al. (2003).
Table 4 Antagonistic activity of Trichoderma on the percentage of mortality 40 dat and severity of the disease induced by F. solani in Miahuateco chili crop.
| Tratamientos | Mortalidad(%)z | Severidad de la enfermedad | Tamaño de planta sin raíz (cm)z | Tamaño de raíz (cm)z | Altura total (cm)z |
|---|---|---|---|---|---|
| F. solani (MX-MIC 798) | 86±1.34a | 3 - 76% | 12±1.12c | 1.3±0.12 | 16±0.28c |
| F. solani vs. T. harzianum (T-H4) | 48±1.16c | 1 - 25% | 18±1.97a | 2.1±0.09b | 24±1.98a |
| F. solani vs. T. koningiopsis (T-K11) | 51±1.31b | 2 - 51% | 17±2.62ab | 1.9±0.30b | 21±1.81a |
| F. solani vs. Tratamiento químico (Mancozeb 80®) | 47±1.36c | 1 - 25% | 21±2.15a | 2.6±0.26a | 25.6±2.78a |
| Grupo control | 23±0.38d | 0 - 0% | 16±0.02b | 1.6±0.18b | 18.4±1.08b |
Evaluation performed on the gradual scale of 0 to 3, where 0 = 0 to 25% of severity, 1 = 26 to 50%, 2 = 51 to 75% and 3 = 76 to 100% proposed by Bokhari and Perveen, 2013.
zDifferent letters mean a significnat difference between treatments according to Tukey-Kramer for p≤0.05.
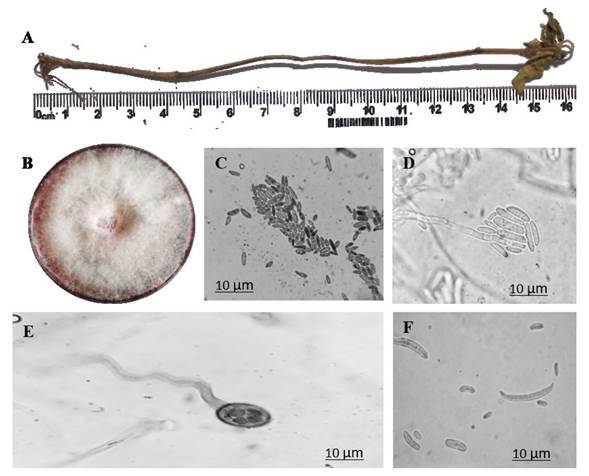
Figure 3 Reproduction structures of F. solani (MX-MIC 798) after 10 days in a PDA medium at 28 °C. A) dead Capsicum annuum var Miahuateco plant inoculated with F. solani MX-MIC 798, B) F. solani culture in PDA medium; C) microconidia of F. solani; D) phialides and microconidia in false F. solani heads; E) F. solani chlamydospores; F) F. solani micro and macroconidia.
Conclusions
The strains T. harzianum (TH-4) and T. koningiopsis (T-K11) displayed classes II and III of antagonism, respectively, on F. solani on the Bell scale. In additionT. harzianum TH-4 displayed the highest rate of development, speed of growth and percentage of inhibition of PICR radial growth on F. solani MX-MIC 798 (53%) under in vitro conditions.
In nursery conditions, T. harzianum TH-4 (82%) and T. koningiopsis T-K11 (74%) presented the best response in the germination of Miahuateco variety chili plants, with the highest percentage of germination in comparison with the chemical treatment (72%), although T. harzianum TH-4 reduced rot caused by the F. solani strain by 75%.
In the town of Santa María Tecomavaca, in the state of Oaxaca, Mexico, T. harzianum (T-H4) and the Mancozeb 80® chemical treatment presented the best response in the reduction of the mortality and severity of the disease in Miahuateco chili plants 40 dat.
Literatura citada
Agrios GN. 2006. Plant Pathology. 5 Edition. Elsevier the Academic Press. London. 838 p. [ Links ]
Al-Fadhal FA, Al-Abedy AN and Alkhafije DA. 2019. Isolation and molecular identification of Rhizoctonia solani and Fusarium solani isolated from cucumber (Cucumis sativus L.) and their control feasibility by Pseudomonas fluorescens and Bacillus subtilis. Egyptian Journal of Biological Pest control 29(47): e2752. https://doi.org/10.1186/s41938-019-0145-5 [ Links ]
Andrade-Hoyos P, Luna-Cruz A, Hernández EO, Gayosso EM, Valenzuela NL and Cureño HJB. 2019. Antagonismo de Trichoderma spp. vs. hongos asociados a la marchitez de chile. Revista Mexicana de Ciencias Agrícolas 10(6): 1259-1272. https://doi.org/10.29312/remexca.v10i6.1326 [ Links ]
Andrade-Hoyos P, Silva-Rojas HV and Romero-Arenas O. 2020. Endophytic Trichoderma Species Isolated from Persea americana and Cinnamomum verum Roots Reduce Symptoms Caused by Phytophthora cinnamomi in avocado. Plants e1220. https://doi.org/10.3390/plants9091220 [ Links ]
Barnett HL and Hunter BB. 1998. Illustrated genera of imperfect fungi. 4th Edition, The American Phytopatological Society, St. Paul Minnesota. USA. 241 p. [ Links ]
Bell DK, Wells HD and Markham CR. 1982. In vitro antagonism of Trichoderma species against six fungal plant pathogens. Phytopathology 72: 379-382. https://doi.org/10.1094/Phyto-72-379 [ Links ]
Black LL, Green KS, Hartman LG and Poulos MJ. 1993. Pepper Diseases: A Field Guide. Asian Vegetable Research and Development Center. Shanhua, Taiwan. 98 p. [ Links ]
Bokhari NA and Perveen K. 2013. Antagonistic action of Trichoderma harzianum and Trichoderma viride against Fusarium solani causing root rot of tomato. African Journal of Microbiology Research 6(44): 7193-7197. https://doi.org/10.5897/AJMR12.956 [ Links ]
Castillo-Aguilar C, Coh-Méndez D, Carrillo-Ávila E and Monsalvo-Espinosa A. 2015. Producción de planta de chile habanero (Capsicum chinense Jacq). Agroproductividad 8(4): 73-78. https://revista-agroproductividad.org/index.php/agroproductividad/article/view/676 [ Links ]
Chen ZH, Gao T, Liang SP, Liu KX, Zhou MG and Chen CJ. 2014. Molecular mechanism of resistance of Fusarium fujikuroi to benzimidazole fungicides. Fems microbiology letters 357(1): 77-84. https://doi.org/10.1111/1574-6968.12504 [ Links ]
Cole JS and Zvenyika Z. 1988. Integrated control of Rhizoctonia solani and Fusarium solani in tobacco transplants with Trichoderma harzianum and triadimenol. Plant Pathology 37(2): 271-277. https://doi.org/10.1111/j.1365-3059.1988.tb02073.x [ Links ]
Duarte-Leal Y, Lamz-Piedra A and Martínez-Coca B. 2017. Antagonismo in vitro de aislamientos de Trichoderma asperellum Samuels frente a Sclerotium rolfsii Sacc. Revista de Protección Vegetal 32(3): 1-11. http://scielo.sld.cu/pdf/rpv/v32n3/rpv03317.pdf [ Links ]
Dubey SC, Suresh M and Singh B. 2007. Evaluation of Trichoderma species against Fusarium oxysporum f. sp. ciceris for integrated management of chickpea wilt. Biological control 40(1): 118-127. https://doi.org/10.1016/j.biocontrol.2006.06.006 [ Links ]
Ezziyyani M, Sánchez CP, Ahmed AS, Requena ME and Castillo MEC. 2004. Trichoderma harzianum como biofungicida para el biocontrol de Phytophthora capsici en plantas de pimiento (Capsicum annuum L.). Anales de biología 26: 35-45. https://revistas.um.es/analesbio/article/view/30441 [ Links ]
Gan H and Wickings K. 2017. Soil ecological responses to pest management in golf turf vary with management intensity, pesticide identity, and application program. Agriculture, Ecosystems and Environment 246: 66-77. https://doi.org/10.1016/j.agee.2017.05.014 [ Links ]
Guenoun K, Chattaoui M, Bouri M, Rhouma A, Naghmouchi K and Raies A. 2019. Biological control of growth promoting rhizobacteria against verticillium wilt of pepper plant. Biologia 74: 237-250. https://doi.org/10.2478/s11756-018-00169-9 [ Links ]
Guigón-López C, Guerrero-Prieto V, Vargas-Albores F, Carvajal-Millán E, Ávila-Quezada GD, Bravo-Luna L, Ruocco M, Lanzuise S, Woo S and Lorito M. 2010. Identificación molecular de cepas nativas de Trichoderma spp., su tasa de crecimiento in vitro y antagonismo contra hongos fitopatógenos. Revista Mexicana de Fitopatología 28(2): 87-96. http://www.scielo.org.mx/pdf/rmfi/v28n2/v28n2a2.pdf [ Links ]
Hernández-Melchor DJ, Ferrera-Cerrato R and Alarcón A. 2019. Trichoderma: importancia agrícola, biotecnológica y sistemas de fermentación para producir biomasa y enzimas de interés industrial. Chilean Journal of Agricultural and Animal Sciences 35(1): 98-112. https://dx.doi.org/10.4067/S0719-38902019005000205 [ Links ]
Hyder S, Inam UH, Bibi S, Humayun A, Ghuffar S and Iqbal S. 2017. Novel potential of Trichoderma spp., as biocontrol agent. Journal of Entomology and Zoology Studies 5(4): 214-222. https://www.entomoljournal.com/archives/2017/vol5issue4/PartC/5-3-244-140.pdf [ Links ]
Illa C, Andrés-Pérez A, Matias T and Pérez MA. 2019. Efecto de biocontrol y promoción del crecimiento en maní por Trichoderma harzianum y Bacillus subtilis en condiciones controladas y campo. Revista Mexicana de Fitopatología 38(1): 119-131. https://doi.org/10.18781/R.MEX.FIT.1910-6 [ Links ]
Instituto Nacional de Estadística, Geografía e Informática (INEGI). 2017. Áreas geográficas. Coordenadas Oaxaca. (Consultado: 01 de julio de 2020). https://www.inegi.org.mx/app/areasgeograficas/?ag=20 [ Links ]
Jagraj C, Vipul K, Seweta S, Adesh K and Vinit PS. 2018. In vitro evaluation of Trichoderma species against Fusarium oxysporum f. sp. lycopersici causing tomato wilt. Plant Pathology Journal 17: 59-64. https://doi.org/10.3923/ppj.2018.59.64 [ Links ]
Launio CC, Labon KO, Bañez AM and Batani RS. 2020. Adoption and economic analysis of using biological control in Philippine highland farms: Case of Trichoderma koningii strain KA. Crop Protection e105177. https://doi.org/10.1016/j.cropro.2020.105177 [ Links ]
Leroux P and Walker AS. 2011. Multiple mechanisms account for resistance to sterol 14α‐demethylation inhibitors in field isolates of Mycosphaerella graminicola. Pest Management Science 67(1): 44-59. https://doi.org/10.1002/ps.2028 [ Links ]
Madhavi M, Kumar CPC, Reddy DRR and Singht TK. 2006. Integrated management of wilt of chilli incited by Fusarium solani. Indian Journal of Plant Protection 34(2): 225-228. [ Links ]
Mannai S, Jabnoun-Khiareddine H, Nasraoui B and Daami-Remadi M. 2018. Rhizoctonia root rot of pepper (Capsicum annuum): Comparative pathogenicity of causal agent and biocontrol attempt using fungal and bacterial agents. Journal of Plant Pathology and Microbiology 9(2): 431-439. https://doi.org/10.4172/2157-7471.1000431 [ Links ]
Michel-Aceves AC, Hernández-Morales J, Toledo-Aguilar R, López JES and Romero-Rosales T. 2019. Capacidad antagónica de Trichoderma spp., nativa contra Phytophthora parasitica y Fusarium oxysporum aislados de cultivos de Jamaica. Revista Fitotecnia Mexicana 42(3): 235-241. https://doi.org/10.35196/rfm.2019.3.235-241 [ Links ]
Michel-Aceves AC, Otero-Sánchez MA, Rebolledo-Domínguez O, Lezama-Gutiérrez R, Ariza-Flores R and Barrios-Ayala A. 2005. Producción y efecto antagónico de quitinasas y glucanasas porTrichodermaspp., en la inhibición deFusarium subglutinansyFusarium oxysporum in vitro .Revista Chapingo Serie Horticultura 11:273-278. https://www.redalyc.org/pdf/609/60911213.pdf [ Links ]
Montealegre J, Donoso S, Herrera R and Besoain X. 2003. Identificación de Fusarium solani (Mart.) Sacc. como agente causal de la podredumbre del pie de tomate. Boletín Micológico 18: 53-55. https://revistas.uv.cl/index.php/Bolmicol/article/view/426/387 [ Links ]
Morales ML, Andrade-Hoyos P, Valencia-de Ita MA, Romero-Arenas O, Silva-Rojas HV and Contreras-Paredes CA. 2020. Caracterización de hongos asociados al cultivo de fresa y efecto antagonista in vitro de Trichoderma harzianum. Revista Mexicana de Fitopatología 38(3): 434-449. http://dx.doi.org/10.18781/R.MEX.FIT.2005-7 [ Links ]
Nagendran K, Loganathan M, Saha S, Mishra A, Pandey KK, Rai RM, Gupta S, Rai AB and Singh B. 2016. Biological management of wilt disease on chilli caused by Fusarium solani .Vegetable Science 43(2): 253-256. https://www.indianjournals.com/ijor.aspx?target=ijor:vgt&volume=43&issue=2&article=019 [ Links ]
Nawrocka J, Małolepsza U, Szymczak K and Szczech M. 2018. Involvement of metabolic components, volatile compounds, proteins, and mechanical strengthening in multilayer protection of cucumber plants against Rhizoctonia solani activated by Trichoderma atroviride TRS25. Protoplasma 255(1): 359-373. https://doi.org/10.1007/s00709-017-11571 [ Links ]
Pérez-Carrasco LJ, Tornero-Campante M A, Escobedo-Garrido JS and Sandoval-Castro E. 2017. El chile poblano criollo en la cultura alimentaria del Alto Atoyac. Estudios Sociales 27(49): 47-66. http://www.scielo.org.mx/pdf/estsoc/v27n49/0188-4557-estsoc-27-49-00047.pdf [ Links ]
Pérez-Torres E, Bernal-Cabrera A, Milanés-Virelles P, Sierra-Reyes Y, Leiva-Mora M, Marín-Guerra S and Monteagudo-Hernández O. 2018. Eficiencia de Trichoderma harzianum (cepa a-34) y sus filtrados en el control de tres enfermedades fúngicas foliares en arroz. Bioagro 30(1): 17-26. https://doi.org/10.1080/21501203.2017.142312 [ Links ]
Perveen K, Haseeb A and Shukla PK. 2010. Effect of Sclerotinia sclerotiorum on the disease development, growth, oil yield and biochemical changes in plants of Mentha arvensis. Saudi Journal of Biological Sciences 17(4): 291-294. https://doi.org/10.1016/j.sjbs.2010.05.008 [ Links ]
Rentería-Martínez ME, Guerra-Camacho MA, Ochoa-Meza A, Moreno-Salazar F, Meza-Moller CA and Guzmán Ortíz JM. 2018. Descripción y comparación entre morfotipos de Fusarium brachygibbosum, F. falciforme y F. oxysporum patogénicos en sandía cultivada en Sonora, México. Revista Mexicana de Fitopatología 37(1): 16-34. https://doi.org/10.18781/R.MEX.FIT.1808-1 [ Links ]
Reyes-Tena A, Rodríguez-Alvarado G, Santillán-Mendoza R, Díaz-Celaya M and Fernández-Pavía SP. 2019. Marchitez causada por Fusarium solani en chile chilaca (Capsicum annuum) en Michoacán. Revista Mexicana de Fitopatología 37(1): 43-47. http://dx.doi.org/10.18781/R.MEX.FIT.1904-1 [ Links ]
Rivera-Jiménez MN, Zavaleta-Mancera HA, Rebollar-Alviter A, Aguilar-Rincón VH, Garcia- Santos G, Vaquera-Huerta H and Silva-Rojas HV. 2018. Phylogenetics and histology provide insight into damping-off infections of ‘Poblano’ pepper seedlings caused byFusariumwilt in greenhouses. Mycological Progress 17: 1237-1249. https://doi.org/10.1007/s11557-018-1441-2 [ Links ]
Rodríguez J, Olvera BVP, Muñoz AG, Corona BM, Manzo F and Liendo LS. 2007. Rescate in situ del chile Poblano en Puebla, México. Revista Fitotecnia Mexicana 30(1): 25-32.http://www.redalyc.org/articulo.oa?id=61030103 [ Links ]
Romero-Arenas O, Amaro LJ, Damián HM, Valencia de Ita MA, Rivera A and Huerta LM. 2017. Bio-preparados de Trichoderma spp., para el control biológico de Phytophthora capsici en el cultivo de tomate de Puebla, México. ITEA 113(4): 313-324. http://doi.org/10.12706/itea.2017.019 [ Links ]
Samson R, Visagie CM, Houbraken J, Hong SB, Hubka V, Klaassen CH, Perrone G, Seifert KA, Susca A, Tanney JB, Varga J, Kocsubé S, Szigeti G, Yaguchi T and Frisvad JC. 2014. Phylogeny, identification and nomenclature of the genus Aspergillus. Mycology 78: 141-173. https://doi.org/10.1016/j.simyco.2014.07.004 [ Links ]
Sánchez-García BM, Espinosa-Huerta E, Villordo-Pineda E, Rodríguez-Guerra R and Mora-Avilés MA. 2017. Identificación molecular y evaluación antagónica in vitro de cepas nativas de Trichoderma spp., sobre hongos fitopatógenos de raíz en frijol (Phaseolus vulgaris L.) cv. Montcalm. Agrociencia 51(1): 63-79. http://www.scielo.org.mx/pdf/agro/v51n1/1405-3195-agro-51-01-00063-en.pdf [ Links ]
Servicio de Información Agroalimentaria y Pesquera (SIAP) 2020. Atlas Agroalimentario. 1080-2020. https://www.gob.mx/siap [ Links ]
Suárez C, Fernández R, Valero N, Gámez R and Páez A. 2008. Antagonismo in vitro de Trichoderma harzianum Rifai sobre Fusarium solani (Mart.) Sacc., asociado a la marchitez en maracuyá. Revista Colombiana de Biotecnología 10(2): 35-43. https://doi.org/10.15446/rev.colomb.biote [ Links ]
Sundaramoorthy S and Balabaskar P. 2013. Biocontrol efficacy of Trichoderma spp., against wilt of tomato caused by Fusarium oxysporum f. sp. lycopersici. Journal of Applied Biology and Biotechnology 1(3): 36-40. https://www.jabonline.in/abstract.php?article_id=15 [ Links ]
Thangavelu R, Palaniswami A and Velazhahan R. 2004. Mass production of Trichoderma harzianum for managing Fusarium wilt of banana. Agriculture, Ecosystems and Environment 103(1): 259-263. https://doi.org/10.1016/j.agee.2003.09.026 [ Links ]
Toledo-Aguilar R, López-Sánchez H, López PA, Guerrero-Rodríguez JDD, Santacruz-Varela A, and Huerta-de la Peña A. 2016. Diversidad morfológica de poblaciones nativas de chile Poblano. Revista Mexicana de Ciencias Agrícolas 7(5): 1005-1015. https://doi.org/10.29312/remexca.v7i5.227 [ Links ]
Tsegaye-Redda ET, Ma J, Mei J, Li M, Wu B and Jiang X. 2018. Biological control of soilborne pathogens (Fusarium oxysporumf. sp.cucumerinum) of cucumber (Cucumis sativus) byTrichodermasp. Journal of Life Sciences 12:1-12. https://doi.org/10.17265/1934-7391/2018.01.001 [ Links ]
Vázquez-Casarrubias G, Escalante-Estrada JAS, Rodríguez-González M, Ramírez-Ayala C and Escalante-Estrada LE. 2011. Edad al trasplante y su efecto en el crecimiento y rendimiento de chile Apaxtleco. Revista Chapingo Serie Horticultura 17(1): 61-65. http://www.scielo.org.mx/pdf/rcsh/v17n1/v17n1a10.pdf [ Links ]
Yedidia I, Benhamou N and Chet I. 1999. Induction of defense responses in cucumber plants (Cucumis sativus L.) by the biocontrol agent Trichoderma harzianum. Applied and Environmental Microbiology 65(3): 1061-1070. https://doi.org/10.1128/AEM.65.3.1061-1070.1999 [ Links ]
Zeravakis G, Philippoussis A, Ioannidou S and Diamantopoulou P. 2001. Mycelium growth kinetics and optimal temperature conditions for the cultivation of edible mushroom species on lignocellulosic substrates. Folia Microbiologica 46(3): 231-234. https://doi.org/10.1007/BF02818539 [ Links ]
Ziogas BN and Malandrakis AA. 2015. Sterol Biosynthesis Inhibitors: C14 Demethylation (DMIs). In: Ishii H., Hollomon D. (eds) Fungicide Resistance in Plant Pathogens. Springer, Tokyo. 199-216 pp. https://doi.org/10.1007/978-4-431-55642-8_13 [ Links ]
Received: January 25, 2021; Accepted: March 18, 2021
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
